
Abstract
Staphylococcus aureus can develop a small colony variant (SCV) phenotype in response to sub-lethal exposure to the biocide triclosan. In the current study, whole genome sequencing was performed and changes in virulence were investigated in five Staphylococcus aureus strains following repeated exposure to triclosan. Following exposure, 4/5 formed SCV and exhibited point mutations in the triclosan target gene fabI with 2/4 SCVs showing mutations in both fabI and fabD. The SCV phenotype was in all cases immediately reversed by nutritional supplementation with fatty acids or by repeated growth in the absence of triclosan, although fabI mutations persisted in 3/4 reverted SCVs. Virulence, determined using keratinocyte invasion and Galleria mellonella pathogenicity assays was significantly (p < 0.05) attenuated in 3/4 SCVs and in the non-SCV triclosan-adapted bacterium. Proteomic analysis revealed elevated FabI in 2/3 SCV and down-regulation in a protein associated with virulence in 1/3 SCV. In summary, attenuated keratinocyte invasion and larval virulence in triclosan-induced SCVs was associated with decreases in growth rate and virulence factor expression. Mutation occurred in fabI, which encodes the main triclosan target in all SCVs and the phenotype was reversed by fatty acid supplementation, demonstrating an association between fatty acid metabolism and triclosan-induced SCV.
Similar content being viewed by others


introduction
Staphylococcus aureus causes a wide range of community and hospital acquired infections1. Coupled with the colonisation of almost a third of the general population2, the ability to cause recurrent infections, and antibiotic resistance, S. aureus treatment is increasingly challenging. In some reports, recurrent S. aureus infections have been associated with the emergence of small colony variants (SCVs), a slow-growing subpopulation of bacteria3,4. Although there are conflicting reports about the antibiotic susceptibility and comparative virulence of S. aureus SCVs, reduced antibiotic susceptibility in SCVs associated with prolonged antibiotic therapy such as aminoglycosides has been reported5. In addition to reduced growth rates, the SCV phenotype may be modulated during infection and the expression of virulence factors regulated accordingly, thus promoting persistence and survival within host cells6,7,8,9.
According to previous reports, S. aureus can form SCVs following exposure to sub-lethal concentrations of the phenolic biocide, triclosan10,11, which is commonly used for disinfection and hygiene purposes. Triclosan has broad-spectrum antibacterial activity, with the major mode of action involving the disruption of fatty acid biosynthesis through the inhibition of the enoyl-acyl protein reductase enzyme FabI at bacteriostatic concentrations12. According to some reports, triclosan also directly targets the bacterial cell membrane13,14 although disruption in fatty acid synthesis may also indirectly affect cell membrane integrity.
The formation of gentamicin-induced S. aureus SCVs has been attributed to a mutation in the electron transport chain genes, menD and hemB7 resulting in auxotrophy for these growth factors. In contrast, triclosan-induced SCVs may not be deficient in any of these growth factors and as such their emergence could be due to impaired energy production10.
The chacteristics of triclosan-induced SCV have been previously investigated for S. aureus ATCC 683511 where the SCVs showed reduced pathogenicity in an invertebrate model and exhibited decreased hemolysis, coagulase and DNase production11. This triclosan-induced SCV also exhibited defects in cell division overexpression of the triclosan target enzyme (FabI)15. However, whilst the emergence of S. aureus SCVs during antibiotic exposure has been previously investigated, comparatively little is known about the mechanisms resulting in the formation of SCVs in response to triclosan, or the implications of adaptation. The aim of the current study therefore was to explore genomic alterations and reversibility in triclosan-adapted S. aureus SCVs. Additionally, to expand the previous observations11,15, relative pathogenicity and cell invasion were assessed in a panel of SCVs and a non-SCV forming strain. Protein expression was also investigated to gain further insights into the phenotypic alterations associated with triclosan adaptation.
Results
Development of the SCV phenotype
After triclosan exposure (P10), 4/5 strains (ATCC 43300, NCTC 13277, SAR17 and Newman) formed the SCV phenotype as evidenced by the formation of pinpoint colonies. The colony morphology of strain SAR2831 remained unchanged. In all cases, the SCV phenotype was unstable, reverting following five or more passages in the absence of triclosan.
Mutations in S. aureus SCVs
Single non-synonymous mutations in the fabI gene were observed in P10 and PX10 SCVs for Newman, NCTC13277 and SAR17 strains (Table 1), where cytosine (C) changed to thymine (T) leading to an amino acid change (A95V). This mutation was sustained when triclosan was removed (PX10) although compensatory mutations were not detected. ATCC 43300 exhibited a different fabI single nucleotide polymorphism (G113C). A mutation was also identified in the fabD gene of both ATCC 43300 (V111D) and the Newman strain (Q228K) following exposure to triclosan. The latter mutation was also observed in the Newman strain after recovery in the absence of triclosan (PX10). Teicoplanin resistance associated membrane gene (tcaA) exhibited a mutation (G373V) in the triclosan exposed non-SCV strain SAR2831which was also observed in PX10.
Triclosan induced SCVs revert with fatty acid supplementation
The Newman S. aureus strain and the SCV derived from ATCC43300 showed the largest reduction in colony size when comparing P10 to the original parent strains (21% and 24% respectively; p < 0.001) while SCV derived from SAR17 and NCTC13277 colonies were almost 50% (p < 0.02) the size of the wild-type strain. When SCVs were supplemented with fatty acids, they underwent full phenotypic reversion (p < 0.05) in the SCV phenotype (Fig. 1).

Colony size of P0 and SCVs (P10) with and without the addition of fatty acids. All P10 SCVs showed reduced colony size compared to P0. When the media supplemented with fatty acids, all SCV colony size reverted to pre-exposure size (P0). *Significant change (p < 0.05) compared to P0 and FA stands for media supplemented with fatty acids (Tween 80). Results are means and standard error from representative colonies (n = 3).
Reduced triclosan susceptibility in S. aureus SCV
Table 2 illustrates the susceptibility of S. aureus to triclosan following triclosan exposure. Strains exhibiting the SCV phenotype (derived from ATCC 43300, NCTC 13277, SAR17 and the Newman strain) exhibited a significant decrease (p < 0.05) in sensitivity to triclosan at MIC and MBC. This contrasts with the non-SCV strain, SAR2831, which exhibited no significant change in either MIC or MBC to triclosan. This alteration in susceptibility was sustained even when triclosan was removed (PX10) in terms of both MIC and MBC.
Reduced relative pathogenicity in triclosan exposed isolates
Results obtained from the relative pathogenicity assessment are shown in Fig. 2. Overall, 3/5 triclosan adapted S. aureus strains (ATCC 43300, Newman and SAR17) exhibited a reduced G. mellonella larvae virulence in comparison to their unexposed counterparts. Pathogenicity for SCV ATCC 43300 (p < 0.001), SAR17 (p < 0.0001) and Newman (p < 0.001) were significantly reduced when compared to the parent strains, which reverted to pre-exposure levels for Newman and ATCC43300 but not for SAR 17 following repeated growth in the absence of triclosan. NCTC 13277 (SCV) and SAR2831 (non-SCV) mutants did not exhibit any significant difference in pathogenicity when compared to their respective parent strain.

Survival curve of four SCVs (NCTC 13277, ATCC 43300, SAR17 and Newman) and SA2831 (non-SCV). All SCVs P10 strains exhibited reduced larval lethality compared to P0 with exception of NCTC 13277. ATCC 43300, Newman and SAR17 P10 exhibited a significant reduction in relative pathogenicity, and PX10 pathogenicity reverted to P0 level with exception of SAR17, which remain attenuated even when triclosan was removed (PX10). *Significant change (p < 0.05) compared to P0. Results are means from three separate experiments.

Proteins altered in 3 SCVs (Newman, ATCC 43300 and SAR17) and non-SCV (SAR2831) compared to P0 categorised based on the function or pathway. Newman exhibited the highest number in protein expression alteration while ATCC 43300 and SAR 17 showed changes in 26 and 48 proteins, respectively.
Alterations in the S. aureus proteome following triclosan exposure
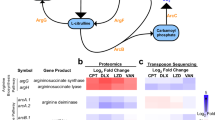
Proteins exhibiting a 2-fold or more significant change in expression were investigated for changes in the proteome. Following triclosan exposure, 50 proteins were significantly (p < 0.05) up-regulated and approximately 55 proteins (p < 0.05) were down-regulated in the Newman strain (Fig. 3). While in SAR17, 28 proteins showed increased expression and 20 showed downregulated expression. ATCC 43300 exhibited the lowest number of protein changes with 8 proteins becoming overexpressed and 18 under-expressed. In the non-SCV forming strain (SAR2831), 20 proteins were significantly up-regulated and approximately 15 proteins were down-regulated (Fig. 3). Protein expression data were categorised into eight groups based on protein function or pathway (Fig. 3). Although the majority of altered proteins in the PX10 isolates reverted to pre-exposure levels in Newman and ATCC 43300, SAR17 PX10 proteins failed to revert to P0 levels.
In terms of virulence associated proteins, in the Newman strain, proteins which play a role in S. aureus virulence, including response regulator SaeR, Immunodominant Antigen B, Gamma-Hemolysin Component A, Immunoglobulin-binding Protein sbi and Leukocidin/Hemolysin Toxin Family S, were down- regulated while response regulator protein GraR and drug resistance transporter EmrB/QacA were up-regulated. In SAR17, Immunoglobulin-Binding Protein sbi, Immunoglobulin G and Immunodominant Antigen B were upregulated; however, /Serine-Rich Adhesin for platelets SraP and Response Regulator Protein GraR were over-expressed. None of these proteins showed a significant change in ATCC 43300 except for EmrB/QacA that exhibited an up-regulation. In the non-SCVs (SAR2831) Gamma Haemolysin and Immunoglobulin G were downregulated.
In Newman and SAR17 (SCVs) as well as the non SCV bacterium SAR2831, fatty acid biosynthesis proteins were upregulated including the triclosan target enzyme enoyl-[acyl-carrier-protein] reductase FabI12 which was increased more than 4-fold, 13-fold and 6- fold, respectively. Several further fatty acids synthesis associated enzymes such as oxoacyl-[acyl-carrier-protein] synthase 2 were also up-regulated in both Newman and ATCC43300 strains (3.3 and 2.1-fold, respectively). The SAR17 and Newman SCV exhibited a greater than 2-fold increase in expression of oxoacyl-[acyl-carrier-protein] synthase 3 enzyme (FabH). Malonyl CoA-acyl transacylase, which is involved in fatty acid metabolism, was down regulated in both ATCC43300 and Newman after triclosan exposure (5 and 2.4-fold, respectively).
In the Newman strain, expression of stress-associated proteins such as threonine dehydratase II and pyruvate formate-lyase-activating enzyme were reduced. Alanine dehydrogenase was down- regulated in Newman, SAR17 and SAR2831 SCVs. Several peptidoglycan biosynthesis enzymes such as transglycosylase SceD and peptidoglycan hydrolase, as well as teichoic acid biosynthesis enzymes and cell membrane proteins were upregulated in the Newman SCV. Bifunctional autolysin was over-expressed in all strains except for ATCC43300. Transmembrane and cell division proteins were however, less expressed in all SCVs. Similarly, DNA synthesis and replication proteins exhibited a reduction in the SCVs as well as the non-SCV triclosan adapted strain. However, DNA binding and urease accessory proteins showed up-regulation following triclosan exposure in SCVs Newman and SAR17 and SAR2831. Transport proteins showed a variation in expression with the majority being downregulated while those associated with amino acids synthesis were overexpressed in all strains. SAR17, ATCC 43300 and SAR2831, showed an increase expression of 50 S ribosomal protein while carbamoyl-phosphate synthase was overexpressed in SAR 17. Several other cell metabolism proteins also exhibited different expression when exposed to triclosan (Supplement 1).
Reduced keratinocyte invasion in triclosan exposed isolates
SCVs, strain Newman (p < 0.001), ATCC 43300 (p < 0.01) and SAR17 as well as non-SCV SAR2831 (p < 0.05) showed a significant reduction in the number of recoverable bacteria after keratinocyte infection when compared to their unexposed counterparts (Fig. 4). In the recovered strains (PX10), the number of recoverable Newman and SAR 2831 reverted to unexposed levels however, SAR17 PX10 failed to revert to levels comparable to pre-exposure baselines (p < 0.05) (Fig. 4).

Keratinocytes invasion assay of P0, P10 and PX10 in four SCVs (NCTC 13277, ATCC 43300, SAR17 and Newman) and SA2831 (Non-SCV). ATCC 43300, SAR17, Newman and SAR2831 P10 exhibited a significant reduction in recovered bacteria and number of intracellular reverted to unexposed level when triclosan was removed. *Significant change (p < 0.05) compared to P0. Results are means and standard error from three separate experiments with two technical replicates.
Discussion
SCVs are a slow growing subpopulation of bacteria that can form in response to an environmental stresses including exposure to certain antibiotics and the biocide triclosan16. The induction of SCV by exposure to triclosan has been previously observed10 and we have reported attenuated virulence11 and reduced competitive fitness15 in SCV induced by this compound. The main purpose of the current investigation was to study the genomic changes underlying the formation and reversion of SCV following exposure to triclosan. In pursuing this aim we have expanded the panel of test bacteria used in previous studies to include clinical isolates and assessed the correlation between an invertebrate virulence assay and a keratinocyte invasion assay to assess the potential implications of the triclosan induced SCV. We have used high-resolution proteomics (LC-MS) to further investigate proteomic changes in SCV and established the SCV phenotype can be reversed by supplementation with fatty acids.
Five strains of S. aureus including MRSA were repeatedly exposed to sub-lethal concentrations of triclosan using a previously validated gradient plate method11. Following ten passages in the presence of triclosan, S. aureus strains demonstrated reduced triclosan susceptibility (Table 2) with 4/5 developing the SCV phenotype, suggesting that reduced triclosan susceptibility did not depend on the formation of SCVs. Reductions in triclosan sensitivity have been previously observed in other S. aureus SCVs15. At the genomic level a nonsynonymous fabI (A95V) mutation was identified in all the SCVs (Table 1) except for ATCC 43300 that instead exhibited (G113C) mutation in the fabI gene. This mutation was sustained when triclosan was removed (PX10) although compensatory mutations were not detected suggesting that with the SCV phenotype is not directly caused by fabI mutation or that phenotypic adaptation overcome the physiological effects of the mutation following repeated growth in the absence of the antimicrobial.
A (A95V) mutation in the FabI-triclosan binding site17 has been previously reported in triclosan adapted S. aureus18 and may contribute to the decrease in triclosan susceptibility (Table 2). According to some previous reports, in lipid-rich environments, Gram positive bacteria can overcome fatty acid inhibition by utilizing exogenous fatty acids19. We therefore investigated whether the SCV phenotype could be reversed by lipid supplementation. Interestingly, all SCV colonies increased in diameter to pre-exposure size immediately during growth with fatty acid supplementation (Fig. 1), suggesting that triclosan-induced SCVs are deficient in fatty acid synthesis. Since triclosan is lipophilic it is notable that reversal of the SCV phenotype occurred in adapted bacteria growing in the absence of triclosan and thus, partitioning of the antimicrobial into the supplemental lipids is unlikely to be responsible.
Despite their association with persistent infections, reductions in virulence have been previously reported in SCVs generated following antibiotic exposure6,7,8,9,20 or through sublethal exposure to the biocide triclosan in S. aureus ATCC 683511. In the current study, relative pathogenicity was further investigated in a collection of triclosan adapted SCV, as well as non-SCV forming strain. In agreement with a previous investigation11, S. aureus SCVs exhibited a reduction in relative pathogenicity, evidenced by a reduced G. mellonella virulence when compared to parent strains (Fig. 2). G. mellonella is a useful model to investigate S. aureus pathogenesis and virulence21 because it has an innate immune system22. Moreover, SCVs have been previously associated with recurrent and persistent infections and have been shown to exhibit an increased ability to adhere to and invade host cells in addition to a reduced antibiotic susceptibility8. To gain further insight into the pathogenic potential of the adapted bacteria the ability of triclosan induced S. aureus to invade primary human keratinocytes was assessed, showing a reduction in recoverable bacteria in 3/4 SCVs as well as in the non-SCV forming strain (Fig. 4). Reduced intracellular persistence may be attributed to down regulation in virulence proteins (immunoglobulin-binding protein sbi) as well as a reduced growth rate evidenced by down expression in nucleic acids synthesis and cell division proteins (FtsL and FtsZ) (Supplement 1). Moreover, reduction in cell division protein (Supplement 1) may account for formation of the SCV phenotype and suggest that the bacteria are defective in cell division which correlates with our previously reported TEM analysis15.
SCV persistence has been associated with overexpression of fibronectin-binding protein FnBP8. In the current study, low FnBP expression in Newman SCV may explain the reduction in intercellular persistence because (FnBP) is crucial for bacterial host cell invasion23. FnBP also participates in adhesion formation (as reviewed by Josse et al.)24, therefore, a reduction in FnBP expression in triclosan adapted SCVs may partly explain impaired biofilm formation observed previously11. Newman SCV also exhibited low expression in both response regulator SaeR and histidine protein kinase SaeS proteins (Supplement 1), which are regulated by the Sae gene25. Sae controls the expression of many virulence genes such as fnbA and hIg, encoding fibronectin-binding and gamma-hemolysin proteins, respectively26. These proteins were down regulated in our Newman strain (Supplement 1). Down regulation of sae has been reported in hemB deficient SCVs27, thus attenuated virulence in our Newman SCV may be due to a down regulation in the sae gene. Moreover, 2 /4 SCV exhibited down-regulation of immune response proteins, immunodominant antigen B (Supplement 1), which could explain the reduced relative pathogenicity in triclosan-induced SCVs.
Proteomic data revealed an upregulation in the triclosan target enzyme FabI in two SCVs as well as the non-SCV forming strain (Supplement 1). This may be attributed to non-synonymous mutation in fabI in response to triclosan exposure (Table 1). Formation of such a mutation is proposed to be a compensatory mechanism whereby S. aureus attempts to compensate for fatty acid depletion28. Moreover, cell membrane proteins exhibited up-regulation, such as, EmrB/QacA a membrane fusion efflux pump protein, suggesting a possible link between efflux pump expression and decreased triclosan susceptibility in S. aureus.
Although SAR2831 did not exhibit the SCV phenotype following ten passages of triclosan exposure, there was a significant reduction in the number of recoverable bacteria in the keratinocyte invasion assay (Fig. 4) as well as a down-regulation in fibrinogen-binding protein FIB_STAAE (Supplement 1). Proteins associated with energy production (nucleoside diphosphate kinase and riboflavin biosynthesis protein) (Supplement 1) were down regulated in the non-SCV suggesting that reduced invasion observed in this bacterium might partially be attributed to reduced cell metabolism. This suggests that triclosan exposure effects S. aureus pathogenicity regardless of the formation of the SCV phenotype potentially through the down-regulation of the proteins. SAR2831 (non-SCV) showed a mutation in tcaA (teicoplanin resistance associated membrane protein) despite no difference being detected in MICs to teicoplanin before and after exposure to triclosan (data not shown).
Conclusion
In this investigation, all triclosan-induced SCVs exhibited mutation in fabI, which encodes the Enoyl-[acyl-carrier-protein] reductase, the major target for triclosan. Triclosan-exposed S. aureus showed reduced pathogenicity and keratinocyte invasion regardless of the SCV phenotype, which may be attributed to a reduction in the expression of cell adhesion-associated proteins following triclosan adaptation. The potential involvement of deficiencies in fatty acid biosynthesis in triclosan SCV was evidenced by the reversal of the phenotype through fatty acid supplementation although fabI mutations persisted following reversion.
Materials and Methods
Bacterial strains and growth conditions
Five different S. aureus strains were used in this study. Two clinical wild type isolates obtained from the Molecular Diagnostics and Personalised Therapeutics Unit at the University of Hail (Hail, Saudi Arabia). Strain ATCC 43300 (MRSA) was acquired from the American Type Culture Collection while strains Newman and NCTC 13277 (MRSA) were supplied by Public Health England (Salisbury, UK). Bacteria were maintained on Tryptone Soya Agar (TSA) and Tryptone Soya Broth (TSB) which were purchased from Oxoid (Basingstoke, UK) and were sterilised at 121 °C and 15 psi for 15 minutes before use.
Whole genome sequencing and data analysis
DNA extraction and whole genome sequencing (WGS) of P0, P10 and PX10 was provided by MicrobesNG using Illumina HiSeq platform. Sequences (length 2 × 250 bp paired-end reads) were analysed through several pipelines; starting by using Kraken to identify the closest reference genome, which confirmed that all sequences are S. aureus. The data were then de novo assembled using the SPAdes (http://bioinf.spbau.ru/spades) followed by variant calling against the closest reference genome (MRSA252 -UID57839, Newman-UID58839, ECT R2 -UID159389 and T0131-UID159861 for ATCC 43300, NCTC 13277, Newman, SAR17 and SAR2831, respectively). Nonsynonymous mutations were identified by comparing P10 and PX10 genomes to the untreated parent isolate (P0).
Exposure to sub-lethal concentrations of triclosan and fatty acid supplementation
Reproducible concentration gradients of triclosan (100 µg/ml to 10 mg/ml) were spread on TSA using a Wasp II spiral plater (Don Whitley, Shipley, United Kingdom)11,15 and allowed to dry for 30 min at room temperature. Following this, overnight suspensions of S. aureus were radially inoculated onto plates in triplicate and incubated for 4 days (37 °C). After 4 days, a heavy inoculum was taken from the edge of the sub-lethal zone and inoculated onto a fresh plate containing the same (or higher) concentration of triclosan. This was passaging process was repeated a further ten times to produce P10. Following this, a further ten passages were performed in the absence of triclosan to create PX10. All stocks were cryopreserved at −80 °C for further testing. For the fatty acid supplementation experiment, MHA was supplemented with Tween 80 (0.1%) and plates were incubated at 37 °C for 24 h. After sample incubation, the plates were photographed, and images were imported into the ImageJ software. Three random colonies diameter were measured, and data presented as a percentage to pre-exposure strain (P0). Parent strains were also incubated with and without fatty acids to ensure that the increase in the colony size is not due to a simple nutritional response.
Minimum Inhibitory Concentrations and Minimum Bactericidal Concentrations
MICs were determined using the broth microdilution assay as described previously29. Briefly, overnight bacterial cultures were diluted in TSB to an OD600 of 0.8 and then further diluted 1: 100 in TSB. Adjusted cultures were then transferred to a 96 well microtitre plate containing doubling dilutions of triclosan. The microdilution plates were incubated at 37 °C for 24 h. The MIC was defined as the lowest concentration of the antimicrobial that inhibits the bacterial growth. Bacterial growth was determined by visual inspection for turbidity in the well. Aliquots (5 μL) were transferred from wells showing no growth and placed into sterile TSA plates and incubated at 37 °C (24 h) to measure MBCs. MBC was defined as the lowest concentration of the antimicrobial required to show no growth after 4 days of incubation.
Galleria mellonella pathogenesis assay
The pathogenesis assay was adapted from11. Final stage G. mellonella larvae were purchased from Live Foods Direct (Sheffield, United Kingdom) and stored in dark for a maximum of 7 days. 10 larvae (15–25 mm in length) were selected for each condition (P0, P10 and PX10). Overnight S. aureus cultures were grown in TSB and washed twice in sterile Phosphate Buffer Solution (PBS) then diluted to OD600 0.3. Hamilton syringe were used to injected hemocele larvae through left proleg with the washed bacterial suspension (5 µl) (0.8–2.6 × 106 CFU/Larvae which confirmed colony count in TSA). The syringe was washed by ethanol (100%) twice before injecting another group. Each group (10 larvae) were placed in sterile petri dish and incubated at 37 °C. A control group (not injected) and a PBS injected group were also incubated as a negative control. Survival of each group were recorded daily for 6 days and larvae considered dead when appear black or no response seen when touch. The experiment was performed three independent times and if two or more of the control larvae died, the experiment was repeated again. Results were presented in survival curve and log-rank test were calculated using Prism v.7 (www.graphpad.com/).
Keratinocyte invasion assay
Invasion assays were performed as described by30. Normal primary human epidermal keratinocytes (NHEK) were purchased from Promocell (Germany) and were grown in 24 well-plate containing keratinocyte growth medium-2 (1 ml) (Promocell, Germany) until 90% confluence (3–5 × 105 cells/well). Overnight S. aureus suspension was diluted in keratinocyte growth medium-2 to OD600 0.4 (1–2.5 × 108 CFU/ml). Bacteria were added at a keratinocyte: bacteria ratio of 1:100 and incubated for 2 h at 37 °C. Keratinocytes were washed twice with sterile PBS (1 ml) followed by addition of gentamicin (100 µl/ ml) and lysostaphin (10 µl/ml) to kill any unattached or loosely adhered bacteria. To lyse keratinocytes, distilled water (100 µl) and trypsin (100 µl) were added and serial dilutions of the lysed suspensions were plated on TSA, before incubation overnight at 37 °C and determining of remaining viable bacteria. Assay was performed in triplicate for each isolate (P0, P10 and PX10) and student t-test was used to determine significance between recoverable levels of bacteria.
Protein extraction and preparation for mass spectrometry
All chemicals used were from Sigma Aldrich (Dorset, UK) unless otherwise stated. SCV that exhibited attenuated relative pathogenicity and significant reduction in recovered bacteria, were included for further protein analysis. Six replicates of P0, P10 and PX10 were grown in TSB overnight (37 °C, 100 Samples were prepared for prepared for mass spectrometry using a modified Filter-aided sample preparation (FASP) method31 with the following modifications: The cultures were pelleted by centrifugation at 4000 rpm for 10 minutes, the supernatant was decanted and the pellet was washed twice in PBS containing 1% protease inhibitors. The washed pellet was resuspended in 1 mL of PBS with lysostaphin (10% v/v) and kept on ice for 15 minutes. The washed cells were transferred to Zirconia/Silica bead-containing tubes (Stratech Scientific, UK) and lysed in a FastPrep 120 (Thermo Scientific, UK) at 40 rpm for 20 seconds, with 20 seconds cooling, for a total of 3 cycles. Sodium Dodecyl Sulphate (SDS) (1%) and dithiothreitol (DTT) were added to a final concentration of 1% (w/v) and 50 mM respectively. The samples were incubated at 90 °C for 5 minutes. The lysate was centrifuged at 10,000 g for 5 minutes and the supernatant was transferred to clean tube and the protein concentration was measured using a Direct Detect Spectrometer (Merck Millipore).
For FASP, clarified lysate (25 µg) was added to 100 µL of wash buffer (8 M urea, 0.1 Tris-HCl (pH 8.5) with 15 mM DTT) and washed twice in a 30 kDalton filter (Merck Millipore). To alkylate the sample, 50 µL of buffer (8 M urea, 0.1 M Tris-HCl (pH 8.5) with 0.05 M iodoacetamide) was added to the filter and the sample was incubated in darkness at room temperature for 30 minutes. The filters were washed twice with washing buffer 1 followed by a further two washes of washing buffer 2 (6 M urea, 0.1 Tris-HCl (pH 8.5) with 25% deionised water) before dilution in 50 mM Tris-HCl (pH 8.5) to bring the urea concentration to 1 M. Samples were digested by the addition of trypsin at a protein: enzyme ratio of 1:100 and incubated overnight at 37 °C. After digestion, peptides were collected by centrifugation at 4000 g at 20 °C for 15 minutes and the filtration units were washed once with washing buffer 2 and subsequently with two washes of 40 mM ammonium bicarbonate. Peptide samples were transferred to 96 well plate with 0.2 µM PVDF membrane (Corning) and desalted with OLIGO R3 reversed-phase media (Applied Biosystems). Desalted peptides were dried down and resuspended in 5% acetonitrile with 0.1% formic acid and analysed by LC-MS/MS using an UltiMate® 3000 Rapid Separation LC (RSLC, Dionex Corporation, Sunnyvale, CA) coupled to an Orbitrap Elite (Thermo Fisher Scientific, Waltham, MA) mass spectrometer. Peptide mixtures were separated using a gradient from 92% A (0.1% FA in water) and 8% B (0.1% FA in acetonitrile) to 33% B, in 44 min at 300 nL min-1, using a 250 mm × 75 μm i.d. 1.7 mM BEH C18, analytical column (Waters). Peptides were selected for fragmentation automatically by data dependant analysis (DDA).
The acquired MS data was analysed using Progenesis LC-MS (v4.1, Nonlinear Dynamics). The retention times in each sample were aligned using one LC-MS run as a reference, then the “Automatic Alignment” algorithim was used to create maximal overlay of the two-dimensional feature maps. Features with charges ≥+ 5 were masked and excluded from further analyses, as were features with less than 3 isotope peaks. The resulting peak lists were searched against the UniProt database with Staphylococcus aureus (Strain Newman) sequences included (UniProt, download date: 2016/02/26) using Mascot v2.5.1, (Matrix Science). Search parameters included a precursor tolerance of 5 ppm and a fragment tolerance of 0.5 Da. Enzyme specificity was set to trypsin and one missed cleavage was allowed. Carbamidomethyl modification of cysteine was set as a fixed modification while methionine oxidation was set to variable. The Mascot results were imported into Progenesis LC-MS for annotation of peptide peaks. Proteins exhibiting 2 or more-fold significant changes (at a p < 0.05) were included in the study.
Data availability
All data generated or analysed during this study are included in this published article (and its Supplementary file).
References
Chambers, H. F. & DeLeo, F. R. Waves of Resistance: Staphylococcus aureus in the Antibiotic Era. Nat Rev Microbiol 7, 629–641 (2009).
Kluytmans, J., van Belkum, A. & Verbrugh, H. Nasal carriage of Staphylococcus aureus: epidemiology, underlying mechanisms, and associated risks. Clin Microbiol Rev 10, 505–20 (1997).
Kahl, B. C. Small colony variants (SCVs) of Staphylococcus aureus - A bacterial survival strategy. Infect Genet Evol 21, 515–522 (2014).
Kahl, B. et al. Persistent infection with small colony variant strains of Staphylococcus aureus in patients with cystic fibrosis. J Infect Dis 177, 1023–9 (1998).
vonEiff, C. et al. A site-directed Staphylococcus aureus hemB mutant is a small-colony variant which persists intracellularly. J Bacteriol 179, 4706–4712 (1997).
Tuchscherr, L. et al. Staphylococcus aureus phenotype switching: an effective bacterial strategy to escape host immune response and establish a chronic infection. Embo Mol Med 3, 129–141 (2011).
Proctor, R. A. et al. Staphylococcus aureus Small Colony Variants (SCVs): a road map for the metabolic pathways involved in persistent infections. Fronti Cell Infect Microbiol 4 (2014).
Kriegeskorte, A. et al. Inactivation of thyA in Staphylococcus aureus Attenuates Virulence and Has a Strong Impact on Metabolism and Virulence Gene Expression. Mbio 5 (2014).
Proctor, R. A. et al. Small colony variants: a pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat Rev Microbiol 4, 295–305 (2006).
Bayston, R., Ashraf, W. & Smith, T. Triclosan resistance in methicillin-resistant Staphylococcus aureus expressed as small colony variants: a novel mode of evasion of susceptibility to antiseptics. J Antimicrob Chemother 59 (2007).
Latimer, J., Forbes, S. & McBain, A. J. Attenuated virulence and biofilm formation in Staphylococcus aureus following sublethal exposure to triclosan. Antimicrob Agents Chemother 56 (2012).
McMurry, L. M., Oethinger, M. & Levy, S. B. Triclosan targets lipid synthesis. Nature 394 (1998).
Escalada, M. G., Russell, A. D., Maillard, J. Y. & Ochs, D. Triclosan-bacteria interactions: single or multiple target sites? Lett Appl Microbiol 41, 476–481 (2005).
Villalain, J., Mateo, C. R., Aranda, F. J., Shapiro, S. & Micol, V. Membranotropic effects of the antibacterial agent Triclosan. Arch Biochem Biophys 390, 128–36 (2001).
Forbes, S., Latimer, J., Bazaid, A. & McBain, A. J. Altered Competitive Fitness, Antimicrobial Susceptibility, and Cellular Morphology in a Triclosan-Induced Small-Colony Variant of Staphylococcus aureus. Antimicrob Agents Chemother 59, 4809–4816 (2015).
Kahl, B. C. Small colony variants (SCVs) of Staphylococcus aureus – A bacterial survival strategy. Infect Genet Evol 21, 515–522 (2014).
Xu, H. et al. Mechanism and inhibition of saFabI, the enoyl reductase from Staphylococcus aureus. Biochemistry 47, 4228–4236 (2008).
Ciusa, M. L. et al. A novel resistance mechanism to triclosan that suggests horizontal gene transfer and demonstrates a potential selective pressure for reduced biocide susceptibility in clinical strains of Staphylococcus aureus. Int J Antimicrob Agents 40 (2012).
Brinster, S. et al. Type II fatty acid synthesis is not a suitable antibiotic target for Gram-positive pathogens. Nature 458, 83–6 (2009).
Balwit, J. M., Vanlangevelde, P., Vann, J. M. & Proctor, R. A. Gentamicin-Resistant Menadione and Hemin Auxotrophic Staphylococcus-Aureus Persist within Cultured Endothelial-Cells. J Infect Dis 170, 1033–1037 (1994).
Purves, J., Cockayne, A., Moody, P. C. E. & Morrissey, J. A. Comparison of the Regulation, Metabolic Functions, and Roles in Virulence of the Glyceraldehyde-3-Phosphate Dehydrogenase Homologues gapA and gapB in Staphylococcus aureus. Infect Immun 78, 5223–5232 (2010).
Ramarao, N., Nielsen-Leroux, C. & Lereclus, D. The Insect Galleria mellonella as a Powerful Infection Model to Investigate Bacterial Pathogenesis. Jove-J Vis Exp (2012).
Nielsen, L. N. et al. Staphylococcus aureus but not Listeria monocytogenes adapt to triclosan and adaptation correlates with increased fabI expression and agr deficiency. BMC Microbiol 13, 177 (2013).
Josse, J., Laurent, F. & Diot, A. Staphylococcal Adhesion and Host Cell Invasion: Fibronectin-Binding and Other Mechanisms. Front Microbiol. 8, 2433 (2017).
Mainiero, M. et al. Differential Target Gene Activation by the Staphylococcus aureus Two-Component System saeRS. J Bacteriol 192, 613–623 (2010).
Kuroda, H., Kuroda, M., Cui, L. & Hiramatsu, K. Subinhibitory concentrations of beta-lactam induce haemolytic activity in Staphylococcus aureus through the SaeRS two-component system. Fems Microbiol Lett 268, 98–105 (2007).
Senn, M. M., Bischoff, M., von Eiff, C. & Berger-Bachi, B. SigmaB activity in a Staphylococcus aureus hemB mutant. J Bacteriol 187, 7397–406 (2005).
Morvan, C. et al. Environmental fatty acids enable emergence of infectious Staphylococcus aureus resistant to FASII-targeted antimicrobials. Nat Commun 7, 12944 (2016).
Moore, L. E., Ledder, R. G., Gilbert, P. & McBain, A. J. In vitro study of the effect of cationic biocides on bacterial population dynamics and susceptibility. Appl Environ Microbiol 74, 4825–34 (2008).
Mempel, M. et al. Invasion of human keratinocytes by Staphylococcus aureus and intracellular bacterial persistence represent haemolysin-independent virulence mechanisms that are followed by features of necrotic and apoptotic keratinocyte cell death. Br J Dermatol 146, 943–51 (2002).
Wisniewski, J. R., Zougman, A., Nagaraj, N. & Mann, M. Universal sample preparation method for proteome analysis. Nat Methods 6, 359–62 (2009).
Acknowledgements
The authors would like to thank Julian Selley and David Knight from the Biological Mass Spectrometry Core Research Facility at the University of Manchester for the support in mass spectrometry data analysis. We thank Mohammed S. Al-Mogbel and Ibraheem Ashankyty from the Molecular Diagnostics and Personalised Therapeutics Unit at the University of Hail for SAR17 and SAR2831 strains. Genome sequencing was provided by MicrobesNG (http://www.microbesng.uk), which is supported by the BBSRC (grant number BB/L024209/1). This work was supported by a PhD scholarship from University of Hail (Saudi Arabia) to Abdulrahman Bazaid.
Author information
Authors and Affiliations
Contributions
Conceived and designed the experiments: A.B., S.F., G.H., R.L., A.M. Performed the experiments: A.B. Protein extraction and preparation for mass spectrometry: R.O. All authors contributed to the final version of the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bazaid, A.S., Forbes, S., Humphreys, G.J. et al. Fatty Acid Supplementation Reverses the Small Colony Variant Phenotype in Triclosan-Adapted Staphylococcus aureus: Genetic, Proteomic and Phenotypic Analyses. Sci Rep 8, 3876 (2018). https://doi.org/10.1038/s41598-018-21925-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-21925-6
This article is cited by
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
