
Abstract
In order to improve the phylogeography of the male-specific genetic traces of Greek and Phoenician colonizations on the Northern coasts of the Mediterranean, we performed a geographically structured sampling of seven subclades of haplogroup J in Turkey, Greece and Italy. We resequenced 4.4 Mb of Y-chromosome in 58 subjects, obtaining 1079 high quality variants. We did not find a preferential coalescence of Turkish samples to ancestral nodes, contradicting the simplistic idea of a dispersal and radiation of Hg J as a whole from the Middle East. Upon calibration with an ancient Hg J chromosome, we confirmed that signs of Holocenic Hg J radiations are subtle and date mainly to the Bronze Age. We pinpointed seven variants which could potentially unveil star clusters of sequences, indicative of local expansions. By directly genotyping these variants in Hg J carriers and complementing with published resequenced chromosomes (893 subjects), we provide strong temporal and distributional evidence for markers of the Greek settlement of Magna Graecia (J2a-L397) and Phoenician migrations (rs760148062). Our work generated a minimal but robust list of evolutionarily stable markers to elucidate the demographic dynamics and spatial domains of male-mediated movements across and around the Mediterranean, in the last 6,000 years.
Similar content being viewed by others


Introduction
Views on the formation of the European gene pool have recently undergone a remarkable increase in complexity, by virtue of genomic data, new methods of analysis, new correlations with archaeological data, and the genotyping of individuals from different temporal strata. A general conceptual framework is that of multiple genetic layers, sequentially stratified as a result of migrations, admixtures, resettlements and replacements. Furthermore, the impact of each of these processes turns out to differ dramatically in different regional areas of the continent1,2,3,4,5,6,7,8. In this context, the contact zones between Europe, Western Asia and Northern Africa have emerged as crucial for the departure, movement and arrival of migrants9. The water bodies, too, are now regarded less of a barrier and more of a bridge from the Mesolithic onwards10,11,12,13,14,15. On the background of the pre-Neolithic occupation of Southern Europe, characterized by low population density and strong divergence, coastal and immediate inland areas offered resources for successful settlement and trade, especially for the conveyors of cultural, technological and warfare advances. This may have promoted population growth, admixture with and absorption of pre-existing groups, and more widespread occupation of inhabitable and exploitable territories16.
A case study is represented by the Southern half of the Italian peninsula. Its genetic distinctiveness is undisputed, and initially attributed to the Greek founders of Magna Graecia in the 1st millennium BC17. Subsequent works18,19,20,21,22,23,24 reconsidered this hypothesis, leaving the relative contributions by seafarers of the early Neolithic, other inputs in the intervening period, and Greeks of Magna Graecia still unresolved. Yet, Southern Italy offered appropriate conditions for successful settlement, demographic growth and hence molecular radiation. The presence of the expected corresponding signatures can directly be tested by a phylogeographic approach, as applicable to non-recombining portions of the genome. A suitable temporal resolution can be attained, provided that the number of variants is high enough to give a faithful record of a rapid succession of mutational events25. This power is not currently attained by massive autosomal SNP data, in which modern patterns of arrangement in a multidimensional space were already established by at least the middle/late Neolithic1,2.
The Y chromosome haplogroup J (Hg J) has long been considered the clearest marker of East-to-West migrations that impacted South and South-eastern Europe26,27,28,29. Our final goal was to determine the source population(s), the relative contribution(s), the routes and the Italian locations where the genetic traces of the above processes are prominent today. This required a deeper understanding of the phyletic affinities between Hg J chromosomes found across the Northern Mediterranean, and of the dynamics of the population(s) to which the Hg J carriers belonged.
We reasoned that lineages ancestral to the Southern Italian (I) ones should be found in one or both of two candidate source populations: Turkey (T) and Greece (G). A topology with Turkish and Greek lineages generally closer than the Italian ones (I,(G,T)), would denote a peculiarity of ancestry in Italy. A mainly (G,(T,I)) topology would be in line with a stronger contribution of Anatolian/Middle Eastern migrants. Finally, a mainly (T,(G,I)) topology would be consistent with a Greek heritage in Southern Italy.
Hence, we used Next Generation Sequencing (NGS) to obtain a large and ascertainment-independent list of variants from 7 subclades within Hg J, each sampled in more than one copy in Turkey, Greece and Italy. This strategy proved successful in providing a first list of evolutionarily stable markers of Greek and Phoenician colonists, but also shed light on demographic processes on a much broader area around the Mediterranean.
Results
Resequencing of 4.4 Mb of Y chromosome in 58 subjects resulted in approximately 2.7 Mb of high quality positions, of which 1079 turned out to be variable among subjects. In our series, four positions displayed an alternative allele different from that reported in dbSNP (https://www.ncbi.nlm.nih.gov/snp) and two a single alternative allele at an otherwise triallelic position (Supplemental Table 2).
When compared with studies addressing targeted regions or the complete chromosome, a remarkable proportion of our SNPs (625/1079 = 58%) turned out to be novel, despite all four previous studies included a considerable number of Hg J carriers (Supplemental Fig. 5). Also, 642 of our variable positions were not recorded in dbSNP 147 (Supplemental Table 2). Both observations indicate that our sampling scheme captured a poorly known quota of Hg J diversity.
Features of the Hg J tree
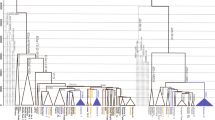
The topological relationships among the 7 selected subclades in the MP tree (Fig. 1 and Supplemental Fig. 3), recapitulated the known Hg J phylogeny, with a first split into J1 and J2. The internal topology of J1 was strongly influenced by a single deeply rooted sample (branch 1) and a subclade (J1-P56, branch 6) clustering a Yemenite and two Ethiopian subjects. A multifurcation grouped together all remaining subjects sampled as J1-M267 (and found to carry J1-P58) in a vast area spanning from Italy to Africa and the Arabian Peninsula.

Unrooted MP tree of 58 Hg J chromosomes based on 1079 variable positions (Supplemental Table 2), with the root placed based on the reference sequence. The seven groupings used in the initial selection of samples are shown on the right. The provenance of subjects is abbreviated for Italy (I), Greece (G) and Turkey (T), or in full for other locations. The markers for the definition of clades and selection of samples are shown next to the branches where they occur, in grey boxes. The number of SNPs defining each branch (branch length) is also reported. Black lozenges indicate nodes discussed in the text, for which specific SNP assays were applied.
Within J2, an early split separated J2a-M410 from J2b-M12. Within this latter, both J2b-M205 (5 subjects) and J2b-M241 (3 subjects) were sampled.
Within J2a-M410, a lineage characterized by the ancestral YCAIII microsatellite pattern with >18 repeats26 separated early. In view of the phylogenetic equivalence with L26/Page55, this lineage was indicated also as J2a(xL26).
Among carriers of the derived YCAIII pattern ≤18 repeats, a single subject (FOG05) was found to define a deep branch, while the remaining subjects defined 5 fast radiating lineages. One of these (branch 60) leads to the selected J2a-L397 clade (8 subjects). Subjects selected as carriers of L397 (associated with the 6 repeats allele at DYS445) were further split by 3 variants.
In the clade defined by M67, three sister lineages were sampled, one of which leading to J2a-M92. Finally, within the J2a-M92 clade one variant (branch 98) clustered 4 of the 6 subjects.
The assignment of SNPs to each of the tree branches (Supplemental Table 2) revealed that, as expected, most of the variants not previously annotated belonged to private branches (Supplemental Fig. 3). However, their distribution greatly contributed to the definition of previously poorly known lineages, i.e. 1) a small clade within J1-P58 (branch 18); 2) a small clade within J2b-M205 (branch 29)30,31; 3) a clade (branch 41) grouping 4 of the 6 J2a(xL26)30,32; 4) three branches within J2a-L397, one of which previously unknown (branch 58); 5) a small clade (branch 62) grouping 3 subjects; 6) a branch within J2a-M92 (branch 98), not reported in any of the previous studies. It is worth noting that all the above small clades grouped subjects from at least 2 of our 3 broad sampling areas. Thus, the low representation of these clades is not due to a geographically insufficient sampling but, rather, to a low frequency despite a large area of occurrence.
Another main aspect of the tree shown in Fig. 1 replicated the peculiarity of Hg J as opposed to other haplogroups in refs31,32,33,34,35,36 (hereafter “previous landmark studies”) and others, i.e. the large variations in branch length and the common occurrence of very deep lineages as compared to the entire haplogroup. Even between sister lineages in sub-terminal branches, large variations in the accumulation of variants were observed. We tested37 the equality of branch lengths and obtained non significant departures from the null, either across the entire tree (Supplemental Fig. 2B), or within J2 and J1 (not shown). In addition, seven branches which appeared as private were equal or longer than 25 mutational steps (Fig. 1). All these features were paralleled by the weak evidence for star-like radiations.
Finally, a main topological characteristic of the tree was immediately at odds with the simple idea that the radiation within Hg J occurred during a progressive westward dispersal of people from Turkey (Anatolia) towards the Aegean Islands, Greece and Italy26,27,28,38: when looking at each of the 7 selected subclades or their internal sub-branches, no evidence for a preferential occurrence of the (T,(G,I)) topology emerged, despite the spatially-specific sampling scheme.
Dating and inferences on demography
We obtained node ages using an ancient specimen sequenced at relatively high coverage, chosen from within Hg J. The calibrated mutation rate fell within the range of values reported in recent studies and obtained with a variety of methods39,40. The dates obtained here for some key nodes of the tree (Supplemental Fig. 4) were in close agreement with those of previous studies. Two temporal windows showed an enrichment of nodes, i.e. 16–14 kya and 6-3 kya. Moreover, within J2a(xL26), J2a-L26(xM67), -M67(xM92) and -M92 a paucity of nodes was observed from 5 kya to present. The Bayesian reconstruction returned a population growth rate as low as 1.4% per generation (95% C.I. 0.6–2.9%)41.
In order to gain insights on the demography of population(s) of which the Hg J carriers are reporters, we generated Bayesian Skyline Plots for the entire tree and some lineages (Supplemental Fig. 5). The whole tree displayed two phases of growth, at 15 kya and 5 kya onward, respectively. Interestingly, this plot showed a slowdown of growth in the last 2.5 kya, replicating the Greek and Turkish results, which differ from the pattern of at least 12 other European populations42.
The above analysis should be considered with caution, as far as the number of chromosomes in each of the 7 selected clades was not necessarily proportional to their frequency in the population. We then repeated the analysis for 5 groups of chromosomes devoid of nested subclades also used in the selection process (e.g. J2a-M92 within J2a-M67). Both J1-M267 and J2b-M12 displayed a sudden increase of population size around 4–5 kya, followed by stasis. J2a(xL26) showed a progressive increase until 5 kya, again followed by stasis. Finally, J2a-L397 and J2a-M92 did not show changes during their respective ages.
Closing on recent growth episodes
The above results returned the picture of Hg J as a collection of ancient lineages surviving within the population at a very modest growth. The resequencing of 18 chromosomes from Southern Italy, where the impact of the Greeks has been proposed17 to dominate the genetic landscape, failed to show the signs of massive growth in any of the selected clades. In these circumstances, we sought to locate more subtle signs of growth, by spotting candidate tree branches and searching subjects falling into them, by means of genotyping. We pinpointed seven variants which could potentially unveil star clusters of sequences (lozenges in Fig. 1). We directly genotyped candidate carriers of these variants and complemented our results with data from resequenced chromosomes (see Materials and Methods).
Within J1-M267 we obtained 32 carriers of the derived allele at rs751613702 (branch 18). They turned out to be sampled at locations (Fig. 2A) as distant as Tuscany (Italy) and Pakistan. Their centroid was significantly shifted to the East as compared to the carriers of the parental lineage (J1-M267). Moreover, the home range of derived alleles was entirely within that of parental lineage, with shorter pairwise distances (t-test p < 1E-4). Further within the same clade, we assayed also rs760148062 (branch 17). We found 14 carriers of the derived allele (Fig. 2A). In this case the distances separating the carriers of the derived allele and their home range denote movements over wider distances than those of the carriers of the ancestral allele (p < 1E-8). In fact 47% (8/17) of the ancestral alleles were confined in the Israeli Bedouins. This difference in migratory history is also witnessed by STRs (Fig. 2A). The large cluster of J1-M267 immediately ancestral to rs751613702 and enriched in Middle Easterners, corresponded to the 14-23-10-11-12 motif at DYS19-390-391-392-393, which matches the haplotypes PCS2+ and PCS4+ providing a Phoenician Colonization Signal28. Subjects derived at rs751613702 but ancestral at rs760148062 mostly had DYS19(14)-DYS393(13), i.e. a combination not listed as PCS. Conversely, subjects derived at rs760148062 retained the 14-23-10-11-12 motif.

Maps of sampling locations for the carriers of the derived allele (white triangle point down) at the indicated SNP vs carriers of the ancestral allele (black triangle point-up), conditioned on identical genotype at the same most terminal marker. Coastlines were drawn with the R packages “map” and “mapproj” v. 3.1.3 (https://cran.r-project.org/web/packages/mapproj/index.html), and symbols added with default functions. (A) J1; (B) J2-L397. The star indicates the centroid of derived alleles. The solid square indicates the centroid of ancestral alleles, with its 95% C.I. (ellipse). In the insets: distributions of the pairwise sampling distances (in Km) for the carriers of the ancestral (black) and derived (white) allele, with solid and dashed lines indicating the respective averages. At right: median joining network of 7-STR haplotypes and SNPs in the same groups, with sectors coloured according to sampling location. Haplotype structure is detailed for some nodes, in the order YCA2a-YCA2b-DYS19-DYS390-DYS391-DYS392-DYS393 (in italics).
Within J2b-M12 (Supplemental Fig. 7), we obtained 14 carriers of the derived allele at rs779180992 (branch 29), which resides within J2b-M205. The centroid of the carriers of the parental lineage (J2b-M12) was shifted to the West as compared with the overall sample, in line with the high frequency of this allele in Albania and Western Greece. The carriers of the derived allele at rs779180992 had a centroid significantly shifted to the East, being found in Italy, the Balkan Peninsula, Northern Turkey, Caucasus and in two Middle Easterners, but neither in Sardinia nor Crete. Their sampling distances did not differ (p = n.s.) from the carriers of the ancestral allele. In the STR network, chromosomes derived at rs779180992 did not show any central haplotype, and were variable at all loci but DYS393.
Within J2a-M92 (Supplemental Fig. 7) we genotyped position Y: 8237851 (branch 98). The derived allele was found in 13 subjects of our core sample but in none of the previous landmark studies. The centroid of the carriers of the ancestral alleles was shifted to the West as compared to the overall sample, in line with the enrichment of J2a-M92 in the Western part of our sample26. The carriers of the derived allele at Y:8237851 had a centroid significantly shifted to the East, being found in Italy (including Sardinia), but also in Turkey and the Eastern Mediterranean (including Crete and Cyprus). STRs indicated that the mutation at Y:8237851 occurred on a 22-19-14-22-10-11-12 haplotype (YCA2a, YCA2b-DYS19-390-391-392-393), found only in Italy and Sardinia, and radiating in Italy and elsewhere.
Within J2a-L397, we genotyped the three variants defining branches 57, 58 and 59 (Fig. 2B). Only 4 samples were ancestral at all three positions (paragroup J2a-L397*). For each of the three positions, the centroids of carriers of the ancestral allele were located in Greece. The overall distribution of J2a-L397 covered only the northern Mediterranean, with no carriers of this lineage in Cretans of our core sample, but reported at 2.6% by in ref.43.
For branch 57 (rs773856887), derived alleles (20) were found at the outer edges of the overall distribution of the carriers of J2a-L397 (p < 1E-7). The STRs network indicated a probable radiation from a 22-19-15-23-9-11-12 haplotype.
The distribution of derived alleles (6) for branch 58 (rs775512766) was strikingly different, with 5 of the 6 instances found in Southern Italy. This concentration resulted in very low pairwise sampling distances (p < 1E-8). Derived alleles were associated with both the 22-19-15-23-9-11-12 and the 22-19-15-25-9-11-12 STR haplotypes. Branches 57 and 58 shared the feature of a centroid for derived alleles shifted to the West as compared to ancestral alleles, though not significantly.
Derived alleles for branch 59 (21) were spread from Italy to Turkey (including Cyprus), covering distances similar, on average, to ancestral alleles (p = n.s.). Here, the STR haplotypes radiated from the founding type to 22-19-16-23-9-11-12 and 22-19-15-24-9-11-12.
Overall, 10/13 of the subjects sampled in a single Italian location (Locri 16.2E, 38.2 N) carried J2a-L397* (as branch 57, 58, or 59), representing 18.5% of all J2a-L397 in the whole sample.
Discussion
In order to improve our understanding of the patrilinear relationships among populations on the northern coasts of the Mediterranean we addressed specifically Hg J. We used a straightforward spatial representation of sampling locations for a composite sample generated in-house and supplemented with published data. Its composition and distribution is heavily dependent on the particular set of examined populations, on the number of subjects studied for each of them and the local Hg J frequency. We derived our approach from ref.44, and calculated the position of centroids and pairwise sampling distances, which are strongly contingent on the sample composition, too. Thus, only comparisons between different partitions of our dataset are meaningful.
Within seven subclades of Hg J we performed a geographically structured sampling and encountered, among others, a number of lineages previously poorly known. We confirmed that J1 contains a number of rare lineages which are diversified and deeply rooted. They have been sampled in Greece, Turkey, the Caucasus and North of it30,31 and, surprisingly repeatedly, in America31,34. The emergence of these lineages was synchronous (approx. 15 kya) with a similar burst within J2, where several lineages radiated within J2a-M410.
Two features of our tree (Fig. 1) are at odds with the simplistic idea of a dispersal of Hg J as a whole from the Middle East towards Greece and Italy and an accompanying radiation26. First, there is little evidence of sudden diversification between 15 and 5 kya, a period of likely population increase and pressure for range expansion, due to the Agricultural revolution in the Fertile Crescent. Second, within each subclade, lineages currently sampled in Turkey do not show up as preferentially ancestral. Both findings are replicated and reinforced by examining the previous landmark studies. Our Turkish samples do not coalesce preferentially to ancestral nodes when mapped onto these studies’ trees.
Additional relevant information on the entire Hg J comes from the discontinuous distribution of J2b-M12. The northern fringe of our sample is enriched in the J2b-M241 subclade, which reappears in the gulf of Bengal38,45, with low frequencies in the intervening Iraq46 and Iran47. No J2b-M12 carriers were found among 35 modern Lebanese, as contrasted to one of two ancient specimens from the same region35.
In summary, a first conclusion of our sequencing effort and merge with available data is that the phylogeography of Hg J is complex and hardly explained by the presence of a single population harbouring the major lineages at the onset of agriculture and spreading westward. A unifying explanation for all the above inconsistencies could be a centre of initial radiation outside the area here sampled more densely, i.e. the Caucasus and regions North of it, from which different Hg J subclades may have later reached mainland Italy, Greece and Turkey, possibly following different routes and times. Evidence in this direction comes from the distribution of J2a-M41045,48 and the early-49 or mid-Holocene50 southward spread of J1.
We obtained poor evidence for population growth after 5 kya. A mid-Holocene drop in the male effective size was reported31, attributed to the rise of farming and the associated cultural and social habits, which may have increased the variance in offspring number both within and between demes. With the caveat of a equalized sampling across clades, we simply observed a stasis in the 15-5 kya time window in the whole tree, followed by a sudden increase at 5 kya, in line with most European populations42. Of the internal clades, only J1-M267 and J2b-M12 displayed the same growth phase at approximately 5 kya. In the context of the recent explosive increase in world population size41,51, genetically detectable in Y chromosomal studies of all continents31 and Europe42, our inferences raise the question of whether individual lineages can survive without undergoing a proportional growth. Here, it is to be noted that the signal of growth for a whole population derives from the multiple haplogroups that contribute to its gene pool. Some of them may have undergone expansion elsewhere, reaching the regions where we found them today, mixed with Hg J, only later. Social and economic circumstances can also act against male-mediated expansions (compare with ref.25). For example, occupational roles that involve a mainly patrilineal transmission of wealth to a single (usually the eldest) son, and/or active persecution directed towards paternally inherited cultural traits are viable possibilities.
In order to tackle more directly our goal, we focused on some lineages which suggested limited expansions in a context of generalized modest growth in the last 5–6 kya. The most evident expansion within Hg J in this time frame is provided by J1-P58 in the Middle East. Studies on ancient specimens are revealing that Hg J emerged during the Bronze Age in Lebanon and Jordan, with an earlier record in Iran1,35. The increase in frequency46,49 and radiation of the J1-P58 lineage is evident in the clustering of subjects from Middle East and Western Arabian Peninsula in this clade. Here, a possible structuring within the Arabian Peninsula is indicated by a single subject (UAE443; Supplemental Fig. 3) from the Eastern side, carrying a lineage (rs867592041; Supplemental Table 2) distinct from those common in Lebanon, Saudi Arabia and Yemen31,35. Within J1-P58, we identified two nested sublineages whose age frames between 5 and 4.4 kya the partition into a subset (rs751613702derived-rs760148062ancestral) that remained basically resident and another that underwent a strong dispersal (rs760148062derived). The geographic distribution of the latter and the associated STR haplotypes28 make this variant a good candidate for Phoenician migrations52.
The lineage defined by rs779180992, belonging to J2b-M205, and dated at 4–4.5 kya, has a radically different distribution, with derived alleles in Continental Italy, Greece and Northern Turkey, and two instances in a Palestinian and a Jew. The interpretation of the spread of this lineage is not straightforward. Tentative hypotheses are linked to Southward movements that occurred in the Balkan Peninsula from the Bronze Age29,53, through the Roman occupation and later54.
The slightly older (5.6–6.3 kya) branch 98 lineage displays a similar trend of a Eastward positioning of derived alleles, with the notable difference of being present in Sardinia, Crete, Cyprus and Northern Egypt. This feature and the low frequency of the parental J2a-M92 lineage in the Balkans27 calls for an explanation different from the above.
Finally, we explored the distribution of J2a-L397 and three derived lineages within it. J2a-L397 is tightly associated with a typical DYS445 6-repeat allele. This has been hypothesized as a marker of the Greek colonizations in the Mediterranean55, based on its presence in Greek Anatolia and Provence (France), a region with attested Iron Age Greek contribution. All of our chromosomes in this clade were characterized also by DYS391(9), confirming their Anatolian Greek signature. We resolved the J2a-L397 clade to an unprecedented precision, with three internal markers which allow a finer discrimination than STRs. The ages of the three lineages (2.0–3.0 kya) are compatible with the beginning of the Greek colonial period, in the 8th century BCE. The three subclades have different distributions (Fig. 2B), with two (branches 57, 59) found both East and West to Greece, and one only in Italy (branch 58). As to Mediterranean Islands, J2a-L397 was found in Cyprus56 and Crete43. Its presence as one of the three branches 57–59 will represent an important test. In Italy all three variants were found mainly along the Western coast (18/25), which hosted the preferred Greek trade cities. The finding of all three differentiated lineages in Locri excludes a local founder effect of a single genealogy. Interestingly, an important Greek colony was established in this location, with continuity of human settlement until modern times. The sample composed of the same subjects displayed genetic affinities with Eastern Greece and the Aegean also at autosomal markers57. In summary, the distributions of branches 57–59 mirror the variety of the cities of origin and geographic ranges during the phases of the colonization process58.
This work revealed that signs of Holocenic Hg J radiations are subtle, but nevertheless concentrated mainly in the Bronze Age, a critical period for the establishment of genetic structure in Europe25,42. Clear signs of Phoenician legacy have been linked to J228, but we detect them also in J1. We found a SNP marker of the Phoenician colonization within a star-like cluster which emerged in the Middle East and sent representatives also beyond the present-day Iran (Fig. 2A).
Molecular signs of the Greek expansion are even more subtle. A peculiarity of our Greek markers is that they relate more to the Aegean and are poorly represented in mainland Greece. This is line with the archeologically documented two step process of establishment of Aegean colonies and subsequent outward colonizations. It is possible that the markers arose in Greek Anatolia. However the intrusion of northern peoples in Continental Greece has been all but excluded based on genetic evidence7, and may have abated the frequency of these alleles.
We provided here the strongest temporal and distributional evidence so far for markers which can be attributed to the Greek settlement of Magna Graecia. This process cannot be expected to have conveyed only J2a-L397, but other lineages as well: for example E-V1359, which left a clear signal across the Mediterranean28,55. J2a-L397 in Central-Southern Continental Italy accounts for only 13% (24/182) of Hg J. At a face value this can be taken as a bottom figure for the Greek contribution to the gene pool of the region. Though the true value may be higher than this, it is unlikely that the bulk of diversity has a Greek origin. Under these circumstances it is not surprising that unequivocal genetic traces of the Greek colonists in Continental Italy have gone undetected so far19.
Our work generated a minimal but robust list of evolutionarily stable markers to elucidate the demographic dynamics and spatial domains of male-mediated movements across and around the Mediterranean. The search of additional ones will be demanding. In fact, the particular tree topology (long private branches) imposes the search of many variable positions to detect possible geographically-confined groups of samples that could inform on the most recent time of arrival of certain lineages. Nonetheless, the markers explored here appear of great usefulness. First, they can be validated by specifically searching for them in test cases such as Western vs. Eastern Sicily (not sampled here) where distinct Phoenician and Greek cultural and genetic inputs are documented60. Other key locations include the North Tunisian coast (Phoenicians), Western Black Sea, North-Eastern Libyan and Mediterranean Iberian coasts (Greeks). Second, they can be directly addressed in available collections of ancient and modern samples never studied at this level of resolution61.
Methods
The sample
The core sample consisted of 469 males, carriers of Hg J, from collections of the authors, assembled in the 1980’s, 1990’s and 2000’s. The original sampling was performed by colleagues and operators at a number of collaborating Institutions and included the recording of the subject’s place of residence and informed consent. The subject was also asked to report the origin of his parents. Recent immigrants were excluded. Anonymized blood or DNA samples were then received at the corresponding author’s laboratory.
All of these subjects were typed at least for the markers reported in Fig. 1, which define 7 Hg J subclades, 6 of which monophyletic. For each subclade, candidates for the capture-NGS experiment were selected, who satisfied the condition of representing at least all the three broad regions of Italy, Greece and Turkey. Selection considered the amount and quality of DNA, disregarding information on markers other than the six mentioned above. Seven additional individuals entered the experiment, specifically for their provenance (Nepal, Arabian Peninsula and Africa), or a previous assignment to an interesting lineage (J1-P56). Overall, 59 subjects entered the resequencing experiment (Supplemental Table 1).
For further genotyping at phylogeographically interesting markers in the overall sample, 42 additional individuals from the CEPH HGDP panel were also considered, with typing obtained in-house or from the literature45.
For data analysis, 375 additional individuals selected from targeted30 or whole genome studies31,32,33,34,35,36 were also considered, taking into account duplication of some subjects. The overall sample thus consisted of 893 subjects, distributed as shown in Table 1 and Supplemental Fig. 1.
This study received approval by the intramural ethical committee (Comitato Etico Indipendente document n. 164/14, dated Nov. 21, 2014) who expressly considered the list of collaborators, anonymity of samples and the compliance with consent regulations of previous publications which included the same samples. The entire research was performed in accordance with relevant guidelines reported in the above mentioned document and those agreed upon by the scientific community.
Experimental procedures
A detailed description of all methods is provided as Supplemental text. Briefly, a Roche Nimblegen custom array was used to capture a total of about 4.4 Mb, which excluded almost all the repetitive elements from the 22 X-degenerated blocks. A > 50× mean depth was obtained upon Illumina HiSeq. 2500 sequencing. Stringent filtering resulted in exclusion of one subject. Overall, 1079 variable positions (Supplemental Table 2) were detected out of 2,711,986 bp (Supplemental Table 3). Quality control included resequencing of six subjects (Supplemental Table 1), independently examined in a previous study30, and tree statistics (Supplemental Fig. 2). The transition/transversion ratio (Supplemental Table 5) and context dependency among the variants fitted genome-wide patterns62. The maximum parsimony (MP) tree was obtained with MEGA63 from the 1079 × 58 matrix of allele states (Fig. 1 and Supplemental Fig. 3). Node ages (Supplemental Table 7 and Supplemental Fig. 4) based on SNP diversity at 817 positions, were obtained with BEAST64 for the tree including also the Hg J individual “Kotias” (9.720 kya)65. The same program was used to obtain Bayesian Skyline Plots. Merging with the previous landmark studies was obtained by intersecting the covered positions and selecting Hg J carriers from the respective vcf files. We refrained from assembling a single tree for all subjects, as different filters applied in each study resulted in uncertain calls for some positions, some of which highly relevant for the Hg J topology. Genotyping of the core sample at 7 selected variable positions was by Sanger sequencing. The comparison of genotypes at the same 7 phylogeographically interesting markers was performed by scrutinizing each vcf file of the previous landmark studies separately.
Bam files have been deposited at the European Nucleotide Archive (https://www.ebi.ac.uk/ena) under study accession n. PRJEB25861 for 53 subjects, and sample accession n. ERS2065802-6 for subjects S206-S210, respectively.
References
Lazaridis, I. et al. Genomic insights into the origin of farming in the ancient Near East. Nature 536, 419–424 (2016).
Haak, W. et al. Massive migration from the steppe was a source for Indo-European languages in Europe. Nature 522, 207–211 (2015).
Hellenthal, G. et al. A genetic atlas of human admixture history. Science 343, 747–751 (2014).
Fu, Q. et al. The genetic history of Ice Age Europe. Nature 534, 200–205 (2016).
Kilinç, G. M. et al. The demographic development of the first farmers in Anatolia. Curr Biol 26, 2659–2666 (2016).
Gamba, C. et al. Genome flux and stasis in a five millennium transect of European prehistory. Nat Commun 5, 5257, https://doi.org/10.1038/ncomms6257 (2014).
Lazaridis, I. et al. Genetic origins of the Minoans and Mycenaeans. Nature 548, 214–218 (2017).
Lipson, M. et al. Parallel palaeogenomic transects reveal complex genetic history of early European farmers. Nature 551, 368 (2017).
Hofmanovà, Z. et al. Early farmers from across Europe directly descended from Neolithic Aegeans. Proc Natl Acad Sci USA 113, 6886–6891 (2016).
Paschou, P. et al. Maritime route of colonization of Europe. Proc Natl Acad Sci USA 111, 9211–9216 (2014).
Binder, D., Aspects de la néolithisation dans les aires padane, provençale et ligure in Néolithisation. British Archaeological Reports International Series 516, (ed. Aurenche, O. and Chauvin, J.)199–226 (Oxford, 1989).
Rowley-Conwy, P. Westward Ho! Curr Anthrop 52, S431–S451 (2011).
Horejs, B. et al. The Aegean in the early 7th millennium BC: Maritime networks and colonization. J World Prehist 28, 289–330 (2015).
Davison, K., Dolukhanov, P., Sarson, G. R. & Shukurov, A. The role of waterways in the spread of the Neolithic. J Archaeol Sci 33, 641–652 (2006).
Pinhasi, R., Thomas, M. G., Hofreiter, M., Currat, M. & Burger, J. The genetic history of Europeans. Trends Genet 28, 496–505 (2012).
Gonzàlez-Fortes, G. et al. Paleogenomic evidence for multi-generational mixing between Neolithic farmers and Mesolithic hunter-gatherers in the lower Danube basin. Cur Biol 27, 1–10 (2017).
Piazza, A., Cappello, N., Olivetti, E. & Rendine, S. A genetic history of Italy. Ann Hum Genet 52, 203–213 (1988).
Brisighelli, F. et al. Uniparental markers of contemporary Italian population reveals details on its pre-Roman heritage. PLoS ONE 7, e50794 (2012).
Tofanelli, S. et al. The Greeks in the West: genetic signatures of the Hellenic colonisation in southern Italy and Sicily. Eur J Hum Genet 24, 429–436 (2016).
Sarno, S. et al. Ancient and recent admixture layers in Sicily and Southern Italy trace multiple migration routes along the Mediterranean. Sci Rep 7, 1984 (2017).
Fiorito, G. et al. The Italian genome reflects the history of Europe and the Mediterranean basin. Eur J Hum Genet 24, 1056–1062 (2016).
Di Gaetano, C. et al. An overview of the genetic structure within the Italian population from genome-wide data. PLoS ONE 7, e43759 (2012).
Sarno, S. et al. An ancient Mediterranean melting pot: Investigating the uniparental genetic structure and population history of Sicily and Southern Italy. PLoS ONE 9, e96074 (2014).
Pala, M. et al. Mitochondrial haplogroup U5b3: a distant echo of the Epipaleolithic in Italy and the legacy of the early Sardinians. Am J Hum Genet 84, 814–821 (2009).
Batini, C. & Jobling, M. A. Detecting past male-mediated expansions using the Y chromosome. Hum Genet 136, 547–557 (2017).
Di Giacomo, F. et al. Y chromosomal haplogroup J as a signature of the post-neolithic colonization of Europe. Hum Genet 115, 357–371 (2004).
Semino, O. et al. Origin, diffusion, and differentiation of Y-chromosome haplogroups E and J: inferences on the neolithization of Europe and later migratory events in the Mediterranean area. Am J Hum Genet 74, 1023–1034 (2004).
Zalloua, P. A. et al. Identifying genetic traces of historical expansions: Phoenician footprints in the Mediterranean. Am J Hum Genet 83, 633–642 (2008).
Cinnioglu, C. et al. Excavating Y-chromosome haplotype strata in Anatolia. Hum Genet 114, 127–148 (2004).
Hallast, P. et al. The Y-chromosome tree bursts into leaf: 13,000 high confidence SNPs covering the majority of known clades. Mol Biol Evol 32, 661–673 (2015).
Karmin, M. et al. A recent bottleneck of Y chromosome diversity coincides with a global change in culture. Genome Res 25, 459–466 (2015).
Mallick, S. et al. The Simons Genome Diversity Project: 300 genomes from 142 diverse populations. Nature 538, 201–206 (2016).
Pagani, L. et al. Tracing the route of modern humans out of Africa by using 225 human genome sequences from Ethiopians and Egyptians. Am J Hum Genet 96, 986–991 (2015).
Poznik, G. et al. Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences. Nat Genet 48, 593–599 (2016).
Haber, M. et al. Continuity and admixture in the last five millennia of Levantine history from ancient Canaanite and present-day Lebanese genome sequences. Am J Hum Genet 101, 274–282 (2017).
Francalacci, P. et al. Low-pass DNA sequencing of 1200 Sardinians reconstructs European Y-chromosome phylogeny. Science 341, 565–569 (2013).
Harris, K. Evidence for recent, population-specific evolution of the human mutation rate. Proc Natl Acad Sci USA 112, 3439–44 (2015).
Singh, S. et al. Dissecting the influence of Neolithic demic diffusion on Indian Y-chromosome pool through J2-M172 haplogroup. Sci Rep 6, 19157 (2016).
Jobling, M. A. & Tyler-Smith, C. Human Y-chromosome variation in the genome-sequencing era. Nat Rev Genet 18, 485–497 (2017).
Balanovsky, O. Toward a consensus on SNP and STR mutation rates on the human Y-chromosome. Hum Genet 136, 575–590 (2017).
Henn, B. M., Cavalli-Sforza, L. L. & Feldman, M. W. The great human expansion. Proc Natl Acad Sci USA 109, 17758–17764 (2012).
Batini, C. et al. Large-scale recent expansion of European patrilineages shown by population resequencing. Nat Commun 6, 7152, https://doi.org/10.1038/ncomms8152 (2015).
King, R. J. et al. Differential Y-chromosome Anatolian influences on the Greek and Cretan Neolithic. Ann Hum Genet 72, 205–214 (2008).
Forster, P. et al. Continental and subcontinental distributions of mtDNA control region types. Int J Legal Med 116, 99–108 (2002).
Sengupta, S. et al. Polarity and temporality of high-resolution Y-chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic influence of central asian pastoralists. Am J Hum Genet 78, 202–221 (2006).
Al-Zahery, N. et al. In search of the genetic footprints of Sumerians: a survey of Y-chromosome and mtDNA variation in the Marsh Arabs of Iraq. BMC Evol Biol 11, 288 (2011).
Regueiro, M., Cadenas, A. M., Gayden, T., Underhill, P. A. & Herrera, R. J. Iran: Tricontinental nexus for Y-Chromosome driven migration. Hum Hered 61, 132–143 (2006).
Herrera, K. J. et al. Neolithic patrilineal signals indicate that the Armenian plateau was repopulated by agriculturalists. Eur J Hum Genet 20, 313–320 (2012).
Chiaroni, J. et al. Theemergence of Y-chromosome haplogroup J1e among Arabic-speaking populations. Eur J Hum Genet 18, 348–353 (2009).
Tofanelli, S. et al. J1-M267 Y lineage marks climate-driven pre-historical human displacements. Eur J Hum Genet 17, 1520–1524 (2009).
Keinan, A. & Clark, A. G. Recent explosive human population growth has resulted in an excess of rare genetic variants. Science 336, 740–743 (2012).
Markoe, G. Phoenicians. (British Museum Press, 2000).
Kristiansen, K. Europe before history. (Cambridge University Press, 1998).
Busby, G. B. et al. The role of recent admixture in forming the contemporary West Eurasian genomic landscape. Curr Biol 25, 2518–2526 and Erratum 2878 (2015).
King, R. J. et al. The coming of the Greeks to Provence and Corsica: Y-chromosome models of archaic Greek colonization of the western Mediterranean. BMC Evol Biol 11, 69 (2011).
Voskarides, K. et al. Y-chromosome phylogeographic analysis of the Greek-Cypriot population reveals elements consistent with Neolithic and Bronze Age settlements. Investig Genet 7, 1 (2016).
Messina, F. et al. Spatially explicit models to investigate geographic patterns in the distribution of forensic STRs: application to the North-Eastern Mediterranean. PLoS ONE 11, e0167065 (2016).
Lefèvre, F. Histoire du mond grec antique. (Librairie Générale Francaise, 2007).
Cruciani, F. et al. Tracing past human male movements in northern/eastern Africa and western Eurasia: new clues from Y-chromosomal haplogroups E-M78 and J-M12. Mol Biol Evol 24, 1300–1311 (2007).
Di Gaetano, C. et al. Differential Greek and northern African migrations to Sicily are supported by genetic evidence from the Y chromosome. Eur J Hum Genet 17, 91–99 (2008).
Kivisild, T. The study of human Y chromosome variation through ancient DNA. Hum Genet 136, 529–546 (2017).
Ségurel, L., Wyman, M. J. & Przeworski, M. Determinants of mutation rate variation in the human germline. Annu Rev Genomics Hum Genet 15, 47–70 (2014).
Tamura, K. et al. MEGA5: Molecular Evolutionary Genetics Analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28, 2731–2739 (2011).
Drummond, A. J. & Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol Biol 7, 214 (2007).
Jones, E. R. et al. Upper Palaeolithic genomes reveal deep roots of modern Eurasians. Nat Commun 6, 8912, https://doi.org/10.1038/ncomms9912 (2015).
Acknowledgements
We thank all anonymous donors for their voluntary participation in this study. We thank all collaborators who performed the interviews, filed them and collected the biological samples. We thank Luca Pagani, Mark Haber and Chris Tyler-Smith for providing data in vcf format. This work was supported by the Italian Ministry of Education, Progetti PRIN (grant numbers 2012JA4BTY_004 to FC and 2012JA4BTY_003 to AN) and by Sapienza University of Rome (grant numbers C26A153PCN and RM116154C9BE2C3F). FM was supported by a fellowship by the Italian Ministry of Education.
Author information
Authors and Affiliations
Contributions
A.N., F.C. and B.T. designed the experiment; A.N., N.A., A.L., E.I.M. and F.C. provided reagents (samples) A.F. and F.M. performed the DNA quality controls; A.F. analysed the NGS data and performed the SNP genotyping; A.N. and A.F. performed population analyses, statistical analysis and data interpretation; A.F., B.T., E.D.A., F.C. and A.N. discussed the results; A.F. and A.N. wrote the paper; All authors approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Finocchio, A., Trombetta, B., Messina, F. et al. A finely resolved phylogeny of Y chromosome Hg J illuminates the processes of Phoenician and Greek colonizations in the Mediterranean. Sci Rep 8, 7465 (2018). https://doi.org/10.1038/s41598-018-25912-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-25912-9
This article is cited by
-
Origin and diffusion of human Y chromosome haplogroup J1-M267
Scientific Reports (2021)
-
Phylogeographic review of Y chromosome haplogroups in Europe
International Journal of Legal Medicine (2021)
-
Phylogenetic history of patrilineages rare in northern and eastern Europe from large-scale re-sequencing of human Y-chromosomes
European Journal of Human Genetics (2021)
-
Middle eastern genetic legacy in the paternal and maternal gene pools of Chuetas
Scientific Reports (2020)
-
Early medieval genetic data from Ural region evaluated in the light of archaeological evidence of ancient Hungarians
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
