
Abstract
Lindera, a core genus of the Lauraceae family, has important economic uses in eastern Asia and North America. However, its historical diversification has not been clarified. In this study, we report nine newly sequenced Lindera plastomes. The plastomes of these nine Lindera species range from 152,211 (L. nacusua) to 152,968 bp (L. metcalfiana) in length, similar to that of another Lauraceae species, Litsea glutinosa (152,618 bp). The length variation of these plastomes derived from the length variation in the loci ycf1, ycf2, ψycf1, and ndhF-ψycf1. Comparing our sequences with other available plastomes in the Lauraceae indicated that eight hypervariable loci, ihbA-trnG, ndhA, ndhF-rpl32, petA-psbJ, psbK-psbI, rps16, trnS-trnG, and ycf1, could serve as DNA barcodes for species delineation, and that the inverted repeats (IRs) showed contraction/expansion. Further phylogenetic analyses were performed using 32 complete plastomes of Lauraceae and seven barcodes from 14 additional species of Lindera and related species in the core Lauraceae. The results showed that these Lindera species grouped into two or four sub-clades, and that two Litsea species and Laurus nobilis were located in the same sub-clade as five Lindera species. These data support a close relationship between the genera Laurus, Lindera, and Litsea, and suggest that Lindera is polyphyletic.
Similar content being viewed by others

Introduction
In plants, the chloroplast is the main locus of photosynthesis and carbon fixation1. The chloroplast (cp) genome ranges from 120 to 180 kb in size and has a characteristic structure, in which two inverted repeat (IR) regions divide the cp genome into four parts: the IRs themselves, a large single copy region (LSC) and a small single copy region (SSC)2. In the LSC, three of the four core plant barcodes, psbA-trnH, rbcL, and matK, have been widely used for identification purposes and phylogenetic analyses in the past twenty years3. Currently, complete cp genomes as well as full-length cp gene sequences are available for an increasing number of taxa, and both have been the subjects of numerous phylogenies. Phylogenomics, a technique for estimating phylogenetic relationships based on high-throughput sequencing, can allow a comprehensive understanding of the evolutionary history of organisms. For instance, Ma, et al.4 used cp phylogenomics to resolve the deep-level relationships of Arundinarieae. Yang, et al.5 used complete cp genome sequences to infer phylogenetic relationships in the genus Quercus. Very recently, Zhang, et al.6 provided important insights into deep phylogenetic relationships and the diversification history of the Rosaceae based on analyzing plastid phylogenomics.
Like Arundinarieae and Rosaceae, Lauraceae are a large monophyletic group, comprising approximately 3,500 known species from over 50 genera worldwide7,8, which are by far the largest family of the order Laurales9. The phylogenetic backbone is well resolved by now, but problems remain within most major clades10,11,12,13. Multiple classification schemes based on a variety of morphological and anatomical characteristics have been proposed, but none has been fully accepted. Therefore, other sources of data, such as genomic information, are needed for classification. As a foundation for further studies of Lauraceae phylogenomics, here we focus on Lindera14, a genus belonging to the core Lauraceae in the sense of Rohwer and Rudolph15, or the core Laureae in the sense of Chanderbali, et al.11 Lindera species have not only popular ornamental and economic uses but also great medicinal and therapeutic value. Lindera is widely distributed in tropical, subtropical, and temperate zones in Asia and North America and includes approximately 100 species16, with Lindera umbellata Thunb. as the type species. The fruits of most Lindera species, particularly Lindera communis Hemsley and Lindera glauca (Siebold & Zucc.) Blume, are rich in fatty oils and thus represent important wild woody oil plants17,18,19. Lindera megaphylla Hemsley is an economically important small deciduous tree, the wood of which can be used for buildings and furniture. Moreover, it is also a courtyard greening species, and its trunk and leaves are rich in alkaloids20,21. More importantly, Lindera plants are widely used in traditional medicine. Lindera obtusiloba Blume has been used as a traditional medicine for the treatment of fever, abdominal pain, extravasation, inflammation and poor blood circulation22,23,24,25. Thus, molecular methods for species delineation in the genus of Lindera are of considerable interest.
The first reported cp genomic markers in the Lauraceae were rbcL and trnL-trnF, which were used for phylogenetic analysis of the Laurales9,26. The matK gene was used to construct a phylogenetic tree to analyse the relationships among Lauraceae genera10, but in this analysis the Laureae (represented by a single species of Actinodaphne, Laurus, Lindera, Litsea and Neolitsea) remained unresolved. Then, Chanderbali, et al.11 constructed larger phylogenetic trees for the Lauraceae using the chloroplast sequences trnL-trnF, psbA-trnH, trnT-trnL, and rpl16 as well as the nuclear barcoding markers 26S rDNA and internal transcribed spacer (ITS) rDNA. The result of their trnL-trnF + psbA-trnH analysis showed Lindera erythrocarpa as sister to Litsea glaucescens, albeit without bootstrap support, whereas in their ITS analysis Lindera erythrocarpa appeared as sister to Laurus nobilis, likewise without bootstrap support. Li, et al.12 and Nie, et al.27, who used ITS plus different chloroplast markers, found that the genus Lindera was not monophyletic. Fijridiyanto and Murakami28 further analysed the phylogenetic relationships of Litsea and its related genera using the nuclear marker rpb2, also finding that Lindera was not a monophyletic group. Most recently, a report29 showed a close relationship between the genera Lindera and Litsea while using rbcL, matK, trnH–psbA, and ITS to investigate the phylogenetic relationships in the Lauraceae.
In this study, we report the complete chloroplast genome sequences of nine Lindera species chosen for their economic importance. Lindera communis, L. glauca, Lindera latifolia Hook. f.14, and Lindera nacusua (D. Don) Merr14 are important wild woody oil plants, L. megaphylla and Lindera robusta (C. K. Allen) H. P. Tsui14 are important timber plants, and Lindera benzoin (L.) Blume30, Lindera metcalfiana var. dictyophylla (C. K. Allen) H. P. Tsui14, and L. obtusiloba are widely used in traditional medicine. In addition, compared to other Lauraceae species, L. benzoin and L. obtusiloba are distributed in more northern areas, and their wide distribution shows the ability to adapt to cold environments. Based on cp sequence information, the characteristics and phylogenetic information of these species were further investigated.
Results
Characteristics of the cp genomes of Lindera
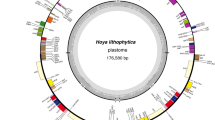
The sizes of the cp genomes of the nine Lindera species range from 152,211 (L. nacusua) to 152,968 bp (L. metcalfiana) (Table 1). The sequences were assembled into a single, circular, double-stranded DNA sequence for each species. The cp genomes have a typical quadripartite structure, comprising the LSC, with a length of 93,573 (L. benzoin) to 93,888 bp (L. metcalfiana), the SSC, with a length of 18,336 (L. nacusua) to 18,978 bp (L. metcalfiana), and a pair of IR copies of 20,048 (L. benzoin) to 20,075 bp (L. obtusiloba) in length (Fig. 1 and Table 1). The cp sequences of Lindera communis, L. glauca, L. latifolia, L. megaphylla, L. metcalfiana, L. obtusiloba, and L. robusta are larger than that of Litsea glutinosa (152,618 bp, GenBank accession number KU382356)8, but shorter than those of Phoebe omeiensis and P. sheareri (152,855 bp, GenBank accession number KX427772; 152,876 bp, GenBank accession number KX427773)31,32. The cp sequences of Lindera benzoin and L. nacusua are all shorter than those of Litsea glutinosa, Phoebe omeiensis and P. sheareri (Table 1).

Circular gene map of Lindera species (Lindera benzoin, L. communis, L. glauca, L. latifolia, L. megaphylla, L. metcalfiana, L. nacusua, L. obtusiloba, and L. robusta) chloroplast genomes. The genes lying outside each circle are transcribed counter-clockwise, while those inside are transcribed clockwise. The coloured bars indicate different functional groups. The dashed darker grey area in the inner circle indicates genome GC content, while the lighter grey area shows AT content. IR = inverted repeat; SSC = small single copy; LSC = large single copy.
All nine Lindera cp genomes contain 113 single-copy genes, among which 79 encode proteins. Sixteen genes have one intron (atpF, ndhA, ndhB, petB, petD, rpl2, rpl16, rpoC1, rps12, rps16, trnA-UGC, trnG-UCC, trnI-GAU, trnK-UUU, trnL-UAA and trnV-UAC), and two genes have two introns (clpP and ycf3) (Table 2). The ycf1 and ycf2 genes are trans-spliced, and the nucleotide sequences of the ycf1 and ycf2 pseudogenes are 1,372, 1,373, 1,376, 1,377, 1,379, 1,383, and 1,389 bp (Lindera communis, L. megaphylla, L. metcalfiana, L. latifolia and L. nacusua, L. robusta, L. glauca, L. benzoin, and L. obtusiloba), and 3,162 bp (all nine Lindera species), respectively, being truncated at the IR boundaries (Fig. 1). Additionally, the ycf15 gene, located in the LSC of the nine Lindera genomes, is also a pseudogene (Table 2). The GC content of these Lindera cp genomes is 39.2%, except for L. obtusiloba (39.1%), which is similar to that of Litsea glutinosa (39.2%), but slightly higher than those of Phoebe omeiensis and P. sheareri (39.1%). The GC content of L. obtusiloba is the same as those of Phoebe omeiensis and P. sheareri32 (Table 1).
Identification of the most variable regions
To elucidate levels of sequence divergence, we calculated the nucleotide variability (Pi) values. The Pi values within 600 bp across the nine genomes vary from 0 to 0.0187, and the mean value is 0.0048 (Fig. 2A), indicating that these sequences have high similarity. However, we identified nine hypervariable loci (Pi > 0.014), which are ihbA-trnG, ndhA, ndhF-rpl32, petA-psbJ, psbK-psbI, rps16, trnS-trnG, and ycf1. The ndhA and ndhF-rpl32 loci are in the SSC region; ihbA-trnG, petA-psbJ, psbK-psbI, rps16, and trnS-trnG are in the LSC region; and ycf1 is in the IR region (Fig. 2A). To investigate the levels of sequence divergence among the genera, we calculated the genetic divergence of the sequenced cp genomes of core Lauraceae, including Alseodaphne, Cinnamomum, Laurus, Lindera, Litsea, Machilus, Nectandra, Persea, Phoebe, and Sassafras. The Pi values vary from 0 to 0.0201 in these 29 sequences (Fig. 2B), indicating that the variation at genus level is significantly larger than that at the species level. All these genomic features are shown in the sequence alignment of the nine Lindera species, Laurus nobilis, and Litsea glutinosa (Fig. S1). According to the alignment results (Fig. S1), all of these species share the same order and orientation of syntenic blocks, indicating that no rearrangement occurred in gene organization. These results accord with those of Male, et al.33 and Asif, et al.34, which illustrate that cp genomes tend to be conserved and perfectly collinear, especially in the same plant family.

Comparision of the nucleotide variability (Pi) values of the nine Lindera plastomes (A) and 29 plastomes of the core Lauraceae (B). X axis: position of the midpoint of a window, Y axis: nucleotide diversity of each window.
Comparative analysis of cp genomes
Size variation in cp genomes is partly a result of contraction and expansion at the borders of the IR regions1. To trace the size differences among Lindera cp genomes, the IR-LSC and IR-SSC boundaries, with full annotations for the adjacent genes, were re-examined across nine Lindera cp genomes (Fig. 3). The entire ycf1 gene crosses the SSC/IRB boundary, while another fragment of ψycf1 is located at the IRA/SSC boundary. A ψycf1 fragment with a length of 1,372–1,389 bp was found in the IRA region because the boundary between the SSC and IRB extended into the ycf1 gene. In the nine Lindera species, the distances between ψycf1 and ndhF vary from 6 (L. obtusiloba) to 37 bp (Litsea glutinosa) in length (Fig. 3).

Comparison of LSC, IR, and SSC junction positions among nine Lindera chloroplast genomes.
Phylogenomic analysis of sequenced Lauraceae plastomes
To determine the phylogenetic relationships of the nine Lindera species with other Lauraceae, we reconstructed a phylogenetic tree based on our nine complete cp genomes of Lindera and 23 fully sequenced cp genomes of related Lauraceae (Fig. 4). The tree shows that the Lindera species can be divided into two sub-clades. Sub-clade I (Lindera benzoin, L. latifolia, L. metcalfiana, L. obtusiloba, and L. robusta) is sister to sub-clade II, which contains the remaining species. Sub-clade I, however, has only 78% bootstrap support, whereas sub-clade II is 100% supported. Within sub-clade I, part I (Lindera communis, L. glauca, and L. nacusua) is sister to part II (Laurus nobilis, Lindera megaphylla, and Litsea glutinosa) (Fig. 4). These data indicate that (among the species investigated) Laurus nobilis and Litsea glutinosa are most closely related to Lindera megaphylla, and the Lindera group, which is most closely related to the Cinnamomum-Ocotea clade, comprises nine Lindera species, Laurus nobilis and Litsea glutinosa.

Molecular phylogenetic tree of 32 species of Lauraceae based on complete plastome sequences. Numbers at the nodes represent bootstrap percentages.
Phylogenetic analysis of Lindera species
To better understand the phylogenetic relationships between our sequenced nine Lindera species and the other Lindera taxa with reported barcoding data, we downloaded available sequences from GenBank including rbcL, matK, trnL-trnF, psbA-trnH, ndhF, ITS and rpb2 of all 33 core Lauraceae species (Table S1). The result of this phylogenetic analysis supports the grouping of all Lindera species with three Litsea species and Laurus nobilis (Fig. 5). This tree is further divided into four main sub-clades. Sub-clade I (Bayesian inference posterior probability, BI-PP = 1.00) includes Lindera communis, L. fragrans, L. glauca, L. megaphylla, L. nacusua, plus Laurus nobilis, Lindera glutinosa, and Litsea tomentosa. Sub-clade II includes only Lindera obtusiloba. Sub-clade III (BI-PP = 1.00) includes Lindera erythrocarpa, L. latifolia, L. longipedunculata, L. lucida, L. metcalfiana, L. polyantha, L. robusta, and Litsea cubeba. Sub-clade IV (BI-PP = 1.00) includes Lindera aggregata, L. benzoin, L. chunii, L. fruticosa, L. kariensis, L. pulcherrima, L. reflexa, L. triloba, L. umbellata, and L. villipes.

Phylogenetic relationships of 33 core Lauraceae species based on concatenated sequences of seven loci. Numbers at the nodes represent Bayesian inference posterior probabilities (BI-PP).
Discussion
In this study, the complete cp genomes of nine Lindera species were sequenced using Illumina sequencing technology. These nine cp genomes possess the typical angiosperm quadripartite structure, which has a large single copy (LSC) region and a small single copy (SSC) region, separated by two short inverted repeat (IR) regions (Fig. 1). Similar to those of other sequenced Lauraceae species, the nine Lindera cp genomes have less length variation in the IR regions but more variation in the LSC and SSC regions. The length of the IR regions ranges from 20,048 bp in L. benzoin to 20,075 bp in L. obtusiloba. As Song, et al.35 reported, both IR regions of Machilus balansae and M. yunnanensis are 20,074 bp in length. The LSC region of Lindera metcalfiana is 93,888 bp in length, which is 315 bp larger than that of L. benzoin. The SSC region of L. metcalfiana is 18,978 bp in length, which is 642 bp larger than that of L. nacusua. A comparative analysis was conducted to explain these size differences,and the results suggest the following: the length of the entire ycf1 gene ranges from 5,550 bp in L. glauca to 5,574 bp in L. megaphylla; the length of the truncated ycf1 gene ranges from 1,372 bp in L. communis and L. megaphylla to 1,389 bp in L. obtusiloba; the length of the entire ycf2 gene ranges from 6,837 bp in L. metcalfiana to 6,858 bp in L. obtusiloba (Fig. 3). It has been reported that the ycf1 and ycf2 genes are located in the boundaries between the IR regions and the LSC and SSC regions, and these two genes experienced incomplete duplication35,36. Thus, length changes in ycf1, ycf2, ψycf1, and ndhF-ψycf1 drive the contraction and expansion of the IR regions in the cp genomes of Lindera.
The ycf1 was identified as a hypervariable locus at the species level within Lindera, as were ihbA-trnG, ndhA, ndhF-rpl32, petA-psbJ, psbK-psbI, rps16, and trnS-trnG. At the genus level, we identified ndhF-rpl32 and ycf1 as variable regions among 29 core Lauraceae species from Alseodaphne, Cinnamomum, Laurus, Lindera, Litsea, Machilus, Nectandra, Persea, Phoebe, and Sassafras. Among these regions, ndhF-rpl32, and ycf1 have been shown to be variable enough across seed plants for use as barcodes in plant taxonomy and phylogeny31,35,37,38,39,40. The four fragments ndhF-rpl32, petA-psbJ, trnS-trnG, and ycf1, have previously been identified as hypervariable regions in Litsea glutinosa, Machilus balansae, M. yunnanensis, Persea americana, Phoebe omeiensis, and P. sheareri8,31,33,35. In addition to the previous results, we found that the fragments ihbA-trnG, ndhA, petA-psbJ, psbK-psbI, rps16, and trnS-trnG seemed to be especially variable loci in Lindera plastomes, and they showed promising levels of variation for application in DNA barcoding or intraspecific studies.
Previously, hypervariable loci have been used as barcoding markers for taxon identification and phylogenetic analysis3,38,41. The chloroplast gene fragments rbcL, matK, and psbA-trnH and the nuclear internal transcribed spacer (ITS) have been reported as available markers for plant barcoding3,29,38,41,42. Our phylogenetic analysis using 17 complete cp genome sequences, five chloroplast regions (rbcL, matK, trnL-trnF, psbA-trnH, and ndhF), and two nuclear sequences (ITS and rpb2) of another 14 Lindera and two Litsea species shows that a group containing 23 Lindera species, three Litsea species, and Laurus nobilis was most closely related to a Cinnamomum-Ocotea clade, with strong support11,43,44,45, as in previously published phylogenetic trees. The two species Litsea glutinosa and L. tomentosa as well as Laurus nobilis are located in a sub-clade with five Lindera species, Lindera communis, L. fragrans, L. glauca, L. megaphylla, and L. nacusua, while Litsea cubeba was located in another sub-clade with seven Lindera species, Lindera erythrocarpa, L. latifolia, L. longipedunculata, L. lucida, L. metcalfiana, L. polyantha, and L. robusta, which is in agreement with a previous phylogenetic result by Fijridiyanto and Murakami28 who defined the relationships among seven Lindera species. And the clade containing Laurus, Lindera, and Litsea was the sister group of the Cinnamomum-Ocotea clade containing Cinnamomum, Nectandra, and Sassafras species, as found in previous studies11,43,44,45. In our study, we further determined the relationships of 16 additional Lindera species, Lindera benzoin, L. chunii, L. communis, L. fragrans, L. fruticosa, L. kariensis, L. latifolia, L. longipedunculata, L. megaphylla, L. metcalfiana, L. nacusua, L. pulcherrima, L. reflexa, L. robusta, L. triloba, and L. villipes. The results of our study are fully compatible with those of Fijridiyanto and Murakami28, as far as well-supported clades are concerned. A topological difference is found in the placement of Lindera glauca, in a clade (with Actinodaphne and Neolitsea, not examined here) that is sister to all taxa examined in both studies in Fijridiyanto and Murakami, vs. in sub-clade I here. This sub-clade, however, has practically no support.
Overall, our study reports nine chloroplast genomes of Lindera for the first time and compares their organizations with those of other Lauraceae species. Four divergent regions were found at the genus level, providing a valuable source of markers for future studies on species delineation and resolution of phylogenetic relationships among the Lauraceae. Our phylogenomic results also suggest that plastid phylogenomics can be regarded as a robust method for tackling difficult phylogenies and could be extended across the genera Lindera and Litsea with additional systematic sampling.
Materials and Methods
Plant materials
Nine Lindera species were used for this research. Fresh young leaves of Lindera benzoin, L. communis, L. glauca, L. latifolia, L. megaphylla, L. metcalfiana, L. nacusua, L. obtusiloba, and L. robusta were harvested from mature plants growing in botanical gardens (Table 3) and then immediately frozen in liquid nitrogen and stored at −80 °C. Specimens were deposited in the herbarium of the Biodiversity Research Group (BRG) of Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences (CAS).
DNA preparation and chloroplast sequencing
The cetyltrimethylammonium bromide (CTAB) method was used to extract total genomic DNA. Next-generation sequencing was performed according to Yang, et al.46, and nine universal primer pairs from their study were also taken to perform long-range PCR. Then, the PCR products were purified and combined. Following the manufacturer’s instructions (Illumina Nextera XT library), the mixture was fragmented and used to construct 500-bp short-insert libraries. All nine complete Lindera cp genomes were sequenced using a Genome Analyzer (Illumina HiSeq2000) at the Germplasm Bank of Wild Species, Kunming Institute of Botany, CAS.
Cp genome assembly and annotation
All nine Lindera raw sequencing reads were filtered using the NGS QC Tool Kit to obtain high-quality short reads. Then, the raw reads were trimmed according to their quality, removing bases from the 5′ and 3′ ends until no base with Q < 20 was found. After that, the chloroplast genome was de novo assembled using the trial version of CLC v.8 (http://www.qiagenbioinformatics.com). The contigs were aligned using the publicly available cp genome of Litsea glutinosa8 in Geneious 4.8 (http://www.geneious.com/) as a reference. Dual Organellar GenoMe Annotator (DOGMA, http://dogma.ccbb.utexas.edu/) software was used to annotate the cp genomes and identify genes encoding proteins47, transfer RNAs (tRNAs), and ribosomal RNAs (rRNAs). The OrganellarGenomeDRAW tool (OGDRAW, http://ogdraw.mpimp-golm.mpg.de/) was used to draw the genome maps of Lindera benzoin, L. communis, L. glauca, L. latifolia, L. megaphylla, L. metcalfiana, L. nacusua, L. obtusiloba, and L. robusta.
Sliding window analysis to identify hypervariable regions
MAFFT (http://mafft.cbrc.jp/alignment/server) was used to align the nine cp genomes with one another. Afterward, we manually adjusted these sequences using BioEdit software (http://www.mbio.ncsu.edu/bioedit/bioedit.html). DnaSP version 5.0 with a sliding window analysis was used to calculate the nucleotide variability values (π) within the chloroplast genomes. The window length was set to 600 bp and the step size to 200 bp. Then, the R program was used to plot values. In addition, we used this method to analyse the hypervariable regions among the nine Lindera cp genomes and those of Alseodaphne gracilis (GenBank accession number MG407593)48, A. huanglianshanensis (GenBank accession number MG407594)48, A. semecarpifolia (GenBank accession number MG407595)48, Cinnamomum camphora (GenBank accession number LC228240)49, C. kanehirae (GenBank accession number KR014245)50, C. micranthum (GenBank accession number KR014245)50, C. verum (GenBank accession number KY635878)9, Laurus nobilis (GenBank accession number KY085912)13, Lindera glauca (GenBank accession number MF188124), Litsea glutinosa (GenBank accession number KU382356)8, Machilus balansae (GenBank accession number KT348517)35, M. pauhoi (GenBank accession number MH178403), M. thunbergii (GenBank accession number MH178404), M. yunnanensis (GenBank accession number KT348516)35, Nectandra angustifolia (GenBank accession number MF939340)13, Persea americana (GenBank accession number KX437771)31, P. chekiangensis (GenBank accession number KY346511), Phoebe omeiensis (GenBank accession number KX437772)31,32, P. sheareri (GenBank accession number KX437773)31,32, P. zhennan (GenBank accession number KY346512), and Sassafras tzumu (GenBank accession number MF939339)13.
Phylogenetic analyses
The plastome sequences of Alseodaphne gracilis, A. huanglianshanensis, A. semecarpifolia, Cinnamomum camphora, C. kanehirae, C. micranthum, C. verum, Endiandra discolor (GenBank accession number KT588615)51, E. globosa (GenBank accession number KT588614)51, Laurus nobilis, Lindera glauca, Litsea glutinosa, Machilus balansae, M. pauhoi, M. thunbergii, M. yunnanensis, Nectandra angustifolia, Persea americana, Phoebe chekiangensis, P. omeiensis, P. sheareri, P. zhennan, and Sassafras tzumu were downloaded from the NCBI GenBank. Thereafter, we used MAFFT to align these 23 sequences and our nine Lindera sequences and manually edited where necessary with BioEdit software. After these steps, the jModelTest 2.0 program was used to calculate nucleotide substitution, and the results indicated that the optimal model was “GTR + G”52. The “GTR + G” model was used for all maximum likelihood (ML) analyses implemented in RAxML version 8.0.20, as suggested in the manual53. Nonparametric bootstrapping was performed with the “fast bootstrap” algorithm of RAxML and 1000 replicates. The cp genomes of Endiandra discolor and E. globosa were used as the outgroup.
We assembled sequence alignments for 33 taxa of Lauraceae, 17 (Cinnamomum camphora, C. kanehirae, C. micranthum, C. verum, Laurus nobilis, Lindera benzoin, L. communis, L. glauca, L. latifolia, L. megaphylla, L. metcalfiana, L. nacusua, L. obtusiloba, L. robusta, Litsea glutinosa, Nectandra angustifolia, and Sassafras tzumu) with complete cp genome sequences, plus ITS and rpb2 sequences, and 14 Lindera species (Lindera aggregata, L. chunii, L. erythrocarpa, L. fragrans, L. fruticosa, L. kariensis, L. longipedunculata, L. lucida, L. polyantha, L. pulcherrima, L. reflexa, L. triloba, L. umbellata, and L. villipes), and two Litsea species (Litsea cubeba and L. tomentosa) with the DNA sequences of rbcL, matK, trnL-trnF, psbA-trnH, ndhF, ITS54 and rpb2 from GenBank (Table S1). MAFFT software was used to analyse these sequences, and incongruous sequences of the same species were removed. The seven-sequence matrix was then manually adjusted, and Sequencher 4.10 (http://www.genecodes.com) was used to merge identical sequences. Then, a joint matrix was constructed using Sequence Matrix v.1.7.8, and jModelTest 2.0 was used to calculate the nucleotide substitution. The optimal model was chosen (“GTR + I + G”) (freqA = 0.3003, freqC = 0.1988, freqG = 0.1919, freqT = 0.3090, R(a) [AC] = 0.8998, R(b) [AG] = 2.0890, R(c) [AT] = 0.2648, R(d) [CG] = 0.4178, R(e) [CT] = 1.9183, R(f) [GT] = 1.0000, p-inv = 0.7020, gamma shape = 0.3000) to construct the phylogenetic tree52. Phylogenetic relationships were reconstructed using Bayesian inference (BI) and ML methods in MrBayes version 3.1.255.
Data archiving statement
The complete cp genome sequence data of the nine Lindera taxa have been submitted to the GenBank of NCBI. The GenBank accession numbers are MH220730 (Lindera benzoin), MH220731 (L. communis), MH220732 (L. glauca), MH220733 (L. latifolia), MH220734 (L. megaphylla), MH220735 (L. metcalfiana), MH220736 (L. nacusua), MH220737 (L. obtusiloba), and MH220738 (L. robusta).
References
Zhao, Y. B. et al. The complete chloroplast genome provides insight into the evolution and polymorphism of Panax ginseng. Frontiers in Plant Science 5, 696, https://doi.org/10.3389/fpls.2014.00696 (2014).
Palmer, J. D. Comparative organization of chloroplast genomes. Annual Review of Genetics 19, 325–354, https://doi.org/10.1146/annurev.ge.19.120185.001545 (1985).
Hollingsworth, P. M. et al. A DNA barcode for land plants. Proceedings of the National Academy of Sciences of the United States of America 106, 12794–12797, https://doi.org/10.1073/pnas.0905845106 (2009).
Ma, P. F., Zhang, Y. X., Zeng, C. X., Guo, Z. H. & Z., L. D. Chloroplast phylogenomic analyses resolve deep-level relationships of an intractable bamboo Tribe Arundinarieae (Poaceae). Systematic Biology 63(6), 933–950, https://doi.org/10.1093/sysbio/syu054 (2014).
Yang, Y. C. et al. Comparative analysis of the complete chloroplast genomes of five Quercus species. Frontiers in Plant Science 7, 959, https://doi.org/10.3389/fpls.2016.00959 (2016).
Zhang, S. D. et al. Diversification of Rosaceae since the Late Cretaceous based on plastid phylogenomics. The New Phytologist 214, 1355–1367, https://doi.org/10.1111/nph.14461 (2017).
Rohwer, J. G. Lauraceae. In: Kubitzki, K., Rohwer, J. G. & Bittrich, V. (eds) The Families and Genera of VascularPlants, vol. 2. Springer-Verlag, Berlin, Heidelberg, New York, London, Paris, Tokyo, Hong Kong, Barcelona, Budapest, ISBN 3-540-55509-9 and 0-387-55509-9, https://doi.org/10.1007/978-3-662-02899-5_46 (1993).
Hinsinger, D. D. & Strijk, J. S. Toward phylogenomics of Lauraceae: The complete chloroplast genome sequence of Litsea glutinosa (Lauraceae), an invasive tree species on Indian and Pacific Ocean islands. Plant Gene 9, 71–79, https://doi.org/10.1016/j.plgene.2016.08.002 (2017).
Renner, S. S. Circumscription and phylogeny of the Laurales: evidence from molecular and morphological data. American Journal of Botany 86(9), 1301–1315 (1999).
Rohwer, J. G. Toward a phylogenetic classification of the Lauraceae: evidence from matK sequences. Systematic Botany 25(1), 60–71, https://doi.org/10.2307/2666673 (2000).
Chanderbali, A. S., van der Werff, H. & Renner, S. S. Phylogeny and historical biogeography of Lauraceae: evidence from the chloroplast and nuclear genomes. Annals of the Missouri Botanical Garden 88, 104–134, https://doi.org/10.2307/2666133 (2001).
Li, J., Christophel, D. C., Conran, J. G. & Li, H.-W. Phylogenetic relationships within the ‘core’ Laureae (Litsea complex, Lauraceae) inferred from sequences of the chloroplast gene matK and nuclear ribosomal DNA ITS regions. Plant Systematics and Evolution 246, 19–34, https://doi.org/10.1007/s00606-003-0113-z (2004).
Song, Y. et al. Evolutionary comparisons of the chloroplast genome in Lauraceae and insights into loss events in the Magnoliids. Genome Biology and Evolution 9, 2354–2364, https://doi.org/10.1093/gbe/evx180 (2017).
Cui, H. B. & Weff, H. 5. LINDERA Thunberg, Nov. Gen. Pl. 64. 1783, nom. cons., not Adanson (1763). Flora of China 7, 142–159 (2008).
Rohwer, J. G. & Rudolph, B. Jumping genera: the phylogenetic positions of Cassytha, Hypodaphnis, and Neocinnamomum (Lauraceae) based on different analyses of trnK intron sequences. Annals of the Missouri Botanical Garden 92(2), 153–178 (2005).
Cao, Y. et al. The genus Lindera: a source of structurally diverse molecules having pharmacological significance. Phytochemistry Reviews 15, 869–906, https://doi.org/10.1007/s11101-015-9432-2 (2015).
Zhou, Z. Y., Wang, W. G., Duan, J. F., Zhu, L. W. & Jiang, J. X. Chinese spicehush (Lindera communis) seed oil catalyzed into biodiesel by enzymatic method. Applied Mechanics and Materials 448–453, 1587–1591, https://doi.org/10.4028/www.scientific.net/AMM.448-453.1587 (2013).
Wang, W.-G., Ma, L., Jiang, J.-X., Jiang, L. & Zhang, Z.-X. Transesterified Chinese spicehush (Lindera communis) seed oil as a biodiesel fuel. Forestry Studies in China 9, 132–136, https://doi.org/10.1007/s11632-007-0020-4 (2007).
Niu, J. et al. Transcriptome analysis of distinct Lindera glauca tissues revealed the differences in the unigenes related to terpenoid biosynthesis. Gene 559, 22–30, https://doi.org/10.1016/j.gene.2015.01.002 (2015).
Chou, C. J., Lin., L. C., Chen., K. T. & Chen, C. F. Northalifoline, a new isoquinolone alkaloid from the pedicels of Lindera megaphylla. Journal of Natural Products 57(6), 689–694 (1994).
Chen, C. C., Lin, C. F. & Huang, Y. L. Bioactive constituents from the flower buds and peduncles of Lindera megaphylla. Journal of Natural Products 59(9), 1426–1425 (1995).
Choi, H. G. et al. A new neolignan and lignans from the stems of Lindera obtusiloba Blume and their anti-allergic inflammatory effects. Archives of Pharmacal Research 37, 467–472, https://doi.org/10.1007/s12272-013-0239-7 (2014).
Hong, C. O., Lee, H. A., Rhee, C. H., Choung, S. Y. & Lee, K. W. Separation of the antioxidant compound quercitrin from Lindera obtusiloba Blume and its antimelanogenic effect on B16F10 melanoma cells. Bioscience, Biotechnology, and Biochemistry 77, 58–64, https://doi.org/10.1271/bbb.120562 (2013).
Freise, C., Kim, K. Y. & Querfeld, U. A Lindera obtusiloba extract blocks calcium-/phosphate-induced transdifferentiation and calcification of vascular smooth muscle cells and interferes with matrix metalloproteinase-2 and metalloproteinase-9 and NF-κB. Evidence-based Complementary and Alternative Medicine: eCAM 2015, 679238, https://doi.org/10.1155/2015/679238 (2015).
Ruehl, M. et al. Extracts of Lindera obtusiloba induce antifibrotic effects in hepatic stellate cells via suppression of a TGF-β-mediated profibrotic gene expression pattern. The Journal of Nutritional Biochemistry 20, 597–606, https://doi.org/10.1016/j.jnutbio.2008.06.003 (2009).
Renner, S. S. Phylogenetic affinities of Monimiaceae based on cpDNA gene and spacer sequences. Perspectives in Plant Ecology, Evolution and Systematics 1, 61–77 (1998).
Nie, Z. L., Wen, J. & Sun, H. Phylogeny and biogeography of Sassafras (Lauraceae) disjunct between eastern Asia and eastern North America. Plant Systematics and Evolution 267, 191–203, https://doi.org/10.1007/s00606-007-0550-1 (2007).
Fijridiyanto, I. A. & Murakami, N. Phylogeny of Litsea and related genera (Laureae-Lauraceae) based on analysis ofrpb2 gene sequences. Journal of Plant Research 122, 283–298, https://doi.org/10.1007/s10265-009-0218-8 (2009).
Liu, Z. F. et al. DNA barcoding evaluation and implications for phylogenetic relationships in Lauraceae from China. Plos One 12, e0175788, https://doi.org/10.1371/journal.pone.0175788 (2017).
Tucker, O., Maciarello, M. J., Burbage, P. W. & George., S. Spicebush [Lindera benzoin (L.) Blume var. benzoin, Lauraceae]: a tea, spice, and medicine. Economic Botany 48(3), 333–336 (1994).
Song, Y., Yao, X., Tan, Y., Gan, Y. & Corlett, R. T. Complete chloroplast genome sequence of the avocado: gene organization, comparative analysis, and phylogenetic relationships with other Lauraceae. Canadian Journal of Forest Research 46, 1293–1301, https://doi.org/10.1139/cjfr-2016-0199 (2016).
Song, Y. et al. Comparative analysis of complete chloroplast genome sequences of two subtropical trees, Phoebe sheareri and Phoebe omeiensis (Lauraceae). Tree Genetics & Genomes 13, https://doi.org/10.1007/s11295-017-1196-y (2017).
Male, P. J. et al. Genome skimming by shotgun sequencing helps resolve the phylogeny of a pantropical tree family. Molecular Ecology Resources 14, 966–975, https://doi.org/10.1111/1755-0998.12246 (2014).
Asif, M. H. et al. Complete sequence and organisation of the Jatropha curcas (Euphorbiaceae) chloroplast genome. Tree Genetics & Genomes 6, 941–952, https://doi.org/10.1007/s11295-010-0303-0 (2010).
Song, Y. et al. Comparative analysis of complete chloroplast genome sequences of two tropical trees Machilus yunnanensis and Machilus balansae in the family Lauraceae. Frontiers in Plant Science 6, 662, https://doi.org/10.3389/fpls.2015.00662 (2015).
Li, P. et al. Comparative genomics and phylogenomics of East Asian tulips (Amana, Liliaceae). Frontiers in Plant Science 8, 451, https://doi.org/10.3389/fpls.2017.00451 (2017).
Korotkova, N., Nauheimer, L., Ter-Voskanyan, H., Allgaier, M. & Borsch, T. Variability among the most rapidly evolving plastid genomic regions is lineage-specific: implications of pairwise genome comparisons in Pyrus (Rosaceae) and other Angiosperms for marker choice. Plos One 9, e112998, https://doi.org/10.1371/journal.pone.0112998 (2014).
Dong, W. P. et al. ycf1, the most promising plastid DNA barcode of land plants. Scientific Reports 5, 8348, https://doi.org/10.1038/srep08348 (2015).
Dong, W. P., Liu., J., Yu., J., Wang., L. & L., Z. S. Highly variable chloroplast markers for evaluating plant phylogeny at low taxonomic levels and for DNA barcoding. Plos One 7(4), e35071, https://doi.org/10.1371/journal.pone.0035071.t001 (2012).
Sarkinen, T. & George, M. Predicting plastid marker variation: can complete plastid genomes from closely related species help? Plos One 8, e82266, https://doi.org/10.1371/journal.pone.0082266 (2013).
Dong, W. P. et al. Discriminating plants using the DNA barcode rbcLb: an appraisal based on a large data set. Molecular Ecology Resources 14, 336–343, https://doi.org/10.1111/1755-0998.12185 (2014).
Rohwer, J. G. et al. Is Persea (Lauraceae) monophyletic? Evidence from nuclear ribosomal ITS sequences. Taxon 58(4), 1153–1167 (2009).
Huang, J. F. et al. Origins and evolution of cinnamon and camphor: A phylogenetic and historical biogeographical analysis of the Cinnamomum group (Lauraceae). Molecular phylogenetics and evolution 96, 33–44, https://doi.org/10.1016/j.ympev.2015.12.007 (2016).
Trofimov, D., Rudolph, B. & Rohwer, J. G. Phylogenetic study of the genus Nectandra (Lauraceae), and reinstatement of Damburneya. Taxon 65(5), 980–996, https://doi.org/10.12705/655.3 (2016).
Rohde, R. et al. Neither Phoebe nor Cinnamomum – the tetrasporangiate species of Aiouea (Lauraceae). Taxon 66(5), 1085–1111, https://doi.org/10.12705/665.6 (2017).
Yang, J. B., Li, D. Z. & Li, H. T. Highly effective sequencing whole chloroplast genomes of angiosperms by nine novel universal primer pairs. Molecular Ecology Resources 14, 1024–1031, https://doi.org/10.1111/1755-0998.12251 (2014).
Wyman, S. K., Jansen, R. K. & Boore, J. L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 20(17), 3252–3255, https://doi.org/10.1093/bioinformatics/bath352. PMID:15180927 (2004).
Song, Y., Yao, X., Liu, B., Tan, Y. & Corlett, R. T. Complete plastid genome sequences of three tropical Alseodaphne trees in the family Lauraceae. Holzforschung 72, 337–345, https://doi.org/10.1515/hf-2017-0065 (2018).
Liu, Z., Chen, S. L., Song, J. Y., Zhang, S. J. & Chen, K. L. Application of deoxyribonucleic acid barcoding in Lauraceae plants. Pharmacognosy magazine 8, 4–11, https://doi.org/10.4103/0973-1296.93301 (2012).
Wu, C. C., Chu, F. H., Ho, C. K., Sung, C. H. & Chang, S. H. Comparative analysis of the complete chloroplast genomic sequence and chemical components of Cinnamomum micranthum and Cinnamomum kanehirae. Holzforschung 71(3), 189–197, https://doi.org/10.1515/hf-2016-0133 (2017).
Rossetto, M., Kooyman, R., Yap, J. Y. & Laffan, S. W. From ratites to rats: the size of fleshy fruits shapes species’ distributions and continental rainforest assembly. Proceedings. Biological Sciences 282, 20151998, https://doi.org/10.1098/rspb.2015.1998 (2015).
Darriba, D., Taboada, G. L., Doallo, R. & Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nature Methods 9, 772, https://doi.org/10.1038/nmeth.2109 (2012).
Stamatakis, A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313, https://doi.org/10.1093/bioinformatics/btu033 (2014).
Li, L., Li, J., Conran, J. G., Li, X. W. & Li, H. W. Phylogeny of Neolitsea (Lauraceae) inferred from Bayesian analysis of nrDNA ITS and ETS sequences. Plant Systematics and Evolution 269, 203–221, https://doi.org/10.1007/s00606-007-0580-8 (2007).
Ronquist, F. & Huelsenbeck, J. P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19, 1572–1574, https://doi.org/10.1093/bioinformatics/btg180 (2003).
Acknowledgements
The authors would like to thank Dr. Bing Liu from the State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, for providing plant tissues of Lindera benzoin and L. glauca, and acknowledge Jun-Bo Yang, Jing Yang, Juan-Hong Zhang, Chun-Yan Lin, Zheng-Shan He, and Ji-Xiong Yang at the Germplasm Bank of Wild Species, Kunming Institute of Botany, Chinese Academy Sciences, for sequencing technology. This work was supported by the CAS “Light of West China” Programme (No. Y7XB061B01), the project of the Southeast Asia Biodiversity Research Institute (No. Y4ZK111B01), and the National Natural Science Foundation of China (No. 31600531). We sincerely thank two anonymous referees for their critical and invaluable comments that greatly improved our manuscript.
Author information
Authors and Affiliations
Contributions
Mei-Li Zhao and Yu Song designed the experiments; Mei-Li Zhao, Yun-Hong Tan and Yu Song collected the samples; Mei-Li Zhao conducted the experiments; Mei-Li Zhao, Jun Ni and Xin Yao analysed the results; and Mei-Li Zhao, Yu Song and Zeng-Fu Xu wrote the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhao, ML., Song, Y., Ni, J. et al. Comparative chloroplast genomics and phylogenetics of nine Lindera species (Lauraceae). Sci Rep 8, 8844 (2018). https://doi.org/10.1038/s41598-018-27090-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-27090-0
This article is cited by
-
Genome size, ploidy estimates, and leaf morphology of temperate Lindera (Lauraceae) cultivated in North America
Genetic Resources and Crop Evolution (2024)
-
Complete chloroplast genome sequence and phylogenetic analysis of Rhododendron molle G. Don, an endangered Ericaceae species located on Dabie Mountains (central China)
Plant Biotechnology Reports (2023)
-
A comparative analysis of complete chloroplast genomes of seven Ocotea species (Lauraceae) confirms low sequence divergence within the Ocotea complex
Scientific Reports (2022)
-
The complete chloroplast genome of Stauntonia chinensis and compared analysis revealed adaptive evolution of subfamily Lardizabaloideae species in China
BMC Genomics (2021)
-
Uncovering dynamic evolution in the plastid genome of seven Ligusticum species provides insights into species discrimination and phylogenetic implications
Scientific Reports (2021)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
