
Abstract
In order to facilitate/expedite the production of effective and affordable snake antivenoms, a novel in vitro potency assay was previously developed. The assay is based on an antiserum’s ability to bind to postsynaptic neurotoxin (PSNT) and thereby inhibit the PSNT binding to the nicotinic acetylcholine receptor (nAChR). The assay was shown to work well with antiserum against Thai Naja kaouthia which produces predominantly the lethal PSNTs. In this work, the assay is demonstrated to work well with antiserum/antivenom against Bungarus candidus (BC), which also produces lethal presynaptic neurotoxins, as well as antivenom against Sri Lankan Naja naja (NN), which produces an abundance of cytotoxins. The in vitro and in vivo median effective ratios (ER50s) for various batches of antisera against BC showed a correlation (R2) of 0.8922 (p < 0.001) while the corresponding value for the anti-NN antivenom was R2 = 0.7898 (p < 0.01). These results, together with the known toxin profiles of various genera of elapids, suggest that this in vitro assay could be used with antisera against other species of Bungarus and Naja and possibly other neurotoxic snake venoms worldwide. The assay should significantly save numerous lives of mice and accelerate production of life-saving antivenoms.
Similar content being viewed by others

Introduction
Snake envenomation is an important yet neglected medical problem in various developing countries with an estimated annual envenomation world wide of about 5.5 million cases1,2. Effective and affordable antivenoms (AV) which are the mainstay for treatment remain unavailable in several parts of the world while research efforts are undertaken to solve this problem3. In the development and production of an AV, an important step involves the in vivo assay to evaluate the potency of the produced antiserum/antivenom. The standard murine lethality neutralizing assay is considered by WHO as an essential AV potency assay. This assay is used to find first, the median lethal dose (LD50) that determines the lethality of the venom, and the median effective dose (ED50) of the AV4,5. The assay is expensive, laborious and, due to biological variation, often give highly variable results. In addition, some murine lethality results might not be consistent with the relevant efficacy outcomes in humans6,7. In Thailand, as well as in many Buddhist countries, it is very difficult to find researchers or students who would agree to do these in vivo experiments.
Because of these reasons, various in vitro assays have been developed to reduce or replace the murine lethality assay. The most widely used assay is based mainly on ELISA8,9,10 but some of these assays have often been shown to give poor correlation with the in vivo assay11,12. Moreover, the antigen-antibody ‘binding’ reaction of ELISA may not result in the ‘neutralization’ of the antigen, and therefore alternative in vitro assay should be developed.
Recently, a novel in vitro assay using solubilized, purified nicotinic acetylcholine receptor (nAChR) binding has been developed for AV potency assay against the Thai cobra Naja kaouthia13. This elapid snake produces mainly postsynaptic neurotoxins (PSNT) which binds specifically and with high affinity to the nAChR at the muscle endplate and thereby inhibits the neuro-muscular transmission causing death in animals. The in vitro assay therefore closely mimics the toxicological reactions in vivo; and the in vivo and in vitro potency assays showed a correlation R2 = 0.9807, p < 0.0001. The assay should also work well with AVs against elapids in the genera other than Naja which produce mainly or exclusively post-synaptic toxins (PSNTs) e.g., King cobra, Ophiophagus hannah14.
However, snakes in the genus Bungarus (kraits) also produce, in addition to PSNTs, the lethal presynaptic neurotoxins (β-neurotoxins) which in elapids, belong to the group 1 phospholipase A2 enzymes. These toxins do not bind to nAChR but react with receptors on the membrane of the motor nerve terminals which contain the acetylcholine vesicles. The toxins hydrolyze the phospholipids of the plasma membrane, resulting in the loss of synaptic vesicles in the nerve terminal. Eventually the nerve terminals degenerate with the failure of the neuromuscular transmission15. The LD50 of the β-neurotoxins is about 10 ng/g16 which is considerable lower than that of the PSNT (0.18 μg/g mouse)17. It is therefore conceivable that in the case of some Bungarus venoms, death could be caused, at least in part, by the β-neurotoxins; but this effect would not be measured by the nAChR binding of the in vitro potency assay. Consequently the developed in vitro assay might not be useful for assay of AV against Bungarus spp.
Another interesting case is the Naja naja (Indian cobra) which is a WHO category 1 medically most important elapid in India, Pakistan and Sri Lanka. Envenomation by this snake resulted in muscle weakness and death by respiratory failure which is likely the effect of the venom PSNTs. Interestingly, the venom of the Sri Lankan snake was shown by proteomics study to contain 71.55% of cytotoxins (cardiotoxins)18. These three finger toxins (3FTs) cause severe local tissue necrosis in most (88%) of the victims18 and could possibly contribute to the venom lethality. It is therefore interesting to investigate whether the developed in vitro nAChR binding assay could be used for potency assay of AV against this cobra.
We report here that the in vitro nAChR binding assay, when used in the potency determinations of AVs against Bungarus candidus (Thailand) and Naja naja (Sri Lanka), gave high correlation with the corresponding in vivo murine lethality neutralization assays.
Results
Studies on the optimal conditions of the in vitro AV potency assay
The optimal concentrations of NK3, nAChR, rat anti-nAChR antibody and goat-anti-rat HRP conjugate used in the assay
The optimal concentrations of these four reagents used in the in vitro potency assay were described in a previous study13. The optimal concentration of NK3 for coating the plates was 15 µg/ml, and 0.707 µg/ml of nAChR for binding to the NK3 coated plate. Rat anti-nAChR serum and goat anti-rat-IgG conjugated HRP were used at 1:1600 dilution and 1:4500 dilution, respectively.
Inhibition of nAChR binding to NK3-coated plate by B. candidus or N. naja venom
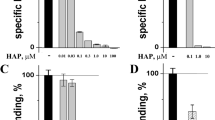
Crude B. candidus and N. naja venoms were separately used to determine the 50% inhibition of the nAChR binding (in vitro IC50). In the first step, various concentrations of crude B. candidus (or N. naja) venom and the purified nAChR (0.707 µg/ml) were incubated for 1 h at 25 °C. After the incubation, the solution was added to the NK3 coated plates. The concentration of the venom that blocked the binding of nAChR by 50% was the IC50. The results (Fig. 1) showed that the IC50s of B. candidus and N. naja venoms were 0.1625 ± 0.0172 µg/ml and 0.4067 ± 0.0292 µg/ml, respectively.

Inhibition of nAChR binding by B. candidus and N. naja venoms to NK3-coated plate. Data were means ± SD of 3 determinations.
Neutralization of B. candidus or N. naja venom by horse monospecific antisera against B. candidus or by Vins antivenom against N. naja as determined by nAChR binding to the NK3-coated plate
Nine horse monovalent anti-B. candidus sera were 2-fold diluted from 1:500 to 1:512,000. These diluted sera of different horses were separately incubated with 5xIC50 of B. candidus venom (1.4029 µg/ml) in the ‘Pre-incubation 1′ experiment. The reaction mixtures were ultrafiltered and subjected to ‘Pre-incubation 2′. The resulting reaction mixtures were then added to the NK3-coated plates. The nAChR binding to the plate was read at OD450nm and the results which were expressed as % nAChR binding are shown in Fig. 2A. The corresponding results for the N. naja venom and the Vins anti-N. naja antivenom are shown in Fig. 2B.

Neutralization of B. candidus venom by horse anti-B. candidus sera (A) and neutralization of N. naja venom by Vins anti-N. naja antivenom (B) as determined by nAChR binding to NK3-coated plate. The results were means ± SD of 3 determinations.
The in vitro ER50s, expressed as µg of B. candidus venom neutralized per µl of antiserum, and the in vivo ER50s (mg venom/ml antiserum) of the 9 horse anti-B. candidus sera are shown in Table 1. The correlation between the in vitro ER50s and the in vivo ER50s of the anti-B. candidus sera as shown in Fig. 3A was R2 = 0.8922 (p < 0.001).

The correlation between the in vitro and in vivo ER50s of the anti-B. candidus sera (A) and of the anti-N. naja antivenom (B).
The in vitro and in vivo ER50s of the horse anti-N. naja antivenom are shown in Table 2. The correlation between the in vitro ER50s and the in vivo ER50s of the anti-N. naja antivenom as shown in Fig. 3B was R2 = 0.7898 (p < 0.01).
Discussion
The in vivo potency assay is an important step in antivenom development and production. Because of the complexity of the snake venoms, which contain various diverse pharmacologically active proteins, different pre-clinical assays are needed to test antivenom efficacy against the various components. The gold standard of antivenom efficacy tests is the murine lethality neutralization assay, which is the only ‘Essential test’ included in the WHO Guidelines on Antivenoms for Production, Control and Regulation of Snake Antivenom Immunoglobulins4. Depending on the pharmacological profile of a given venom, several other supplementary ‘Recommended tests’ have been proposed e.g., neutralization assays of hemorrhagic, nephrotoxic, myotoxic, neurotoxic, edema-forming, dermonecrotic and measurement of in vitro coagulant and defibrinogenating activities4.
However, these assays, especially the ‘Essential’ murine lethality neutralization assay, which involves the killing a large number of animals, are faced with various experimental, ethical, religious and regulatory difficulties. Consequently, many in vitro assays have been developed, mostly based on ELISA formats, to be used in place of, or to reduce the use of, in vivo murine assays. However, because ELISA involves the binding of antivenom antibodies to venom proteins whose toxicity might not be neutralized, results of these in vitro assays usually show poor correlation with results from the murine lethality neutralization assay. We have recently developed a novel in vitro assay13 based on nAChR binding to determine the potency of antivenom against Naja kaouthia, an elapid that produces, almost exclusively, postsynaptic neurotoxins that are responsible for death in victims. This assay estimates the binding of venom postsynaptic neurotoxins to nAChR, which is the toxicological reaction involved in the lethal effects of cobra envenomation. In this study, it is shown that the in vitro assay should be suitable for the evaluation of horse antisera against Bungarus candidus, which produces clinically relevant lethal presynaptic neurotoxins (β-neurotoxins) in addition to PSNT. Moreover, the assay could also be used to evaluate commercial F(ab’)2 antivenoms against Naja naja (Sri Lanka), which produces very high levels of cytotoxins in its venom18.
Bungarus candidus, also known as the Malayan krait, is a WHO Category 1 medically important snake in Southeast Asian countries. The venom of this snake contains β-neurotoxins that are highly lethal, in addition to PSNTs19,20. These toxins are also produced by several other krait species. They have molecular weights (MW) ranging from 18–22 kDa and are heterodimers with one subunit exhibiting phospholipase A2 activity21. They do not bind to nAChR; however, they cause neuromuscular blockade leading to death by respiratory failure16. The venom of B. candidus, (Thailand), contains about 26.9% long and short 3FT of subtype PSNT and about 13.5% of β-neurotoxins (CH Tan, unpublished data).
In the present study, it was shown that the in vitro nAChR binding potency activity of antisera against B. candidus venom was highly correlated with the murine in vivo lethality neutralization assay results (R2 = 0.8922, p < 0.001). Since the in vitro assay measures postsynaptic neurotoxin binding, and therefore does not detect the clinically important binding of β-neurotoxin to nAChR, it is of interest to know the reason behind the observed high correlation between the results of this assay and those of the in vivo lethality neutralization assay when evaluating B. candidus antisera. This observation may be due to the experimental conditions used for the in vivo lethality neutralization assay described in WHO Guidelines4. When the mice were challenged with 5xLD50 of the venom, the smaller (7–8 kD) faster acting postsynaptic neurotoxin rapidly inhibits the nAChR at the muscle endplate resulting in neuromuscular blockage and death in mice. The β-neurotoxin, which is larger (18–22 kD) and slower acting, has to undergo a number of steps before finally depleting the acetylcholine store of the synaptic vesicles to completely inhibit the neuromuscular transmission. Thus, in the lethality assay (LD50 determination), death was likely due to the action of postsynaptic neurotoxins before the action of β-neurotoxins took effect. In the in vivo ED50 determination, when the antiserum antibodies were added to react with the venom, the antibodies most likely neutralized both the postsynaptic and β-neurotoxins. This is because the β-neurotoxins are larger and has a more complex chemical structure than the postsynaptic neurotoxins. Consequently, the β-neurotoxin should be more immunogenic and have more antibodies against it in the antisera. Thus, under the in vivo lethality neutralization assay, postsynaptic neurotoxin plays the major role in determining the lethality neutralization activity. Therefore, the high correlation between the two assays could be due to the fact that both the in vivo and in vitro assays primarily measured the activity of the postsynaptic neurotoxins. If this is in fact the case, then the murine lethality neutralization assay, when applied to B. candidus antivenom, is another example of an antivenom potency test where the clinically relevant toxin in human envenomation is not measured6,7.
Thus, in the development and production of antivenom against B candidus, it may be necessary to perform an additional ‘neurotoxicity neutralization assay’ to determine the neutralization activity for the β-neurotoxin in the venom. This can be done according to the WHO ‘Recommended antivenom efficacy test’ guidelines, which involve the use of isolated chick biventer cervicis nerve-muscle preparations and/or mouse hemidiaphragm phrenic nerve preparations4. This ex vivo assay should also shed light on the inhibition of β-neurotoxin in the present in vivo assay. However, the tests are difficult to perform, require costly equipment and expert technological help and are unlikely to be practicable for most laboratories. Because of the technical difficulty involved in the neurotoxicity assay, WHO Guidelines4 concluded that the murine lethality neutralization assay is a reliable assay in predicting the neutralization of neurotoxicity of venoms.
Naja naja, i.e. the Indian cobra, is also a WHO Category 1 medically important snake in South Asia. The percent abundances of α-neurotoxins were only 4.8% and 8.91% in N. naja (India) and N. naja (Sri Lanka) venoms, respectively, while the cytotoxins were the most abundant venom 3FT subtype in both N. naja (India) (69.34%) and N. naja (Sri Lanka) (71.55%)18. The cytotoxins, also known as cardiotoxins, are polypeptides with a MW of about 6–7 KDa, and are found only in cobra venom22. At lower concentrations, cardiotoxins increase the heart rate and, at higher concentrations, kill the animal by inducing cardiac arrest23. Thus the abundant cytotoxins in N. naja venom might contribute to lethality in the in vivo assay. Since the in vitro nAChR assay estimates only the nAChR binding of the α-neurotoxins, it is possible that this difference could affect the correlation between the in vivo and in vitro potency assays and limit the usefulness of the assay for N. naja antivenom. However, it was shown in the present study that for the antivenom against N.naja (Sri Lanka), a good correlation between the in vitro nAChR-PSNT binding activity and the in vivo murine lethality neutralization activity was obtained. The reason behind this observation could be that the presence of a large amount of cytotoxins in the venom did not affect the results of the in vitro nAChR - PSNT binding assay since the assay has been shown to be specific to PSNT and not to cytotoxins13. Furthermore, the results suggested that death is mainly the result of the α-neurotoxins since α-neurotoxins are about 10 times more toxic, with an LD50 of ca. 0.1 μg/g in mice while that of the cytotoxins is about 1.0–1.5 μg/g24,25.
The observations that the in vitro potency assay based on nAChR-PSNT binding could be used to evaluate antivenoms against B. candidus and N. naja may have important implications for the use of this assay to assess antivenoms against other elapids. For example, Micrurus nigrocinctus, the Central American coral snake, is a medically important snake which has been shown in proteomics studies to contain a variety of toxins. These included several isoforms of short-chain α-neurotoxins as well as muscarinic-like toxins and proteins with similarity to long-chain κ-2 bungarotoxin while the α-neurotoxins predominated26,27. Alape-Giron et al.28 have shown that various venom proteins of M. nigrocinctus specifically bind to the Torpedo spp. acetylcholine receptor while the majority of PLA2 fractions exhibit low lethality. The predominant effects of the M. nigrocinctus venom are neurotoxicity and myotoxicity with death from the venom to be due to respiratory failure caused by the α-neurotoxins. The α-neurotoxins of this venom play a leading role in the lethality of this venom27,28. Thus, it is highly likely that the present in vitro assay could give high correlation with the in vivo murine lethality neutralization assay, and could be used for antivenom potency assays for this snake.
Another interesting group of elapids are the Dendroaspis spp. For example, Dendroaspis polylepis, an African elapid known as the black mamma, is an extremely deadly snake. The venom of this snake contains postsynaptic neurotoxins29 and also dendrotoxins, which are unique to the genus Dendroaspis30. Dendrotoxins are Kunitz-type proteinase inhibitors. They block the voltage-dependent potassium channels resulting in the facilitation of the release of neurotransmitters at the presynaptic nerve terminal31. The α-neurotoxins in Dendroaspis polylepis venom are the most toxic proteins in the venom, followed by dendrotoxins. Death due to black mamba envenomation is caused mainly by the postsynaptic neurotoxins, while the less toxic dendrotoxins probably play a secondary role in the lethal action of the venom31,32. Based on this information, it is quite likely that the present in vitro potency assay will be applicable to the antisera against D. polylepis as well. A similar conclusion could probably be made with Hydrophis schistosus (sea snake) and Laticauda colubrina (sea krait) that produce mainly short-chain and long-chain α-neurotoxins, respectively, which have been shown to cause paralysis and death in animals33,34. If these predictions are shown to be the case experimentally, the assay is likely to be useful for potency assays of antivenoms against most elapids of the world.
The Vins antivenom against N. naja used in this study was a F(ab′)2 antivenom. ‘Pre-incubation 1′ of the N. naja venom with the F(ab′)2 antibody was followed by ultrafiltration using a membrane with a MWCO of 50kD, instead of the customary 100 kD membrane13. This was done to remove the bound and free F(ab′)2 antibody, which has a molecular weight of about 100 kDa.
The ability to use the F(ab′)2 as well as IgG antibody in the in vitro assay is more advantageous compared to ELISA. In ELISA where a secondary antibody is used, different signal intensity will be obtained when the first antibody is a F(ab′)2 or an IgG. Of the two, use of an IgG will result in more secondary antibody binding (to the Fc portion) resulting in the generation of a relatively stronger color signal. In ELISA potency assays, where some samples consisted of IgG and some with F(ab′)2 antivenoms, comparison between the results may not be possible due to the above consideration. However, this difference in the antibody chemical structures should not be a problem in the present nAChR-PSNT binding assay.
An interesting aspect of this study was that the Vins antivenom is polyspecific against 2 vipers and 2 elapids. Therefore, the antivenom also contained F(ab′)2 antibody against the viper proteins but the ‘heterologous’ antibody did not seem to interfere with the nAChR assay as shown by the good correlation result observed. This in vitro assay, unlike most ELISA, which employ crude venoms as the antigens and thus often show cross reactivity of the heterologous antibodies with other venom proteins, gave more precise results as would be expected from the highly specific binding of α-neurotoxin to nAChR.
The present in vitro potency assay should be very useful in the development and production of effective antivenoms against elapid snakes which, in various countries, are the most medically important cause of snakebite morbidity and mortality. In the development of new antivenoms, whether traditional serum based or synthetic35, potency assays of the products have to be carried out. The lethality neutralization assay is an essential pre-clinical test for an antivenom. The present in vitro assay could be used to reduce, or in some cases replace, the in vivo assay during AV development. Even with the final antivenom product, which must be assayed using the murine lethality assay36, the in vitro assay could be employed in dose range determination and consequently could reduce the use of mice in this trial and error process.
The nAChR-PSNT binding assay is simple, less variable, less time-consuming and less expensive and could facilitate the development of effective and wide-paraspecific ‘universal’ antivenoms. This would ultimately reduce the mortality and morbidity of snake envenomation, which occurs mostly in poor countries/regions of the world37.
Materials and Methods
The venom of Bungarus candidus which was from a pool of several adult snakes of Thai origin, and the horse monovalent antisera against B. candidus were purchased from Queen Soavabha Memorial Institute (QSMI). Lyophilized Naja naja (Sri Lanka) milked from a large pool of adult snakes was obtained from Dr C. Ariaranee. The polyspecific equine lyophilized F(ab′)2 antivenom was from Vins Bioproduct Limited, Hyderabad, India; it was prepared against saw-scaled viper (Echis carinatus), Indian cobra (N. naja), Russell’s viper (Daboia russelii) and common krait (Bungarus caeruleus), Batch no. 01AS12041, Exp date 03/2018. Naja kaouthia postsynaptic toxin 3 (NK3) was purified according to Karlsson et al.38. All other reagents were from Sigma Chemical, St Louis. Missouri, unless stated otherwise. nAChR from Torpedo californica electroplaque was purified and the anti-nAChR antisera was generated in rats as described by Ratanabanangkoon et al.13.
In vivo neutralizing activities of horse monospecific antisera against B. candidus venom and F(ab′)2 antivenom against N. naja venom
The median lethal doses (LD50s) of B. candidus and N. naja venoms, were determined by intravenous route as described previously17
Neutralization of lethality in mice was carried out according to Tan et al.39 with slight modification from Ramos-Cerrillo et al.40. Briefly, a challenge dose (higher than LD100) of the venom constituting 5xLD50 for B. candidus (or 2xLD50 for N. naja) in 50 μl saline was pre-incubated at 37 °C for 30 min with varying dilutions of the horse antisera/antivenom in normal saline, to give a total volume of 250 μl.
The venom-antiserum or venom-antivenom mixture was then injected into the caudal vein of mice (n = 4–5, 20–25 g). Food and water were provided ad libitum. The results (dead/alive) after 12, 24, 36 and 48 h were recorded. The median effective dose (ED50) was determined as the volume of antiserum (µl) that protected 50% of the challenged mice tested. The in vivo median effective ratios (ER50s) of the antiserum and antivenom were also calculated as described by Morais et al.41.
The in vitro neutralization potency assays of antisera against B. candidus and Vins antivenom against N. naja using nAChR-PSNT binding assay
Optimal concentrations/conditions of nAChR, rat anti-nAChR antibody and goat-anti-rat HRP conjugate used in binding to NK3 coated plate
The format of the basic assay for binding of purified nAChR to the NK3 immobilized microtiter plate was performed as described previously by Ratanabanangkoon et al.13. Optimal concentrations of NK3, nAChR, rat anti-nAChR antibody and goat-anti-rat-HRP conjugate were established13 and used in the experiments that followed. The IC50 values were reported as mean ± standard deviation based on findings from three independent experiments.
Inhibition of nAChR binding to the NK3 coated plate by B. candidus or N. naja venom
The ability of B. candidus and N. naja venoms to individually inhibit the binding of nAChR to NK3 coated plate were expressed as IC50 (venom concentration inhibiting 50% of the nAChR binding or the median inhibitory concentration). In this experiment, B. candidus or N. naja venom at various concentrations were pre-incubated (25 °C, 1 hr) with a fixed and optimal concentration of nAChR. The mixture was then added to the NK3-coated plate and incubated at 25 °C for 1 h. This was followed by additions of rat anti-nAChR serum at 1:1,600 dilution and incubated at 25 °C for 1 h. Thereafter, 1:4,500 diluted goat-anti-rat-HRP conjugate (Abcam®) was added and the mixture was incubated for 60 min at 25 oC. A parallel experiment, in which purified NK3 was used as the reference standard in place of the venom, was carried out. The concentration of the venom used in the pre-incubation experiment that blocked 50% of the nAChR binding to the immobilized NK3 was the IC50 of that venom.
Inhibition of the B. candidus or N. naja venom binding to nAChR by horse antisera against B. candidus or Vins F(ab′)2 antivenom against N. naja
The horse antiserum/antivenom potency (in vitro ED50) was determined using the above described format. Varying amounts of horse sera (0.94 nl – 0.96 µl) were pre-incubated with a fixed amount (5xIC50) of B. candidus venom for 1.5 hr at 37 °C in a total volume of 480 µl. This was called ‘Pre-incubation 1′. The antibody-toxin complexes together with the free antibodies were removed from the mixture by filtration through a 100 kDa MWCO ultrafiltration membrane (Amicon®). The free venom PSNTs and β-neurotoxins in the filtrates (126 µl) were then allowed to react with an optimal amount of nAChR (14 µl) at 25oC for 1 hr; this was called ‘Pre-incubation 2’. The mixtures were transferred to the NK3 coated microtiter wells, followed by the rat anti-nAChR antibody, goat-anti-rat HRP conjugate, etc. The reaction products were then measured. Background control consisted of wells containing non-immune horse serum instead of antisera.
For the N. naja venom and Vins F(ab′)2 antivenom pair, similar and parallel experiment was carried out. In this case, 3xIC50 of crude N. naja venom was incubated with various dilutions of the Vins antivenom and the reaction mixture from the ‘Pre-incubation 1′ was ultrafiltered using a 50 kDa MWCO membrane (Amicon®). The filtrate was then incubated with nAChR in the ‘Pre-incubation 2′ and processed as described above.
The percentage of nAChR binding was calculated from the following equation:
‘OD max’ were the binding of nAChR (optimal amount) without pre-incubation with the venom or antiserum.
‘OD Ag control’ were the binding of nAChR after pre-incubation with 5xIC50 of B. candidus venom (or 3xIC50 of N. naja venom) and without antiserum/antivenom in ‘Pre-incubation 1′.
‘OD sample’ were the binding of nAChR to the NK3 immobilized plate after the nAChR (optimal amount) was pre-incubated with filtrate from ‘Pre-incubation 1′.
The dose–response curves of horse sera volumes versus percent of nAChR binding were then constructed. The in vitro neutralizing activities (ED50s) were the volumes of horse antiserum at which the nAChR binding was blocked by 50 percent compared to wells incubated with non-immune horse sera in place of antisera. The in vitro median effective ratio, ER50, represented µg venom/µl antiserum that inhibited 50% of nAChR binding was calculated.
Similar manipulation and calculations were carried out with the results obtained from the Vins F(ab′)2 antivenom and N. naja venom pair.
Ethics approval
The protocol of animal study on mice followed the Council for International Organizations of Medical Sciences (CIOMS) guidelines and was approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Malaya (Ethical clearance No. 2014–09–11/PHAR/R/TCH).
Miscellaneous procedures
The concentration of protein was determined as described by Lowry et al.42 and by BCA Protein assay Kit (PierceTM) using bovine serum albumin as the standard. GraphPad Prism 5.0 program was used to determine the IC50 and ED50 values. Linear regression with GraphPad Prism 5.0 software was employed in the correlation analysis. Briefly, the correlation coefficient R was determined from the linear regression model, and R2 is the square of the correlation coefficient. An R2 of 0.8–1.0 indicates that the regression line fits well the data in correlation. The statistical significance of the correlation test was set at p < 0.05.
References
Kasturiratne, A. et al. The global burden of snakebite: a literature analysis and modelling based on regional estimates of envenoming and deaths. PLOS medicine 5, 218 (2008).
Chippaux, J. P. Snake-bites: appraisal of the global situation. Bull World Health Organ 76, 515–524 (1998).
Williams, D. J. et al. Ending the drought: new strategies for improving the flow of affordable, effective antivenoms in Asia and Africa. Journal of Proteomics 74, 1735–1767 (2011).
World Health Organization. WHO Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins, 2nd ed.; World Health Organization: Geneva, Switzerland, 2017; In press.
WHO. Guideline for the management of snake-bites (World Health Organization, Geneva, 2016).
Maduwage, K., Silva, A., O’Leary, M. A., Hodgson, W. C. & Isbister, G. K. Efficacy of Indian polyvalent snake antivenoms against Sri Lankan snake venoms: lethality studies or clinically focussed in vitro studies. Sci Rep 6, 26778 (2016).
Silva, A., Hodgson, W. C. & Isbister, G. K. Antivenom for Neuromuscular Paralysis Resulting From Snake Envenoming. LID - E143 [pii] LID -https://doi.org/10.3390/toxins9040143 Toxins (Basel) 9 (2017).
Theakston, R. D. The application of immunoassay techniques, including enzyme-linked immunosorbent assay (ELISA), to snake venom research. Toxicon 21, 341–352 (1983).
Maria, W. S., Cambuy, M. O., Costa, J. O., Velarde, D. T. & Chavez-Olortegui, C. Neutralizing potency of horse antibothropic antivenom. Correlation between in vivo and in vitro methods. Toxicon 36, 1433–1439 (1998).
Rial, A., Morais, V., Rossi, S. & Massaldi, H. A new ELISA for determination of potency in snake antivenoms. Toxicon 48, 462–466 (2006).
Ibrahim, N. M. & Farid, N. M. Comparison between Two In Vitro ELISA-Based Assays in the Determination of Antivenom Potency. Journal of Applied Sciences Research 5, 7 (2009).
Barbosa, C. F., Rodrigues, R. J., Olortegui, C. C., Sanchez, E. F. & Heneine, L. G. Determination of the neutralizing potency of horse antivenom against bothropic and crotalic venoms by indirect enzyme immunoassay. Braz J Med Biol Res 28, 1077–1080 (1995).
Ratanabanangkoon, K. et al. A novel in vitro potency assay of antisera against Thai Naja kaouthia based on nicotinic acetylcholine receptor binding. Sci Rep 7, 8545 (2017).
Petras, D., Heiss, P., Sussmuth, R. D. & Calvete, J. J. Venom Proteomics of Indonesian King Cobra, Ophiophagus hannah: Integrating Top-Down and Bottom-Up Approaches. J Proteome Res 14, 2539–2556 (2015).
Harris, J. B. & Scott-Davey, T. Secreted phospholipases A2 of snake venoms: effects on the peripheral neuromuscular system with comments on the role of phospholipases A2 in disorders of the CNS and their uses in industry. Toxins (Basel) 5, 2533–2571 (2013).
Rowan, E. G. What does beta-bungarotoxin do at the neuromuscular junction? Toxicon 39, 107–118 (2001).
Ratanabanangkoon, K. et al. A Simple and Novel Strategy for the Production of a Pan-specific Antiserum against Elapid Snakes ofAsia. PLOS neglected tropical diseases 10, 1–20 (2016).
Sintiprungrat, K. et al. A comparative study of venomics of Naja naja from India and Sri Lanka, clinical manifestations and antivenomics of an Indian polyspecific antivenom. Journal of Proteomics 132, 131–143 (2016).
Abe, T., Alema, S. & Miledi, R. Isolation and characterization of presynaptically acting neurotoxins from the venom of Bungarus snakes. Eur J Biochem 80, 1–12 (1977).
Pungercar, J. & Krizaj, I. Understanding the molecular mechanism underlying the presynaptic toxicity of secreted phospholipases A2. Toxicon 50, 871–892 (2007).
Spokes, J. W. & Dolly, J. O. Complete purification of beta-bungarotoxin. Characterization of its action and that of tityustoxin on synaptosomal accumulation and release of acetylcholine. Biochim Biophys Acta 596, 81–93 (1980).
Kini, R. M. & Doley, R. Structure, function and evolution of three-finger toxins: mini proteins with multiple targets. Toxicon 56, 855–867 (2010).
Dufton, M. J. & Hider, R. C. Structure and pharmacology of elapid cytotoxins. Pharmacol Ther 36, 1–40 (1988).
Tan, N. H., Wong, K. Y. & Tan, C. H. Venomics of Naja sputatrix, the Javan spitting cobra: A short neurotoxin-driven venom needing improved antivenom neutralization. Journal of Proteomics 157, 18–32 (2017).
Wong, K. Y., Tan, C. H. & Tan, N. H. Venom and Purified Toxins of the Spectacled Cobra (Naja naja) from Pakistan: Insights into Toxicity and Antivenom Neutralization. Am J Trop Med Hyg 94, 1392–1399 (2016).
Fernandez, J. et al. Venomic and antivenomic analyses of the Central American coral snake, Micrurus nigrocinctus (Elapidae). J Proteome Res 10, 1816–1827 (2011).
Rosso, J. P., Vargas-Rosso, O., Gutierrez, J. M., Rochat, H. & Bougis, P. E. Characterization of alpha-neurotoxin and phospholipase A2 activities from Micrurus venoms. Determination of the amino acid sequence and receptor-binding ability of the major alpha-neurotoxin from Micrurus nigrocinctus nigrocinctus. Eur J Biochem 238, 231–239 (1996).
Alape-Giron, A. et al. Characterization of multiple nicotinic acetylcholine receptor-binding proteins and phospholipases A2 from the venom of the coral snake Micrurus nigrocinctus. FEBS Lett 380, 29–32 (1996).
Ranawaka, U. K., Lalloo, D. G. & de Silva, H. J. Neurotoxicity in snakebite–the limits of our knowledge. PLoS Negl Trop Dis 7, e2302 (2013).
Dufton, M. J. & Harvey, A. L. Dendrotoxins: How Does Structure Determine Function? Journal of Toxicology: Toxin Reviews 17, 161–182 (1998).
Harvey, A. L. Twenty years of dendrotoxins. Toxicon 39, 15–26 (2001).
Laustsen, A. H., Lomonte, B., Lohse, B., Fernandez, J. & Gutierrez, J. M. Unveiling the nature of black mamba (Dendroaspis polylepis) venom through venomics and antivenom immunoprofiling: Identification of key toxin targets for antivenom development. JJournal of Proteomics 119, 126–142 (2015).
Tan, C. H., Tan, K. Y., Lim, S. E. & Tan, N. H. Venomics of the beaked sea snake, Hydrophis schistosus: A minimalist toxin arsenal and its cross-neutralization by heterologous antivenoms. Journal of Proteomics 126, 121–130 (2015).
Tan, C. H., Wong, K. Y., Tan, K. Y. & Tan, N. H. Venom proteome of the yellow-lipped sea krait, Laticauda colubrina from Bali: Insights into subvenomic diversity, venom antigenicity and cross-neutralization by antivenom. Journal of Proteomics 166, 48–58 (2017).
Gutierrez, J. M. et al. Snakebite envenoming. Nat Rev Dis Primers 3, 17079 (2017).
WHO. Progress in the characterization of venoms and standardization of antivenoms (World Health Organization, Geneva, 1981).
Harrison, R. A., Hargreaves, A., Wagstaff, S. C., Faragher, B. & Lalloo, D. G. Snake envenoming: a disease of poverty. PLoS Negl Trop Dis 3, e569 (2009).
Karlsson, E., Arnberg, H. & Eaker, D. Isolation of the principal neurotoxins of two Naja naja subspecies. Eur J Biochem 21, 1–16 (1971).
Tan, C. H., Liew, J. L., Tan, K. Y. & Tan, N. H. Assessing SABU (Serum Anti Bisa Ular), the sole Indonesian antivenom: A proteomic analysis and neutralization efficacy study. Scientific Reports 6, 37299, https://doi.org/10.1038/srep37299 (2016).
Ramos-Cerrillo, B. et al. Characterization of a new polyvalent antivenom (Antivipmyn Africa) against African vipers and elapids. Toxicon 52, 881–888 (2008).
Morais, V., Ifran, S., Berasain, P. & Massaldi, H. Antivenoms: potency or median effective dose, which to use? Journal of Venomous Animals and Toxins including Tropical Diseases 16, 191–193 (2010).
Lowry, O. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J. Protein measurement with the Folin phenol reagent. J Biol Chem 193, 265–275 (1951).
Acknowledgements
This study was supported by a research grant (no. IM 2011-01 to KR) from the Chulabhorn Research Institute and a research grant from the University of Malaya (grant no. BKS003–2017 to CHT). The authors are deeply grateful to Dr James M Dubbs, Associate Professor Dr. Sirida Youngchim, Dr. Lawan Chanhome of the Queen Soavabha Memorial Institute and Mr. Sutat Lapanan for their kind assistance and suggestions.
Author information
Authors and Affiliations
Contributions
K.R. and C.H.T designed the study; P.S., K.P., K.Y.T., B.C. and S.E. undertook the laboratory studies; K.Y.T., C.H.T., K.R. and P.S. undertook the analysis; K.R and C.H.T. drafted the manuscript and all authors contributed to the final version; K.R. and C.H.T. are responsible for the manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ratanabanangkoon, K., Simsiriwong, P., Pruksaphon, K. et al. An in vitro potency assay using nicotinic acetylcholine receptor binding works well with antivenoms against Bungarus candidus and Naja naja. Sci Rep 8, 9716 (2018). https://doi.org/10.1038/s41598-018-27794-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-27794-3
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
