
Abstract
Clostridium botulinum is an anaerobic spore forming bacterium that produces the potent botulinum neurotoxin that causes a severe and fatal neuro-paralytic disease of humans and animals (botulism). C. botulinum Group II is a psychrotrophic saccharolytic bacterium that forms spores of moderate heat resistance and is a particular hazard in minimally heated chilled foods. Spore germination is a fundamental process that allows the spore to transition to a vegetative cell and typically involves a germinant receptor (GR) that responds to environmental signals. Analysis of C. botulinum Group II genomes shows they contain a single GR cluster (gerX3b), and an additional single gerA subunit (gerXAO). Spores of C. botulinum Group II strain Eklund 17B germinated in response to the addition of L-alanine, but did not germinate following the addition of exogenous Ca2+-DPA. Insertional inactivation experiments in this strain unexpectedly revealed that the orphan GR GerXAO is essential for L-alanine stimulated germination. GerX3bA and GerX3bC affected the germination rate but were unable to induce germination in the absence of GerXAO. No role could be identified for GerX3bB. This is the first study to identify the functional germination receptor of C. botulinum Group II.
Similar content being viewed by others
Introduction
Clostridium botulinum is the name given to a heterogeneous species that comprises four distinct groups of bacteria (C. botulinum Groups I to IV) that form the deadly botulinum neurotoxin1,2,3,4,5. The botulinum neurotoxin is the most potent toxin known, with a human lethal dose potentially being as little as 50 ng of neurotoxin2,5. There are a number of botulinum neurotoxin serotypes and subtypes6,7,8,9,10,11. Botulism in humans is most commonly associated with neurotoxin serotypes A, B or E, and more rarely with serotype F. There are three main types of human botulism. Foodborne botulism is a severe intoxication caused by consumption of food containing neurotoxin formed by C. botulinum during its growth in the food, whilst infant and wound botulism are both infections involving growth and neurotoxin formation in the body1,2,3,4,5. C. botulinum Groups I to IV are sufficiently distinct as to be considered separate species, and two of these Groups (C. botulinum Groups I and II) are associated with human botulism cases.
C. botulinum Group I is a mesophilic highly proteolytic bacterium that forms exceptionally heat resistant spores that are the target of the botulinum cook (121 °C/3 min) given to low acid canned foods1,2,3,4,5. Strains form up to three neurotoxins of serotype A, B and/or F. C. sporogenes is a close relative of C. botulinum Group I, and is often used as a surrogate in food safety tests2,5,12. However, recent genomic analysis indicates that C. sporogenes is not simply a non-toxigenic version of C. botulinum Group I. A majority (but not all) strains of C. botulinum Group I form botulinum neurotoxin, and while a fraction of C. sporogenes strains form serotype B neurotoxin, the majority of C. sporogenes strains do not form botulinum neurotoxin8,13,14,15,16. C. botulinum Group II is a psychrotrophic highly saccharolytic bacterium that forms spores of moderate heat resistance. Strains form a single neurotoxin of serotype B, E or F1,2,3,4,5. C. botulinum Group II is a concern for the continued safe production of minimally heated chilled foods17,18,19,20. The genomic diversity within C. botulinum Group II is greater than that within C. botulinum Group I/C. sporogenes, and C. botulinum Group II strains separate into two distantly related lineages. Lineage one contains the majority of serotype E strains, and these are strongly associated with the aquatic environment. Lineage two is dominated by serotype B strains, and also contains serotype F and occasional serotype E strains, and these strains are more frequently linked with terrestrial ecosystems. Interestingly, physiological differences have been reported between strains in the two lineages5,8,21,22,23,24,25,26,27,28,29,30. Spores formed by C. botulinum Groups I and II are widely distributed in the environment, and the prevention of human botulism relies on the identification and application of control measures that either prevent spores entering food or the body, or that prevent spore germination, cell multiplication and neurotoxin formation in food or the body. Spore germination is the key step in the transformation of the dormant resistant spore into multiplying vegetative cells that ultimately form botulinum neurotoxin. A greater understanding of spore germination can make an important contribution to microbiological food safety.
Spore germination has been widely studied in Bacillus species, and to a lesser extent in various species of Clostridium31,32,33,34. Spores germinate in response to nutrient germinants (e.g. amino acids) and to non-nutrient germinants (e.g. Ca2+: pyridine-2, 6-dicarboxylic acid (Ca2+-DPA)). Under favourable conditions spore dormancy is broken and germination ensues. In the environment, spore germination is generally initiated by a germinant receptor (GR) located in the spore inner membrane that responds to a nutrient germinant35,36. Following this initial step, the large depot of Ca2+-DPA within the spore is released, peptidoglycan in the spore cortex is hydrolysed, and core swelling and hydration is then followed by cell outgrowth. However, in C. difficile and C. perfringens spore cortex hydrolysis precedes Ca2+-DPA release. This general pattern of germination is followed in most species, with some notable differences for example in the germinants and how they are sensed, and how the cortex peptidoglycan is hydrolysed. Genes encoding four types of GR (gerX1-4) and 12 GR sub-types (that were designated by an additional letter) have been identified in C. botulinum Groups I to IV37. Genes encoding putative GR are well conserved within each C. botulinum Group, and are generally composed of three protein units GerA, GerB, and GerC. The position is similar in most other spore-forming bacteria, with germination requiring all three protein units to be present34. The genomic and physiological differences between the four C. botulinum Groups is also reflected in their germination apparatus and germination pathways37. Despite the importance of C. botulinum Groups I and II to both the food and pharmaceutical industry, our current understanding of nutrient induced germination and the role of the GR is limited, but is presently better understood in strains of C. botulinum Group I (and C. sporogenes) than in strains of C. botulinum Group II37,38,39.
Spores of Group I and Group II strains germinate in response to a variety of amino acids interacting with a GR37,39,40,41. Analysis of genome sequences has revealed the presence of three to five GR gene clusters in strains of C. botulinum Group I/C. sporogenes37,39. Detailed analysis revealed that amino acid induced germination in C. botulinum Group I strain ATCC 3502 required two GR (GerX1a and GerX1d) that were only functional when acting in synergy, while a function could not be identified for a third GR (GerX2b)39. In contrast, amino acid induced spore germination in C. sporogenes strain ATCC 15579 needed only a single GR (GerX1d), with two other GR contributing to the germination rate (GerX2c and GerX3a), and a fourth GR (GerX1a) having no discernible function39. The genetic complement of GR genes in C. botulinum Group II, however, is very different. Analysis of more than 150 C. botulinum Group II genome sequences has revealed the presence of only a single germinant receptor gene cluster (gerX3b) in each strain37,40. The gerX3b GR gene cluster has an unusual CA-B configuration that is different to the standard ABC configuration found in other C. botulinum Groups, as it has a bicistronic organization of gerAC and gerB genes orientated in opposite directions37,40. However, recently Clauwers et al. reported that this GR was not essential for amino acid induced spore germination in the C. botulinum Group II serotype E strain NCTC 1121940. Deletion of the gerX3b locus in this strain did not affect spore germination induced by a variety of amino acids, indicating that one or more unidentified GR are responsible for nutrient induced germination in strain NCTC 11219, and possibly also in other strains of C. botulinum Group II40. Strain NCTC 11219 belongs to C. botulinum Group II lineage one. The aim of the present study was to determine whether the GerX3b GR plays a major role in spore germination in strain Eklund 17B (that belongs to C. botulinum Group II lineage two) or whether the situation is similar to that reported by Clauwers et al.40 for strain NCTC 11219. We show that the GerX3b GR does not play a major role in spore germination in strain Eklund 17B, and have also identified a novel orphan germinant receptor, GerXAO, that is essential for amino acid induced germination.
Materials and Methods
Bacterial strains and growth conditions
C. botulinum Group II strain Eklund 17B (NRP) (serotype B) was grown anaerobically at 30 °C in tryptone-yeast-glucose medium (TYG). The Escherichia coli strain CA43442 was used for conjugal transfer and grown aerobically in Luria-Bertani medium (LB) at 37 °C. Where appropriate, growth media were supplemented with antibiotics at the following final concentrations; chloramphenicol 25 µg/ml, cycloserine 250 µg/ml, thiamphenicol 15 µg/ml, erythromycin 5 µg/ml and the chromogenic substrate 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-Gal) 80 µg/ml. All bacterial media supplements were purchased from Sigma (Gillingham, UK). All culture and incubation experiments were performed in an anaerobic cabinet (Don Whitley, Shipley, UK) using an atmosphere of CO2: H2: N2 (5: 10: 85, v/v).
Comparative genomics and phylogenetic analysis
For identification of gerX genes in the genomes of C. botulinum Group II strains, genome sequences were re-annotated using Prokka version 1.1243. Roary version 3.5.744 was used for comparative genomics. Alignments were made with Clustal omega45 and MEGA version 6.046 and used for generation of phylogenetic trees using the Neighbour Joining option. Figtree47 was used for annotation of phylogenetic trees. Conserved domains were identified using the Conserved Domain Database (NCBI)48.
Spore preparation
C. botulinum strain Eklund 17B was maintained as described previously41. Spores were produced on a biphasic cooked meat medium plate (CMP). Robertson’s cooked meat broth (CMB) (20 ml) (Southern Group Laboratories) was inoculated with 0.1 ml of culture (grown overnight in TYG broth at 30 °C) and incubated for 24 h at 30 °C. CMP were prepared using homogenised CMB (200 ml) with 3 g of agar (Sigma) and 0.2 g of glucose (Fisher Scientific, UK). CMP were subsequently inoculated with 3 ml of the CMB culture and incubated at 30 °C for 7 days. Spores were harvested, cleaned, separated from cell debris by discontinuous density gradient (Urografin 370, Schering, Germany) centrifugation and stored at 2 °C in water, as described previously41. Phase contrast microscopy confirmed that the suspensions consisted of >95% phase-bright spores.
Mutant generation
Genes encoding presumptive germination receptor (GR) sub-units (gerX3bA, gerX3bB, gerX3bC) in C. botulinum strain Eklund 17B were identified previously37, and gerXAO was identified in the present study. Mutants of C. botulinum strain Eklund 17B gerX3b subunits (gerX3bA; CB17B_RS15165, gerX3bB; CB17B_RS15170, gerX3bC; CB17B_RS15160, gerXAO; CB17B_RS12225) were generated using the Clostron system as previously described39. Briefly, target sites were identified using the Perutka method49 and mutants were generated as described50. Re-targeted introns were synthesised and ligated into the pMTL007C-E2 vector by DNA 2.0 (Menlo Park, USA). Retargeted intron plasmids were transformed into E. coli CA434. Confirmed (sequenced) plasmids were then transferred by conjugation into their respective clostridial host. For mutant complementation, plasmid pMTL83151 was used51. Primers bearing restriction sites compatible with pMTL83151 (BamHI and NheI) were used to amplify the gerX3b GR sub-units and its 5′ noncoding region, covering the predicted putative promoter. The resulting PCR product was digested with BamHI and NheI before being ligated into the pMTL83151 plasmid. Following confirmation by sequencing, complementation plasmids were transconjugated into their respective mutants using E. coli CA434 as described previously. Constructed mutants and plasmids utilised in this study are presented in Table S1. Primers used for verification of successful insertion events are listed in Table S2. PCR experiments were carried out using Phusion High-Fidelity PCR Master Mix with GC Buffer kit (Thermo Fisher). Plasmid isolation and PCR purification was performed using the Wizard Plus SV Minipreps DNA Purification System and the Wizard SV Gel and PCR Clean-Up System (Promega) respectively, as described in the provided Technical Manual. Chromosomal DNA isolation from suspected mutants were prepared as previously described52.
Spore germination
The germinant L-alanine (50 mM) (Sigma) was prepared in phosphate buffer (100 mM, pH 7.0) with NaHCO3 (50 mM) and L-lactate (50 mM). Germinant solutions were filter sterilised (0.45-µm syringe filter, Millipore, Bedford, MA, USA) and used within 1 h. Spore germination was also evaluated using Ca2+-DPA (60 mM, pH 7.4) in 20 mM Tris-HCl (pH 7.4) and NaHCO3 (50 mM). All spore suspensions were heat activated at 60 °C for 15 min, prior to the addition of potential germinants. Germination of spores at 30 °C was measured by a decrease in optical density (OD) at 600 nm every 5 min using a Bioscreen C analyser system (Labsystems, Basingstoke, UK). All germination experiments were performed in an anaerobic cabinet (Don Whitley) containing CO2: H2: N2 (5: 10: 85). Germination was expressed in terms of measured OD600 as a percentage of the initial OD600. To validate the OD600 measurements the proportion of germinated spores was visualised by the assessment of 200 spores in at least ten fields using phase-contrast microscopy. Typically, full germination was indicated when the OD600 fell to ~50–60% of its initial value. In some tests, a small fall in OD600 was observed (<10% of initial value). This was attributed to settling of spores in the Bioscreen wells, and was not accompanied by microscopic observation of spore germination. All experiments were performed in triplicate and the standard deviation calculated from three independent experiments. Statistical analysis of germination was completed using the two-tailed Student’s T-test with a significance level of 0.05.
Data Availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Results and Discussion
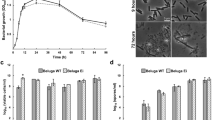
The effect of two potential germinants, L-alanine and exogenous Ca2+-DPA, on the germination of C. botulinum strain Eklund 17B spores was evaluated (Fig. 1). In the presence of L-lactate (50 mM) and NaHCO3 (50 mM) at 30 °C, the addition at 50 mM of L-alanine initiated spore germination, as observed by a > 50% drop of initial OD600, after approximately 30 min (Fig. 1). Direct spore counts by phase contrast microscopy revealed a ~50% drop in OD600 correlated to >99% spore germination. Similarly, amino acid induced spore germination of other C. botulinum Group II strains has been previously reported40,41. As previously described, L-lactate was an essential co-germinant for L-alanine germination but did not stimulate germination on its own41. Similarly, although experiments were performed under anaerobic conditions, this was considered non-essential41. The mode of action of L-lactate on spore germination is unknown, but may include one of the following; (1) L-alanine and L-lactate interact before interacting with the GR, (2) L-lactate may directly affect the GR together or sequentially with L-alanine or (3) L-lactate affects yet unidentified proteins that are part of the germination pathway. The GR-independent germinant Ca2+-DPA (60 mM) is a non-nutrient germinant of some Clostridium and Bacillus species34, including C. sporogenes and C. perfringens47,53,54. However, exogenous Ca2+-DPA failed to induce spore germination in C. botulinum Eklund 17B, as determined by optical density measurement and phase contrast microscopy (Fig. 1). This observation is consistent with that made in a recent study with C. botulinum Group II strain NCTC 11219 where exogenous Ca2+-DPA also failed to induce spore germination40. Spores of strains in both C. botulinum Group II lineage one (strain NCTC 11219) and in lineage two (strain Eklund 17B) are therefore not germinated by exogenous Ca2+-DPA. It would be interesting to test the effect of exogenous Ca2+-DPA on strains in all four C. botulinum Groups. Spores of Clostridium and Bacillus species both contain a large amount of a 1:1 chelate of Ca2+ and pyridine-2, 6-dicarboxylic acid (DPA) within their core. In Bacillus and C. sporogenes, exogenous Ca2+-DPA activates the cortex lytic enzyme CwlJ to trigger spore germination47,54, whereas in C. perfringens Ca2+-DPA triggers germination through the GerK GR53. The failure of exogenous Ca2+-DPA to induce C. botulinum Eklund 17B spore germination in the present study (and strain NCTC 11219 previously) could be due to the absence of CwlJ, as there are no genes encoding CwlJ homologs in C. botulinum Group II genomes37. Moreover, C. botulinum Group II genomes do not contain the cortex lytic enzyme SleB found in most Bacillus and clostridia34 but instead possess the cortex lytic enzyme SleC as a zymogen. As with C. botulinum Eklund 17B, C. difficile spores do not germinate with the addition of Ca2+-DPA55, and their genomes lack genes encoding a homolog of CwlJ and also a ‘classical’ nutrient GR55,56 although they do possess a SleC similarly to C. botulinum Group II strains.

Effect of L-alanine (50 mM) and exogenous Ca2+-DPA (60 mM) on germination of C. botulinum Eklund 17B wild type spores. Data labels (right) refer to percentage germination observed by phase contrast microscopy at 3 hours. Error bars represent the standard deviation of 3 independent experiments.
In most species, spore germination is initiated by a germinant receptor (GR) located in the spore inner membrane. The C. botulinum Eklund 17B genome includes a gerX3b (gerX3bA, gerX3bB, gerX3bC) receptor (Fig. 2a,b), similar to those described in other C. botulinum Group II strains37,40. Based on the sequence of the entire germinant receptor cluster, strains of C. botulinum Group II can be separated into two clades (Fig. 2a) that directly align with the two lineages derived from whole genome analysis8,28. The gerX3b germinant receptor in strain Eklund 17B is located in clade two in Fig. 2a (while the gerX3b germinant receptor in strain NCTC 11219 studied by Clauwers40 is found in clade one). Clade one exclusively contains strains forming type E neurotoxin, while clade two comprises strains forming type B, E or F neurotoxin (Fig. 2a). The gerX3b receptor gene cluster has a bicistronic organization, with gerAC and gerB genes being orientated in the opposite direction (CA-B configuration) (Fig. 2b). Analysis of more than 150 C. botulinum Group II genome sequences also revealed the presence of a single gerA subunit (termed here gerXAO, Fig. 2c) in each strain, indicating its likely importance. This single gerA subunit was distantly located on the chromosome from the gerX3b receptor gene cluster. The gerXAO gene, in general, is flanked by an asparagine synthetase (asnB) and an uncharacterized S1 RNA binding domain (yitL) gene. The annotation of this single gerA subunit is somewhat inconsistent with annotations frequently describing it as encoding a germination protein (GerA spore germination protein), or more rarely a sporulation protein (Stage V sporulation protein AF (SpoVAF)). However, Conserved Domain analysis confirmed that the GerXAO protein belongs to the GerA superfamily (cl10605). BlastP analysis also revealed high similarity between orphan GerA gene products from C. botulinum Group II strains (90–100%) and those identified by sequence similarity as spore germination proteins from C. perfringens and C. taenosporium (61% and 90% respectively). These are also not associated with either an ABC- or CA-B-type cluster structure. Interestingly, peptide sequence alignment of the GerXAO gene product with the receptor-associated (CA-B) GerA revealed only ~27% similarity at the amino acid level.

Relatedness of GerX3b receptor gene clusters in Clostridium botulinum Group II and genetic organization of the gerX subtypes. (a) Relatedness of gerX3b types in C. botulinum Group II (toxin types shown in coloured boxes). gerX associated proteins used for comparisons; gerX3bA (Locus tag; CB17B_RS15165), gerX3bB (Locus tag; CB17B_RS15170), gerX3bC (Locus tag; CB17B_RS15160) and gerXAO (Locus tag; CB17B_RS12225). *Represents strain C. botulinum Eklund 17B (NRP) (Accession number; FR745875) used in this study. †Represents strain C. botulinum NCTC 11219 for comparison. Clade I consist of strains forming neurotoxin type E, and Clade II includes strains forming type neurotoxin types B, E or F. Strains used for comparison of the gerX3b types see Brunt et al.37. The phylogenetic tree is based on single nucleotide polymorphisms (SNPs). Values shown at branches represent bootstrap values provided by ParSNP. (b) Genetic organization of the gerX3b gene cluster. (c) gerXAO is flanked by asparagine synthetase (asnB) and an uncharacterized S1 RNA binding domain protein (yitL).
To characterise the functionality of the putative GR encoded by gerX3b (gerX3bA, gerX3bB, gerX3bC) in C. botulinum Eklund 17B, single insertional mutants of each gene were generated and confirmed by PCR and sequencing (Fig. 3). However, it should be noted that given the absence of formal characterisation of any C. botulinum gene promoters, mutation of gerX3bA may have a polar effect on gerX3bC. Mutation of C. botulinum Group II is challenging40, and several attempts were needed to isolate mutants for this strain. Spores generated from these mutants were analysed for L-alanine induced germination (Fig. 4). Mutants gerX3bA− and gerX3bC− failed to germinate (<10% fall in OD600) with L-alanine after 1 h of exposure (Fig. 4a). However, gerX3bC− spores started to germinate slowly after 1 h (as determined by a fall in OD600). The OD600 of wild type spores decreased (∼50%) after ~30 min, representing efficient and complete spore germination (≥99%) as observed by phase contrast microscopy. Interestingly, the gerX3bB− mutant exhibited comparable germination profiles to those of wild type spores indicating a lack of function in this stage of the germination process. This is in contrast to GerKB in C. perfringens, which is involved in spore viability and outgrowth57. The lack of phenotype for the Eklund 17B gerX3bB− mutant is also surprising as the GerB protein in Bacillus is proposed to be responsible for germinant binding and influencing the GerC protein58,59,60. This apparent lack of function is additionally unexpected when the abundance of monocistronic gerB genes is considered, especially in the knowledge that C. botulinum Group II genomes are relatively stable with the potential for horizontal gene transfer considered low8,28. Further work is required, including examination of the orphan monocistronic gerB genes in C. botulinum Group I.

Confirmation of insertional mutagenesis. (a) PCR screens of C. botulinum gerX gene mutants confirming that the intron has successfully inserted into the target gene. Numbers represent primer sets used. Gel image has been cropped, full image available in supplementary figure (S1). (b) C. botulinum germination genes showing Clostron insertion sites (red arrows). Black arrows represent primer annealing sites. Primer set 1 (gerC-F/R) anneals to the target gene CB17B_RS15160 gerX3bC either side of the intron insertion site and confirms the intron is present in the target gene (expected band size of mutants ~2 kb). Primer set 2 (gerAB-F/R) anneals to the target gene CB17B_RS15165 gerX3bA and CB17B_RS15170 gerX3bB either side of the intron insertion sites and confirms the intron is present in the target gene. Primer set 3 (gerAO-F3/R3) anneals to the target gene CB17B_RS12225 gerXOA either side of the intron insertion sites and confirms the intron is present in the target gene.

Effect of L-alanine (50 mM) on spore Germination of C. botulinum Eklund 17B gerX3bA−, gerX3bB−, gerX3bC−, gerXAO− and wild type. (a) Kinetics of spore germination determined using fall in OD600. (b) Spore germination after 3 hours (percentage of phase dark spores as determined by phase contrast microscopy). (c) Spore germination after 24 hours (percentage of phase dark spores as determined by phase contrast microscopy). (d) Kinetics of spore germination of the complemented mutants determined using fall in OD600. Error bars represent the standard deviation of three independent experiments. *Student’s T-test with a significance level of 0.05. **Not significant at level of 0.05 compared to the wild type. +complemented strains.
Phase contrast microscopy was used to determine the percentage of spores that had germinated (i.e. become phase dark) after 3 h and 24 h at 30 °C (Fig. 4b,c). Incubation with L-alanine for 3 h, resulted in negligible germination of mutant gerX3bA− and limited germination of mutant gerX3bC−, but complete germination of the wild type and gerX3bB− (Fig. 4b). Following incubation with L-alanine for 24 h, mutants gerX3bA− and gerX3bC− displayed germination approaching that of the wild type and gerX3bB− (Fig. 4c). Complementation of gerX3bA− and gerX3bB− using plasmid pMTL83151 containing the gerX3b GR sub-units and their 5′ noncoding region restored the germination phenotype of gerX3bA− and did not alter the wild type phenotype of gerX3bB−. Albeit, the rates of germination of these complemented mutants were not fully restored compared to that of the wild type (Fig. 4d). Complementation of gerX3bC− did not restore the germination rate phenotype; this lack of successful complementation has been observed for many generated mutants in clostridia, but the reason is unclear37,39,61,62. However, in the absence of formal characterisation of any C. botulinum gene promoters, apart from BotR regulated ones in the neurotoxin gene cluster, we predicted that the promoter to be located in the intergenic region upstream of the gerX3bA gene and therefore the complementation of gerX3bC− may have failed due to the lack of a native promoter. Furthermore, attempts at expressing the whole receptor failed so complementation of gerX3bC− strain could not be assessed. The configuration of the bicistronic divergent gerCA and gerB gene cluster found in all C. botulinum Group II strains is comparable to the gerK locus reported in C. perfringens which comprises of a bicistronic gerKA-gerKC and monocistronic gerKB in the opposite orientation53,63. The similarity of the GR amino acid sequences for C. perfringens SM101 is approximately GerKA 63%, GerKB 36% and GerKC 46% when compared to those of C. botulinum Eklund 17B GerX3bA, GerX3bB, and GerX3bC, respectively. In C. perfringens, the GerKC protein is considered to be the main GR with GerAA and GerKB proteins playing a minor role in spore germination53,63. However, a recent report has now demonstrated that in the C. botulinum Group II strain NCTC 11219 GerX3b was not required for nutrient induced germination40. In this work, the entire GR cluster was deleted, and germination was similar in the wild type and mutant after 4 h/30 °C, as measured by number of heat resistant spores and DPA loss. Phase contrast microscopy additionally revealed that the rate of formation of phase dark spores was similar and virtually complete after 1 h/30 °C40. The findings made in the present study with strain Eklund 17B are consistent with the observation made with strain NCTC 11219 that the GerX3b receptor is not essential for spore germination in strains of C. botulinum Group II, and importantly now extend the finding to both major C. botulinum Group II lineages. We also demonstrate, for the first time, that in strain Eklund 17B two of the GR receptor proteins contribute to the germination rate and in their absence the germination rate is much slower with L-alanine compared to the wild type. The presence of gerX3b in the genome of all C. botulinum Group II strains examined to date37,40 indicates that the product of this gene cluster is likely to have an important biological role, for example it may respond to an as yet unknown nutrient/non-nutrient germinants, or perform a structural role for instance in the organisation of a germinosome64.
Given the findings presented here and those made by Clauwers et al.40 that the GerX3b receptor does not play a major role in spore germination in C. botulinum Group II, functional GR were now sought, and the potential role of GerXAO was assessed. Following insertional mutagenesis, spores were produced, and tested for L-alanine induced germination. Mutant gerXAO− failed to germinate (<10% fall in OD600) with more than 99% of spores appearing phase bright after incubation for 24 h (Fig. 4). Additionally, complementation of gerXAO− using plasmid pMTL83151 restored germination to similar levels of the wild type (≥99%). Thus, GerXAO plays a major role in spore germination in Eklund 17B.
An orphan gerA gene is located in the genome of several species of Clostridium32,65, and is present in the genome of all of more than 150 strains of C. botulinum Group II examined in the present study, which indicates there is an evolutionary advantage to its retention as a functional gene. However, the orphan GerA present in C. perfringens played only a minor role in germination53. Tentative evidence in this study also revealed that sporulation efficiency appeared to be affected by mutation of gerXAO, as determined by phase contrast microscopy. Again, the reasons for this are unclear, we can only speculate that the loss of this receptor protein has an effect on an as yet unknown protein or signal that is involved in sporulation. Although this is the first report of an essential requirement for a monocistronic GerA in spore germination, function has been attributed to a monocistronic GerB which is involved in germinant specificity and rate in spores of Bacillus megaterium QM B155166. Moreover, this GerB receptor subunit interacts with GerUA and GerUC to form a functional receptor. In C. botulinum Group II strain Eklund 17B, the rate of spore germination is more rapid when GerXAO interacts with GerX3bA and/or GerX3bC, than when either GerX3bA or GerX3bC are absent. However, further work is certainly warranted to understand the exact mechanism of the GerXAO receptor and to whether it responds to further germinants. Interestingly, the position in C. botulinum Group II strain Eklund 17B, where GerXAO brings about germination on its own with other proteins (GerX3bA and/or GerX3bC) increasing the rate, contrasts with the situation in most other spore-forming bacteria where germination involves a GR with three protein units (GerA, GerB, and GerC). A potential role for presently unidentified proteins that interact with GerXAO to bring about germination cannot also be excluded. Following the results presented here we now propose an updated version37 of the germination model for C. botulinum Group II strains (Fig. 5). Firstly, the nutrient germinant binds to the GerXAO receptor, followed by Ca2+-DPA release through the SpoVA channel (Stage I). In stage II, the cortex lytic enzyme SleC is activated by CspB, followed by cortex hydrolysis, membrane and coat degradation, the resumption of metabolism and ultimately cell outgrowth. Although, the order of these events still needs to be resolved in C. botulinum group II strains (Fig. 5)

Comparison of the models proposed for the germination pathways of C. botulinum Group I and Group II. (a) Group II, with GerXAO (shaded grey) as the major germinant receptor and GerX3bA & GerX3bC (shaded green) involved in germination rate. Initially, L-alanine binds to the GerXAO receptor, followed by Ca2+-DPA release through the SpoVA channel (Stage I). In stage II, the cortex lytic enzyme SleC is activated by CspB, followed by cortex hydrolysis, membrane, and coat degradation, the resumption of metabolism and ultimately cell outgrowth. *Cortex lytic enzymes include SleC2 a/b and SleB2. (b) Germination pathway for C. botulinum strain ATCC3502 (Group I)37,39. This system involves the recognition of nutrient germinants by their cognate receptor GerX1a &GerX1d acting together, followed by Ca2+-DPA release through the proposed SpoVA channel (Stage I). In stage II, the CLEs CwlJ and SleB (†) are activated, followed by cortex hydrolysis, membrane and coat degradation, the recommencement of metabolism and eventually cell outgrowth. “?” indicates that the timing of the release of Ca2+-DPA is presently unknown in C. botulinum Group I and Group II and may be released following germinant binding or after cortex hydrolysis.
In summary, this is the first report that successfully identifies the key component of the germination machinery of C. botulinum Group II strain Eklund 17B. The presence of the orphan gene encoding GerXAO in all C. botulinum Group II genomes so far examined strongly suggests that the germination model proposed in this work will stand for all Group II strains. The role of GerX3bB, a gene equally conserved but with no obvious function, remains un-resolved. However, it is clear that gene products of its neighbouring operon interact with that of gerXAO to increase the efficiency of the germination process.
References
Bruggemann, H., Wollherr, A., Mazuet, C. & Popoff, M. In Genomes of Foodborne and Waterborne Pathogens Vol. ISBN-13: 978-1555814571 (eds Fratamico P., Liu Y., & Kathariou S.) Ch. 13, 185–212 (ASM Press, 2011).
Hauschild, A. H. W. Clostridium botulinum. In: Doyle M.P., editor. Foodborne bacterial pathogens. Marcel Dekker; New York (1989).
Johnson, E. A. Clostridium botulinum. In Doyle M. P. & Buchanan R. L. (Eds), Food Microbiology: Fundamentals and Frontiers (4th ed., pp. 441–463). Washington, DC: ASM Press (2013).
Lindström, M., Fredriksson-Ahomaa, M. & Korkeala H. In Clostridia, molecular biology in the post-genomic era(ed Brüggemann H., Gottschalk, G.)103–130 (2009).
Peck, M. W. Biology and genomic analysis of Clostridium botulinum. Adv Microb Physiol 55, 183–265, 320, https://doi.org/10.1016/S0065-2911(09)05503-9 (2009).
Hill, K. K., Xie, G., Foley, B. T. & Smith, T. J. Genetic diversity within the botulinum neurotoxin-producing bacteria and their neurotoxins. Toxicon 107, 2–8, https://doi.org/10.1016/j.toxicon.2015.09.011 (2015).
Peck, M. W. et al. Historical perspectives and guidelines for Botulinum neurotoxin subtype nomenclature. Toxins (Basel) 9, 38, https://doi.org/10.3390/toxins9010038 (2017).
Peck, M. W. & van Vliet, A. H. Impact of Clostridium botulinum genomic diversity on food safety. Curr Opin. Food Sci 10, 52–59, https://doi.org/10.1016/j.cofs.2016.09.006 (2016).
Brunt, J., Carter, A. T., Stringer, S. C. & Peck, M. W. Identification of a novel botulinum neurotoxin gene cluster in Enterococcus. FEBS Lett 592, 310–317, https://doi.org/10.1002/1873-3468.12969 (2018).
Zhang, S. et al. Identification of a Botulinum neurotoxin-like toxin in a commensal strain of Enterococcus faecium. Cell Host & Microbe 23, 169–176 e166, https://doi.org/10.1016/j.chom.2017.12.018 (2018).
Zhang, S. et al. Identification and characterization of a novel botulinum neurotoxin. Nat Commun 8, 14130, https://doi.org/10.1038/ncomms14130 (2017).
Brunt, J., Cross, K. L. & Peck, M. W. Apertures in the Clostridium sporogenes spore coat and exosporium align to facilitate emergence of the vegetative cell. Food Microbiol 51, 45–50, https://doi.org/10.1016/j.fm.2015.04.013 (2015).
Carter, A. T. et al. Independent evolution of neurotoxin and flagellar genetic loci in proteolytic Clostridium botulinum. BMC Genomics 10, 115, https://doi.org/10.1186/1471-2164-10-115 (2009).
Weigand, M. R. et al. Implications of genome-based discrimination between Clostridium botulinum Group I and Clostridium sporogenes strains for bacterial taxonomy. Appl Environ Microbiol 81, 5420–5429, https://doi.org/10.1128/AEM.01159-15 (2015).
Williamson, C. H. et al. Comparative genomic analyses reveal broad diversity in botulinum-toxin-producing Clostridia. BMC Genomics 17, 180, https://doi.org/10.1186/s12864-016-2502-z (2016).
Williamson, C. H. D. et al. Differentiating Botulinum neurotoxin-producing Clostridia with a simple, multiplex PCR assay. Appl Environ Microbiol, https://doi.org/10.1128/AEM.00806-17 (2017).
Peck, M. W. Clostridium botulinum and the safety of minimally heated, chilled foods: an emerging issue? J Appl Microbiol 101, 556–570, https://doi.org/10.1111/j.1365-2672.2006.02987.x (2006).
Peck, M. W., Goodburn, K. E., Betts, R. P. & Stringer, S. C. Assessment of the potential for growth and neurotoxin formation by non-proteolytic Clostridium botulinum in short shelf-life commercial foods designed to be stored chilled. Trends Food Sci. Technol. 19, 207–216, https://doi.org/10.1016/j.tifs.2007.12.006 (2008).
Peck, M. W. & Stringer, S. C. The safety of pasteurised in-pack chilled meat products with respect to the foodborne botulism hazard. Meat Sci 70, 461–475, https://doi.org/10.1016/j.meatsci.2004.07.019 (2005).
Peck, M. W., Stringer, S. C. & Carter, A. T. Clostridium botulinum in the post-genomic era. Food Microbiol 28, 183–191, https://doi.org/10.1016/j.fm.2010.03.005 (2011).
Carter, A. T. & Peck, M. W. Genomes, neurotoxins and biology of Clostridium botulinum Group I and Group II. Res Microbiol 166, 303–317, https://doi.org/10.1016/j.resmic.2014.10.010 (2015).
Hielm, S., Hyytia, E., Andersin, A. B. & Korkeala, H. A high prevalence of Clostridium botulinum type E in Finnish freshwater and Baltic Sea sediment samples. J Appl Microbiol 84, 133–137, https://doi.org/10.1046/j.1365-2672.1997.00331.x (1998).
Hill, K. K. et al. Genetic diversity among Botulinum neurotoxin-producing clostridial strains. J Bacteriol 189, 818–832, https://doi.org/10.1128/JB.01180-06 (2007).
Leclair, D. et al. Distribution of Clostridium botulinum type E strains in Nunavik, Northern Quebec, Canada. Appl Environ Microbiol 79, 646–654, https://doi.org/10.1128/AEM.05999-11 (2013).
Mazuet, C. et al. Diversity of Group I and II Clostridium botulinum strains from France including recently identified subtypes. Genome biology and evolution 8, 1643–1660, https://doi.org/10.1093/gbe/evw101 (2016).
Mazuet, C. et al. An atypical outbreak of food-borne botulism due to Clostridium botulinum types B and E from ham. J Clin Microbiol 53, 722–726, https://doi.org/10.1128/JCM.02942-14 (2015).
Raphael, B. H. et al. Analysis of a unique Clostridium botulinum strain from the Southern hemisphere producing a novel type E botulinum neurotoxin subtype. BMC Microbiol 12, 245, https://doi.org/10.1186/1471-2180-12-245 (2012).
Stringer, S. C. et al. Genomic and physiological variability within Group II (non-proteolytic) Clostridium botulinum. BMC Genomics 14, 333, https://doi.org/10.1186/1471-2164-14-333 (2013).
Wachnicka, E., Stringer, S. C., Barker, G. C. & Peck, M. W. Systematic assessment of Nonproteolytic Clostridium botulinum spores for heat resistance. Appl Environ Microbiol 82, 6019–6029, https://doi.org/10.1128/AEM.01737-16 (2016).
Weedmark, K. A. et al. Clostridium botulinum Group II isolate phylogenomic profiling using whole-genome sequence data. Appl Environ Microbiol 81, 5938–5948, https://doi.org/10.1128/AEM.01155-15 (2015).
Bhattacharjee, D., McAllister, K. N. & Sorg, J. A. Germinants and their receptors in Clostridia. J Bacteriol 198, 2767–2775, https://doi.org/10.1128/JB.00405-16 (2016).
Paredes-Sabja, D., Setlow, P. & Sarker, M. R. Germination of spores of Bacillales and Clostridiales species: mechanisms and proteins involved. Trends Microbiol 19, 85–94, https://doi.org/10.1016/j.tim.2010.10.004 (2011).
Setlow, P. Germination of spores of Bacillus species: what we know and do not know. J Bacteriol 196, 1297–1305, https://doi.org/10.1128/JB.01455-13 (2014).
Setlow, P., Wang, S. & Li, Y. Q. Germination of spores of the orders Bacillales and Clostridiales. Annu Rev Microbiol 71, 459–477, https://doi.org/10.1146/annurev-micro-090816-093558 (2017).
Paidhungat, M. & Setlow, P. Localization of a germinant receptor protein (GerBA) to the inner membrane of Bacillus subtilis spores. J Bacteriol 183, 3982–3990, https://doi.org/10.1128/JB.183.13.3982-3990.2001 (2001).
Hudson, K. D. et al. Localization of GerAA and GerAC germination proteins in the Bacillus subtilis spore. J Bacteriol 183, 4317–4322, https://doi.org/10.1128/JB.183.14.4317-4322.2001 (2001).
Brunt, J., van Vliet, A. H., van den Bos, F., Carter, A. T. & Peck, M. W. Diversity of the germination apparatus in Clostridium botulinum Groups I, II, III, and IV. Front Microbiol 7, 1702, https://doi.org/10.3389/fmicb.2016.01702 (2016).
Broussolle, V. et al. Molecular and physiological characterisation of spore germination in Clostridium botulinum and C. sporogenes. Anaerobe 8, 89–100, https://doi.org/10.1006/anae.2002.0424 (2002).
Brunt, J. et al. Functional characterisation of germinant receptors in Clostridium botulinum and Clostridium sporogenes presents novel insights into spore germination systems. PLoS Pathog 10, e1004382, https://doi.org/10.1371/journal.ppat.1004382 (2014).
Clauwers, C., Lood, C., van Noort, V. & Michiels, C. W. Canonical germinant receptor is dispensable for spore germination in Clostridium botulinum group II strain NCTC 11219. Sci Rep 7, 15426, https://doi.org/10.1038/s41598-017-15839-y (2017).
Plowman, J. & Peck, M. W. Use of a novel method to characterize the response of spores of non-proteolytic Clostridium botulinum types B, E and F to a wide range of germinants and conditions. J. Appl. Microbiol. 92, 681–694, https://doi.org/10.1046/j.1365-2672.2002.01569.x (2002).
Purdy, D. et al. Conjugative transfer of clostridial shuttle vectors from Escherichia coli to Clostridium difficile through circumvention of the restriction barrier. Mol. Microbiol. 46, 439–452, https://doi.org/10.1046/j.1365-2958.2002.03134.x (2002).
Seemann, T. Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069, https://doi.org/10.1093/bioinformatics/btu153 (2014).
Page, A. J. et al. Roary: rapid large-scale prokaryote pan genome analysis. Bioinformatics 31, 3691–3693, https://doi.org/10.1093/bioinformatics/btv421 (2015).
Sievers, F. et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol Syst Biol 7, 539, https://doi.org/10.1038/msb.2011.75 (2011).
Tamura, K. et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28, 2731–2739, https://doi.org/10.1093/molbev/msr121 (2011).
Wang, S., Brunt, J., Peck, M. W., Setlow, P. & Li, Y. Q. Analysis of the germination of individual Clostridium sporogenes spores with and without germinant receptors and cortex-lytic enzymes. Front Microbiol 8, 2047, https://doi.org/10.3389/fmicb.2017.02047 (2017).
Marchler-Bauer, A. et al. CDD/SPARCLE: functional classification of proteins via subfamily domain architectures. Nucleic Acids Res 45, D200–D203, https://doi.org/10.1093/nar/gkw1129 (2017).
Perutka, J., Wang, W. J., Goerlitz, D. & Lambowitz, A. M. Use of computer-designed group II introns to disrupt Escherichia coli DExH/D-box protein and DNA helicase genes. J. Mol. Biol. 336, 421–439, https://doi.org/10.1016/j.jmb.2003.12.009 (2004).
Heap, J. T. et al. The ClosTron: Mutagenesis in Clostridium refined and streamlined. J Microbiol Methods 80, 49–55, https://doi.org/10.1016/j.mimet.2009.10.018 (2010).
Heap, J. T., Pennington, O. J., Cartman, S. T. & Minton, N. P. A modular system for Clostridium shuttle plasmids. J Microbiol Methods 78, 79–85, https://doi.org/10.1016/j.mimet.2009.05.004 (2009).
Sebaihia, M. et al. Genome sequence of a proteolytic (Group I) Clostridium botulinum strain Hall A and comparative analysis of the clostridial genomes. Genome Res 17, 1082–1092, https://doi.org/10.1101/gr.6282807 (2007).
Paredes-Sabja, D., Torres, J. A., Setlow, P. & Sarker, M. R. Clostridium perfringens spore germination: characterization of germinants and their receptors. J Bacteriol 190, 1190–1201, https://doi.org/10.1128/JB.01748-07 (2008).
Paidhungat, M., Ragkousi, K. & Setlow, P. Genetic requirements for induction of germination of spores of Bacillus subtilis by Ca(2+)-dipicolinate. J Bacteriol 183, 4886–4893, https://doi.org/10.1128/JB.183.16.4886-4893.2001 (2001).
Wang, S., Shen, A., Setlow, P. & Li, Y. Q. Characterization of the dynamic germination of individual Clostridium difficile spores using Raman spectroscopy and differential interference contrast microscopy. J Bacteriol 197, 2361–2373, https://doi.org/10.1128/JB.00200-15 (2015).
Francis, M. B., Allen, C. A., Shrestha, R. & Sorg, J. A. Bile acid recognition by the Clostridium difficile germinant receptor, CspC, is important for establishing infection. PLoS Pathog 9, e1003356, https://doi.org/10.1371/journal.ppat.1003356 (2013).
Paredes-Sabja, D., Setlow, P. & Sarker, M. R. Role of GerKB in germination and outgrowth of Clostridium perfringens spores. Appl Environ Microbiol 75, 3813–3817, https://doi.org/10.1128/AEM.00048-09 (2009).
Christie, G., Gotzke, H. & Lowe, C. R. Identification of a receptor subunit and putative ligand-binding residues involved in the Bacillus megaterium QM B1551 spore germination response to glucose. J Bacteriol 192, 4317–4326, https://doi.org/10.1128/JB.00335-10 (2010).
Christie, G., Lazarevska, M. & Lowe, C. R. Functional consequences of amino acid substitutions to GerVB, a component of the Bacillus megaterium spore germinant receptor. J Bacteriol 190, 2014–2022, https://doi.org/10.1128/JB.01687-07 (2008).
Cooper, G. R. & Moir, A. Amino acid residues in the GerAB protein important in the function and assembly of the alanine spore germination receptor of Bacillus subtilis 168. J Bacteriol 193, 2261–2267, https://doi.org/10.1128/JB.01397-10 (2011).
Li, J., Chen, J., Vidal, J. E. & McClane, B. A. The Agr-like quorum-sensing system regulates sporulation and production of enterotoxin and beta2 toxin by Clostridium perfringens type A non-food-borne human gastrointestinal disease strain F5603. Infect Immun 79, 2451-2459. https://doi.org/10.1128/IAI.00169-11 (2011).
Meaney, C. A., Cartman, S. T., McClure, P. J. & Minton, N. P. Optimal spore germination in Clostridium botulinum ATCC 3502 requires the presence of functional copies of SleB and YpeB, but not Cwl. J. Anaerobe 34, 86–93, https://doi.org/10.1016/j.anaerobe.2015.04.015 (2015).
Banawas, S. et al. The Clostridium perfringens germinant receptor protein GerKC is located in the spore inner membrane and is crucial for spore germination. J Bacteriol 195, 5084–5091, https://doi.org/10.1128/JB.00901-13 (2013).
Griffiths, K. K., Zhang, J., Cowan, A. E., Yu, J. & Setlow, P. Germination proteins in the inner membrane of dormant Bacillus subtilis spores colocalize in a discrete cluster. Mol Microbiol 81, 1061–1077, https://doi.org/10.1111/j.1365-2958.2011.07753.x (2011).
Xiao, Y., Francke, C., Abee, T. & Wells-Bennik, M. H. Clostridial spore germination versus bacilli: genome mining and current insights. Food Microbiol 28, 266–274, https://doi.org/10.1016/j.fm.2010.03.016 (2011).
Christie, G. & Lowe, C. R. Role of chromosomal and plasmid-borne receptor homologues in the response of Bacillus megaterium QM B1551 spores to germinants. J Bacteriol 189, 4375–4383, https://doi.org/10.1128/JB.00110-07 (2007).
Acknowledgements
The authors are grateful to Dr. John Walshaw for his contribution to the genomic analysis. The authors acknowledge support from the Biotechnology and Biological Sciences Research Council as part of the BBSRC Institute Strategic Programme on Gut Health and Food Safety BB/J004529/1.
Author information
Authors and Affiliations
Contributions
J.B., A.T.C. and M.W.P. conceived the study. J.B., A.T.C. and H.V.P. performed the experiments. J.B., A.T.C., H.V.P. and M.W.P. analysed the data. J.B., A.T.C., and M.W.P. wrote the paper. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Brunt, J., Carter, A.T., Pye, H.V. et al. The orphan germinant receptor protein GerXAO (but not GerX3b) is essential for L-alanine induced germination in Clostridium botulinum Group II. Sci Rep 8, 7060 (2018). https://doi.org/10.1038/s41598-018-25411-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-25411-x
This article is cited by
-
Phage lysin that specifically eliminates Clostridium botulinum Group I cells
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
