
Abstract
Coffee is a major tropical commodity crop that can provide supplementary habitat for native wildlife. In Asia, coffee production is an increasingly important driver of landscape transformation and shifts between different coffee species is a major dimension of agroforestry trends. Yet few studies have compared the ecological impacts of conversion between different coffee species. We evaluated whether or not the two species of coffee grown globally—Coffea arabica and C. canephora (denoted “robusta”)—had equivalent avian conservation value in the Western Ghats, India, where robusta production has become increasingly dominant. We found that habitat specialist and functional guild diversity was higher in arabica, and that arabica was more profitable. However, robusta farms generally supported the same or slightly higher abundances of habitat specialists and functional guilds, largely due to dense canopy and landscape-level forest cover. Farming practices, chiefly pesticide use, may affect the suitability of coffee agroforests as habitat for avian specialists, and at present, robusta farmers tended to use less pesticide. Given future projections for arabica to robusta conversion in tropical Asia, our study indicates that certification efforts should prioritize maintaining native canopy shade trees and forest cover to ensure that coffee landscapes can continue providing biodiversity benefits.
Similar content being viewed by others


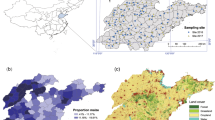
Introduction
Coffee (Coffea spp.) is one of the most valuable and widely planted tropical commodity crops with two major species in production: C. arabica (henceforth, arabica), constituting about 60% of global production, and C. canephora (robusta), around 40% of global production1. Coffee production systems range from shaded, low-intensity farming where coffee trees are interspersed with native forest, more typical of arabica production, to high-intensity full-sun monoculture characteristic of robusta agroforests. Shade-grown coffee retains biodiversity at higher levels than more intensely farmed, full-sun or monoculture systems2,3,4,5.
Shade grown coffee has declined precipitously in the past twenty years6,7, largely due to greater robusta production, particularly in Asia8,9,10,11. Price equalization between the two species and declining arabica productivity in the face of climate change may further accelerate conversion to robusta across tropical Asia12,13. Understanding habitat specialist responses to arabica versus robusta is critical as production may shift away from arabica toward robusta in many parts of the tropics.
Demand for coffee is rising much more rapidly in Asia (3.7%) than the global average (1.3%)14. While coffee production has declined in the Neotropics and Afrotropics, in South and Southeast Asia, it has increased by more than 100%7. Currently, India is the world’s sixth largest coffee producer and coffee acreage in India has increased by 150% from 1990 to 201515,16. The majority of this expansion occurred in a global biodiversity hotspot, the Western Ghats17,18,19.
Across the Western Ghats, coffee agroforest area is slightly more than a quarter of the land area that is formally protected16; whether coffee can serve as buffer habitat for wildlife will be critically important to conservation outcomes19,20. From 1950 to 2015, the planted area of robusta grew by 840% while arabica acreage increased by 327% in India16. Previous research posited that robusta expansion could be ecologically detrimental as is often grown in more open conditions where farmers fell older trees and lop more branches to open up the canopy21.
In this article, we analyze avian habitat specialist trends in arabica and robusta farms and integrate household interviews to explore what policy levers may be most powerful in securing a biodiversity-friendly future for coffee production lands. Despite coffee’s importance as a driver of landscape transformation, and the unclear impacts of robusta supplanting arabica, there have been limited studies comparing these two agroforests from a biodiversity perspective22. The majority of existing assessments have often not distinguished between coffee species, and have instead compared coffee against other crops, sacred groves, and native forests19,23,24,25,26,27,28.
Results
Arabica and robusta production practices and trends
Across the entire household survey dataset (n = 344), 213 households planted arabica and 236 planted robusta. Of the arabica planters, 50.2% solely planted arabica; for robusta planters, 55.1% solely planted robusta; 106 households grew both varieties of coffee. In 2003, 215 of the respondents produced robusta and 196 farmed arabica. Compared to this baseline, the net change in the number of farmers planting each species was a 9.3% increase for robusta and 7.7% increase for arabica.
The average planted area of arabica was 14.3 ± 0.1 hectares (n = 213, median: 4.9 ha, maximum: 242.8 ha) and 11.6 ± 0.1 hectares for robusta (n = 236, median: 6.1 ha, maximum: 121.4 ha). At the time of the study, the mean planted areas for each crop were not significantly different (W = 22510, padj = 0.2, Fig. 1A). On the other hand, the mean acreage of robusta increased significantly over the past decade (V = 977, padj = 3.7 × 10−6, Fig. 1A).

(A) Changes in planted area for arabica and robusta among the surveyed households. Notched boxplots of planted area a decade ago and during the studied period for both crops. (B) The net profits for each crop (in 2013 USD$) per unit hectare. After subtracting the cost of inputs per hectare, arabica was more profitable than robusta on average.
Both arabica and robusta stands had fairly closed canopies with a median canopy density score of 94.6% (mean: 94.6%, range: [79.7, 99.7]%) for arabica and 79.2% for robusta (mean: 77.2%; [50.4, 99.8]%). Moreover, all coffee agroforests tended to be situated in regions with dense forest cover with a median tree cover of 92.1% for arabica within a 2 km buffer around each farm (mean: 88.8%, [67.5, 99]%) and 88.9% for robusta (mean: 86.3%, [55.3, 98.7]%).
Farmers employed a variable usage rate for five surveyed inputs; for arabica, the majority of farmers used pesticides as well as organic and conventional fertilizer, while for robusta, the majority of farmers used conventional fertilizer, and less than half of the farmers used the other four inputs (Table 1). The cost per hectare spent on each type of input was wide-ranging for both coffee species (Table 1). Arabica was significantly more profitable per unit area than robusta (arabica: $1555.98 ± 2.7 versus robusta: $1439.53 ± 2.2; W = 26407, padj = 0.002; Fig. 1B).
Avian richness patterns across the two coffee species
Across the sampled agroforests, the number of forest-dependent species ranged from 22–63 with a total of 79 forest-dependent species recorded in the full dataset. Fourteen endemic species were observed with anywhere from two to seven species found in the individual agroforests. Three IUCN Red-Listed species were observed in the course of study: Alexandrine Parakeet (Psittacula eupatria, near-threatened), Grey-headed Bulbul (Pycnonotus priocephalus, near-threatened), and Nilgiri Wood-pigeon (Columba elphinstonii, vulnerable). Only three agroforests did not have a single Red-listed species present Twenty-six frugivorous, 54 insectivorous, and 26 omnivorous species were recorded.
Arabica supported more speciose assemblages of forest-dependent, endemic, frugivorous, insectivorous, and omnivorous birds (Table 2). There were nearly twice as many endemic bird species in arabica compared to robusta (Table 2, n a,j1 = 19 ± 2.2, n r,j1 = 11 ± 0). For IUCN Red-Listed species richness, arabica and robusta agroforests had indistinguishable asymptotic richness, though this analysis was limited by the small number of threatened species (Table 2).
The five most commonly observed forest-dependent, endemic, and functional guild species were similar across robusta and arabica (Appendix 1, Table A1). However, the forest-dependent Malabar grey hornbill (Ocyceros griseus), frugivorous Plum-headed parakeet (Psittacula cyanocephala), and insectivorous Oriental magpie-robin (Copsychus saularis) exhibited different patterns in commonness across the two coffee species (Appendix 1, Table A1).
Habitat specialist species accumulation and community composition
A sufficient number of endemic and forest-dependent species were observed to conduct individual-based rarefaction. The rarefaction results indicated that the initial accumulation of forest-dependent and endemic species was similar across arabica and robusta; however, the asymptotic richness of endemics and forest birds were higher in arabica and more individuals were observed for both these groups in arabica than in robusta (Fig. 2).

Individual-based rarefaction for arabica and robusta agroforests for forest-dependent (FD) and endemic species richness (ED).
A permutational MANOVA demonstrated that the forest-dependent and endemic communities were significantly different between the two coffee species (FFD = 3.95, p adj = 0.006; FED = 3.87, p adj = 0.02). For endemics and forest birds, there were no significant partitioning effects associated with environmental variables (tree richness, canopy density, and distance to protected area). Correspondence analysis (CCA) indicated that the forest-dependent and endemic species communities occupied non-overlapping ordination space (Fig. 3).

Community composition of (A) Forest Dependent birds and (B) Endemics across arabica and robusta agroforests. Ordination scores were calculated at the level of farms for each set of species using correspondence analysis. The 95% confidence ellipses for the arabica and robusta site centroids are shown in green and orange, respectively.
Abundance patterns for habitat specialists, threatened birds, and foraging guilds
Forest dependent species
The mean detection of forest-dependent individuals was 0.09 while it was 0.14 for clusters. A range of [0.4, 48.7] forest-dependent birds per hectare were detected in arabica and [0.3, 28.3] in robusta. The range of flocks per hectare (clusters) that were observed in the two coffee species were [0.4, 19.9] flocks/ha in arabica and [0.4, 15] in robusta. There were an average of 1.54 ± 0.04 birds per flock in arabica and 1.46 ± 0.03 in robusta. We found that robusta supported significantly higher densities of forest-dependent flocks, but not individual birds (Table 3).
Endemic species abundance
The mean detectability for endemic birds was 0.09 for individuals and 0.13 for clusters. The range of individual endemics observed per hectare was [0, 33.5] in arabica and [0.4, 23.3] in robusta. For flocks, the range of clusters per hectare was [0, 9.5] in arabica and [0.4, 9.3] in robusta. The typical flock size was 1.4 ± 0.07 endemic birds per flock in arabica and 1.3 ± 0.05 in robusta. The results did not indicate any significant difference between arabica and robusta in terms of endemic bird density (Table 3).
IUCN Red-Listed species
There were only 20 observations of IUCN red-listed species, preventing abundance analysis; however, a global distance analysis indicated an average detection probability of 0.27.
Foraging guilds
Across the three foraging guilds, detection probabilities for individual birds centered on 0.1 and was around 0.14 for flocks. Individual bird densities (birds/ha) across all three guilds ranged from [0.4, 38.8] in arabica and [0.4, 22.6] in robusta. Flock densities spanned 0.4 to 13.2 clusters per hectare in arabica (average flock size: 1.5 ± 0.06 birds) and 0.4–9.4 in robusta (flock size: 1.4 ± 0.04 birds). In general, there were no significant differences in the density of individual birds or flocks among the three foraging guilds (Table 3). However frugivore flock densities were significantly higher in robusta.
Relating habitat specialist diversity and functional guild abundance to ecological and land use covariates
Using a global model containing all relevant covariates (Appendix 1, Tables A2–A6), we evaluated if there was evidence of residual spatial autocorrelation for all three guilds as well as the forest-dependent and endemic species29. There was no evidence of residual spatial autocorrelation for the habitat specialists or foraging guilds (Moran’s I ranged from [−0.06, 0.02] with p ∈ [0.16, 0.41]), consistent with the findings of Karanth et al.5.
For the forest-dependent species, six of the eight candidate models garnered sufficient support for model averaging (Appendix 1, Table A2). The final ensemble model included all of the ecological covariate variables except for canopy structure and tree species richness; however, pesticide, distance to protected area, and tree cover had the highest relative variable importance scores (1, 0.82, and 0.7 respectively). Yet the confidence intervals for all variables crossed 0, indicating that none of these variables had a clear, directional effect on forest-dependent richness (Fig. 4A).

Model-averaged regression coefficients predicting (A) forest-dependent and (B) endemic species richness in arabica and robusta. The variable names are shown in column on the left. “Tree SR” represents tree species richness; “Dist. to PA” distance to protected area.
On the other hand, for the endemics, only one model was chosen under the model selection framework (weight: 0.92), as the nearest model had a ΔAICc = 5.1 (Table A2). Tree species richness, crop type, distance to protected area, and pesticide use were contained in the most-supported model. Higher tree species richness tended to decrease endemic diversity, and while there was a trend toward robusta predicting higher endemic diversity, this variable ultimately was not significant (Fig. 4B).
Across the three guilds, five to six of the candidate models were highly supported. For insectivores, the most important variables were pesticide use, distance to protected area, and tree cover (relative importance scores: 1, 0.71, 0.7) (Fig. 5B), while the most important variables for omnivore and frugivore abundance were distance to protected area, pesticide use, and type of coffee agroforest (Fig. 5A–C, scoring 1, 1, 0.6 for both guilds). Although the impact of environmental covariates on foraging guild density was generally unclear, omnivore densities significantly increased further from protected areas (Fig. 5). Moreover, while pesticide use was potentially detrimental for frugivore and insectivore abundance, it is possible that the density of omnivores would rise.

Model-averaged regression coefficients predicting (A) frugivore, (B) insectivore, and (C) omnivore density (individual birds per hectare) in arabica and robusta.
Discussion
We found lower levels of forest-dependent, endemic, and foraging guild species richness in robusta than arabica. Forest-dependent, frugivorous, insectivorous, and endemic birds are sentinels of habitat quality, and are often the first taxa to disappear from modified habitats30,31,32. Yet on the whole, compared to other major cash crops in the Western Ghats such as areca (Areca catechu) and rubber (Hevea brasiliensis), Karanth et al.5 noted that coffee—aggregated across both arabica and robusta—supported higher overall avian richness, endemic richness, and greater densities for the majority of the foraging guilds as well as three out of four vertical structure guilds (low canopy, mid-, and high-canopy guilds).
Robusta agroforests typically supported the same or slightly higher densities of habitat specialists and foraging guilds as arabica, though many of these differences were not significant. This pattern was likely due to the most common forest-dependents and endemics; the observation rates for the five most common forest and endemic bird species were similar across the arabica and robusta agroforests. Existing farming practices may contribute to this outcome; only 19% of robusta farmers used pesticides compared to 75% of the arabica farmers. Reduced pesticide use in robusta farms could lead to increased food resources for insectivore populations. Although pesticide use did not have a clear directional impact on habitat specialist and functional guild responses, it was consistently one of the most important variables.
Our result that flock densities were slightly, but significantly, higher in robusta while overall habitat specialist and foraging guild richness was lower echoes recent findings that flocking behaviors may permit for habitat specialists to use more modified or disturbed habitats. Goodale et al.28 observed significant avifaunal community turnover between forest and agricultural habitat in Southern India and Sri Lanka. However, human-altered landscapes supported similar flock densities as native forest, and these mixed-species flocks appeared to recruit more forest-interior species to these more open habitats than would be otherwise expected.
Distance to protected area did not exhibit a large effect for forest and endemic bird diversity. Moreover, there was evidence that frugivore and omnivore abundance may still be high far from protected areas, suggesting that coffee agroforests can produce substantial economic and biodiversity benefits as buffers3,5,33,34. The pattern we observed is likely not an artifact of the range of distances to protected areas. Across the agroforests, the range of distances to protected areas extended from 0–35.9 km in arabica and 0.4–34.6 km in robusta; the maximum distances were four times farther than a comparable study in the Western Ghats26.
The importance of coffee agroforests as supplementary habitat may be heightened in regions with small protected areas embedded in human-use landscapes5,19,20,23. In other parts of tropical Asia where farming practices are more dissimilar to native forest, distance to protected area is often a significant predictor with a large impact on bird diversity35,36.
Arabica yielded higher profits per-hectare than robusta. Yet the planted area statistics indicated that robusta production has increased over the past decade. Indian robusta is distinguished by its high cup quality and resistance to disease, rendering it an attractive crop to farmers37. Additionally, certain robusta varieties are approaching price equalization with arabica12,14,15,16,37. As such, it is encouraging that robusta agroforests are capable of supporting abundant avian populations, both in terms of habitat specialists and foraging guilds. Nevertheless, these communities are less speciose than arabica assemblages.
Although our survey data did not include direct measurements of yield, there was no indication that reported pesticide use—an important determinant of production intensity—significantly affected endemic or forest-dependent avian diversity. Previous work in Southeast Asia focusing on cacao noted that yield did not necessarily correlate negatively with reduced conservation value34.
Managing shade tree species composition and landscape forest cover appear to be major levers for improving the biodiversity conservation value of coffee agroforests. In fact, the surveyed robusta agroforests possessed canopy and forest cover three times higher than shade-grown coffee farms in Indonesia and instead scored similarly to forest plots in Bukit Barisan Selatan reserve10. In general, the high prevalence of shade-grown coffee differentiates Indian arabica and robusta production globally, driven by historical concerns about coffee rust (Hemileia vastatrix)6,38. Ensuring the persistence of extensive forest cover at landscape scales and dense canopies of native trees would present two practical guidelines for certifying both arabica and robusta6,10,26,33. In highly populated landscapes such as the Western Ghats in India, and other rural tropical regions, it is critically important that calls to conserve wildlife within human-altered landscapes offer meaningful pathways to improve local people’s livelihoods that respect their aspirations3,15,33,39.
Certification efforts in Southeast and South Asia have largely relied on price signals which have had an equivocal impact on biodiversity conservation6,11,19,25,38,39,40. Across tropical Asia, there are repeated instances of rising coffee prices leading to clearance and conversion of protected areas or opening up of the canopy as select trees are cut down in bad years by farmers11,41. Wide variation across certification standards could actually incentivize the removal of shade trees that are critical for retaining habitat specialist vertebrates23,24,42. In fact, we observed that higher tree species richness tended to decrease the diversity of endemic birds. This is likely driven by smallholders planting exotic trees as an additional source of income; a high diversity of exotic tree species at the expense of native shade trees can be disruptive for sensitive avifauna26,27.
Unfortunately, recent efforts by the Rainforest Alliance to certify coffee production in the Western Ghats did not increase the conservation value of these lands; certified farmers retained on average 100 fewer native trees per hectare than non-certified producers15. Frequent audits and a requirement for bookkeeping may engender future hostility toward conservation interventions in this landscape, as producers expressed disappointment in measureable outcomes and certification’s limited environmental management requirements. Our research emphasizes the importance of practical recommendations for both birds and farmers.
Despite the shortcomings highlighted by Bose et al.15, due to the small median landholdings in this landscape, successful certification efforts in the Western Ghats would provide a unique and meaningful opportunity to identify management factors that are a triple win for poverty alleviation, human well-being, and conservation38,39,40,41. Certification often poses insurmountable financial demands for the smallest and most cash-poor farm holdings to demonstrate adherence to ecological or livelihood targets6,11,38.
Carbon credits as well as more rigorously audited and locally tailored management schemes could help ensure that certified coffee would be both livelihood and wildlife friendly39,40,41,42,43,44,45. It is evident that wildlife certification schemes should use scientific assessments of wildlife and be developed locally to truly enhance the value of existing coffee production systems and promote regional biodiversity. Continued work in tropical production landscapes should seek to quantify the relationship between yield, crop type, planting practice (canopy cover, tree density, retention of native trees) and a broader suite of habitat specialist taxa.
Methods
Social survey data
We surveyed 344 coffee agroforest owners across the three highest growing districts in Karnataka: Chikmagalur, Hassan and Kodagu5. More than 75% of farms in the region are <10 hectares in size. The farms selected comprised 113 arabica growers, 135 robusta growers, and 96 growing both varieties. The surveys were carried out by six trained research assistants between June 2013-July 2014.
The survey covered household demographics and socio-economics such as family size, education, income, farm size, and characteristics. Farmers were questioned about their coffee growing history in 2003, a decade before the study occurred. We also obtained details about yield and farm management, such as tree species grown, crop varieties planted, shade management and chemical inputs as well as access to institutions. Coffee production areas ranged from 2 to 250 hectares, and the range of arabica and robusta planted area was [0, 242.8 hectares] and [0, 121.4 hectares], respectively.
Ecological data
61 coffee agroforests were surveyed for avian diversity with 30 in arabica and 31 in robusta5. In each farm, all point count stations were placed in only one of the crop types if both crops were grown on the farm. A minimum distance of 1 km was maintained between each sampled agroforest. Sampling occurred during the dry season (January to May 2013) and the surveys were conducted between 6:30–9:30 a.m., and 4:00–6:30 p.m, maximizing detection and visibility for passerines and near-passerines.
We evaluated bird occurrence using point counts. The number of points per farm was proportional to the size of the agroforest, ranging from 2 to 9. Each point was spaced 200 m apart for quasi-independence. At each point, two trained observers recorded all birds that were heard or sighted for 7 min after an initial wait period of 2 min to minimize the effect of disturbance. The sighting distance to each bird or clusters of birds was measured. Each point was revisited six times over three days to achieve adequate numbers of detections46. We sampled a total of 274 points in coffee. We excluded migrant species from our analysis to avoid biasing overall richness and density estimates. We grouped avian species into the following categories based on published sources: forest-dependent47,48,49, IUCN Red-Listed49,50, endemic24,50, and three foraging guilds—frugivorous, insectivorous, and omnivorous51 (Appendix II).
We examined differences between arabica and robusta management practices such as shade tree retention and species composition, coffee tree spacing, and coppicing as well as variable levels of nutrient, water and pesticide inputs (Robbins et al., in review). We also measured several covariates potentially associated with species occurrence including elevation, slope, weather, canopy structure, canopy density, presence of leaf litter, presence of water bodies and pesticide use5. Slope was measured using a compass. Canopy density was measured in all four cardinal directions at each point using a canopy densiometer. The point-centered quarter method was used to estimate tree densities at each point52,53.
Analyses
Socioeconomic data
We calculated the mean, median, and range for planted area under each coffee species as well as the inputs used for each crop type and tree cover statistics. We identified the rate of change in acreage and proportion of planted land allocated to each coffee species. We evaluated differences in means using Wilcoxon Rank Sum tests. We applied Bonferroni familywise-error adjustment.
The avifaunal point count stations were matched to farms using a unique identifier, in order to associate the point count locations with environmental and farming practices covariates.
Ecological data
The asymptotic richness and community composition of forest-dependent, endemic, and threatened birds as well as the three foraging guilds was calculated using the package vegan (v 2.4.1) in R (v 3.3.1) at the level of individual agroforests and across crop types54,55. We determined the abundance of individual birds and flocks using the package Distance (v 0.9.6) in R56. Crop type (arabica versus robusta) was defined as the region, and the total area for each region was summed across all point count stations within each crop type, assuming that each point count had a radius of 100 m. The individual samples were the point count stations; as such, effort was the number of visits to each station. Truncation was performed at 100 m.
We used asymptotic richness estimates as response variables for the habitat specialists, while the estimated densities of foraging guilds were used as the modelled response. We constructed generalized linear models with a Gaussian error distribution to ascertain the relationship between avian habitat specialist diversity or foraging guild abundance and several habitat and farming practice covariates. All predictor variables were normalized and showed no evidence of multicollinearity. For the habitat specialist richness regressions, the survey effort at each farm (the number of point count stations per farm in this case) was supplied as an offset. Model suitability was visually assessed using diagnostic residual, Q-Q, and leverage plots.
We performed multimodel inference to evaluate empirical support for several hypotheses related to crop type, non-coffee tree cover and species richness, non-coffee tree density, canopy density, canopy structure, distance to the nearest protected area, and pesticide usage. A total of eight candidate models were compared using the package MuMIn in R (v 1.15.6)57. We performed full model averaging with a shrinkage estimator across the most parsimonious candidates (ΔAICc ≤4)58. We evaluated whether or not there was evidence of residual spatial autocorrelation using Moran’s I with the R packages ape (v 4.1) and geosphere (v 1.5-5)59,60.
Data archiving statement
The ecological data have been formatted for replication analyses in R, and saved as an.Rdata file with an accompanying R script for running the relevant analyses, as well as a description of each object. These objects can be accessed in the article’s Supplementary Information and are mirrored at https://github.com/charlottehchang/WCS-India-Coffee. The appendix details how to access the replication data and perform analyses. Due to legal and ethical constraints given the sensitivity of surveying farmers in India, we are unable to provide the socio-economic household data.
Change history
11 April 2018
A correction to this article has been published and is linked from the HTML and PDF versions of this paper. The error has not been fixed in the paper.
References
Food and Agriculture Organization of the United Nations (FAO). FAO Statistical Pocketbook–Coffee 2015 (2015).
Perfecto, I., Rice, R. A., Greenberg, R. & Van der Voort, M. A. Shade Coffee: A Disappearing Refuge for Biodiversity. BioScience 46, 598–608 (1996).
Bhagwat, S. A., Willis, K. J., Birks, H. J. B. & Whittaker, R. J. Agroforestry: A Refuge for Tropical Biodiversity? Trends Ecol. Evol. 23, 261–67 (2008).
Fischer, J. et al. Should Agricultural Policies Encourage Land Sparing or Wildlife-Friendly Farming? Front. Ecol. Environ. 6, 380–85 (2008).
Karanth, K. K. et al. Producing Diversity: Agroforests Sustain Avian Richness and Abundance in India’s Western Ghats. Front. Ecol. Evol. 4, 111 (2016).
Jha, S. et al. A Review of Ecosystem Services, Farmer Livelihoods, and Value Chains in Shade Coffee Agroecosystems in Integrating Agriculture, Conservation and Ecotourism: Examples from the Field (eds Campbell, W. B. & López Ortíz, S.) 141–208 (Springer, 2011).
Jha, S. et al. Shade Coffee: Update on a Disappearing Refuge for Biodiversity. BioScience 64, 416–28 (2014).
D’haeze, D. J., Deckers, D., Raes, D., Phong, T. A. & Loi, H. V. Environmental and Socio-Economic Impacts of Institutional Reforms on the Agricultural Sector of Vietnam: Land Suitability Assessment for Robusta Coffee in the Dak Gan Region. Agr. Ecosyst. Environ. 105, 59–76 (2005).
Guingato, P., Nardone, E. & Notarnicola, L. Environmental and Socioeconomic Effects of IntensiveAgriculture: The Vietnam Case. J. Commod. Sci. Tech. Qual. 47, 135–151 (2008).
Philpott, S. M., Bichier, P., Rice, R. A. & Greenberg, R. Biodiversity Conservation, Yield, and Alternative Products in Coffee Agroecosystems in Sumatra, Indonesia. Biodivers. Conserv. 17, 1805–20 (2008).
Meyfroidt, P., Vu, T. P. & Hoang, V. A. Trajectories of Deforestation, Coffee Expansion and Displacement of Shifting Cultivation in the Central Highlands of Vietnam. Global Env. Chang. 23, 1187–1198 (2013).
Ghoshray, A. On price dynamics for different qualities of coffee. Rev. Market Intg. 1, 103–118 (2009).
Ovalle-Rivera, O. et al. Projected shifts in Coffea arabica suitability among major global producing regions due to climate change. PLoS One 10(4), e0124155 (2015).
International Trade Centre. The Coffee Exporter’s Guide, 2016 (2016).
Bose, A., Vira, B. & Garcia, C. Does environmental certification in coffee promote “business as usual”? A case study from the Western Ghats, India. Ambio 45, 946–955 (2016).
Indian Coffee Board. Database on Coffee, July 2016 (2016).
Myers, N., Mittermeier, R. A., Mittermeier, C. A., Da Fonseca, G. A. B. & Kent, J. Biodiversity Hotspots for Conservation Priorities. Nature 403, 853–858 (2000).
Robbins, P., Chhatre, A. & Karanth, K. Political ecology of commodity agroforests and tropical biodiversity. Conserv. Lett. 8, 77–85 (2015).
Anand, M. O., Krishnaswamy, J., Kumar, A. & Bali, A. Sustaining Biodiversity Conservation in Human-Modified Landscapes in the Western Ghats: Remnant Forests Matter. Biol. Conserv. 143, 2363–74 (2010).
Ranganathan, J., Krishnaswamy, J. & Anand, M. O. Landscape-Level Effects on Avifauna Within Tropical Agriculture in the Western Ghats: Insights for Management and Conservation. Biol. Conserv. 143, 2909–2917 (2010).
DaMatta, F. M. Ecophysiological Constraints on the Production of Shaded and Unshaded Coffee: A Review. Field Crops Res. 86, 99–114 (2004).
Upendranadh, C. & Subbaiah, C. A. Labour shortage in coffee plantation areas in Globalisation, Development and Plantation Labour In India (eds Joseph, K. J. & Viswanathan, P. K.) 167-195 (Routledge, 2016).
Bhagwat, S. A., Kushalappa, C. G., Williams, P. H. & Brown, N. D. A Landscape Approach to Biodiversity Conservation of Sacred Groves in the Western Ghats of India. Conserv. Biol. 19, 1853–62 (2005).
Raman, T. R. S. Effects of Habitat Structure and Adjacent Habitats on Birds in Tropical Rainforest Fragments and Shaded Plantations in the Western Ghats, India in Forest Diversity and Management (eds Hawksworth, D. L. & Bull, A. T.) 517-547 (Springer, 2006).
Bali, A., Kumar, A. & Krishnaswamy, J. The Mammalian Communities in Coffee Plantations Around a Protected Area in the Western Ghats, India. Biol. Conserv. 139, 93–102 (2007).
Anand, M. O., Krishnaswamy, J. & Das, A. Proximity to Forests Drives Bird Conservation Value of Coffee Plantations: Implications for Certification. Ecol. Appl. 18, 1754–63 (2008).
Ambinakudige, S. & Sathish, B. N. Comparing Tree Diversity and Composition in Coffee Farms and Sacred Forests in the Western Ghats of India. Biodivers. Conserv. 18, 987–1000 (2009).
Goodale, E. et al. The Response of Birds and Mixed-Species Bird Flocks to Human-Modified Landscapes in Sri Lanka and Southern India. Forest Ecol. Manag. 329, 384–392 (2014).
Kühn, I. & Dormann, C. F. Less than eight (and a half) misconceptions of spatial analysis. J. Biogeogr. 39, 995–998 (2012).
Castelletta, M., Sodhi, N. S. & Subaraj, R. Heavy Extinctions of Forest Avifauna in Singapore: Lessons for Biodiversity Conservation in Southeast Asia. Conserv. Biol. 14, 1870–80 (2000).
Harvey, C. A. & González Villalobos, J. A. Agroforestry Systems Conserve Species-Rich but Modified Assemblages of Tropical Birds and Bats. Biodivers. Conserv. 16, 2257–2292 (2007).
Şekercioğlu, Ç. H., Loarie, S. R., Oviedo Brenes, F., Ehrlich, P. R. & Daily, G. C. Persistence of Forest Birds in the Costa Rican Agricultural Countryside. Conserv. Biol. 21, 482–494 (2007).
Garcia, C. A. et al. Biodiversity Conservation in Agricultural Landscapes: Challenges and Opportunities of Coffee Agroforests in the Western Ghats, India. Conserv. Biol. 24, 479–88 (2010).
Clough, Y. et al. Combining High Biodiversity with High Yields in Tropical Agroforests. Proc. Natl Acad. Sci. 108, 8311–8316 (2011).
Peh, K. S. H., de Jong, J., Sodhi, N. S., Lim, S. L. H. & Yap, C. A. M. Lowland rainforest avifauna and human disturbance: persistence of primary forest birds in selectively logged forests and mixed-rural habitats of southern Peninsular Malaysia. Biol. Conserv. 123, 489–505 (2005).
Zhang, M., Chang, C. H. & Quan, R. Natural forest at landscape scale is most important for bird conservation in rubber plantation. Biol. Conserv. 210, 243–252 (2017).
Prakash, N. S., Devasia, J., Jayarama & Aggarwal, R. K. Coffee Industry in India: Production to Consumption—A Sustainable Enterprise in Coffee in Health and Disease Prevention (ed. Preedy, V. R.) 61–70 (Academic Press, San Diego, 2015).
Neilson, J. et al. Challenges of Global Environmental Governance by Non-State Actors in the Coffee Industry: Insights from India, Indonesia and Vietnam in Agriculture, Biodiversity and Markets: Livelihoods and Agroecology in Comparative Perspective (eds Lockie, S. & Carpenter, D.) 175–200 (Earthscan, 2010).
Bawa, K. S., Joseph, G. & Setty, S. Poverty, Biodiversity and Institutions in Forest-Agriculture Ecotones in the Western Ghats and Eastern Himalaya Ranges of India. Agr. Ecosyst. Environ. 121, 287–295 (2007).
Gobbi, J. A. Is Biodiversity-Friendly Coffee Financially Viable? An Analysis of Five Different Coffee Production Systems in Western El Salvador. Ecol. Econ. 33, 267–281 (2000).
Gaveau, D. L. A. et al. Three Decades of Deforestation in Southwest Sumatra: Effects of Coffee Prices, Law Enforcement and Rural Poverty. Biol. Conserv. 142, 597–605 (2009).
Noponen, M. R. A., Haggar, J. P., Edwards-Jones, G. & Healey, J. R. Intensification of Coffee Systems Can Increase the Effectiveness of REDD Mechanisms. Agr. Syst. 119, 1–9 (2013).
Beuchelt, T. D. & Zeller, M. Profits and Poverty: Certification’s Troubled Link for Nicaragua’s Organic and Fairtrade Coffee Producers. Ecol. Econ. 70, 1316–24 (2011).
Chiputwa, B., Spielman, D. J. & Qaim, M. Food Standards, Certification, and Poverty Among Coffee Farmers in Uganda. World Dev. 66, 400–412 (2015).
Perfecto, I., Vandermeer, J., Mas, A. & Soto Pinto, L. Biodiversity, Yield, and Shade Coffee Certification. Ecol. Econ. 54, 435–446 (2005).
Buckland, S. T. et al. Introduction to distance sampling: estimating abundance of biological populations (Oxford University Press, 2001).
Beukema, H., Danielsen, F., Vincent, G., Hardiwinoto, S. & Van Andel, J. Plant and Bird Diversity in Rubber Agroforests in the Lowlands of Sumatra, Indonesia. Agroforest. Syst. 70, 217–42 (2007).
Del Hoyo, J., Elliott A. & Christie D. Handbook of the Birds of the World: HBW Alive (Lynx Edicions 2016).
BirdLife International. Bird Life Data Zone http://datazone.birdlife.org/home (2017).
LePage, D. Avibase–the World Bird Database https://avibase.bsc-eoc.org (2016).
Wilman, H. et al. EltonTraits 1.0: Species‐level foraging attributes of the world’s birds and mammals. Ecology 95, 2027–2027 (2014).
Runkle, J. R. Canopy tree turnover in old-growth mesic forests of eastern North America. Ecology 81, 554–567 (2000).
Mitchell, K. Quantitative Analysis by the Point-Centered Quarter Method (Hobart and William Smith Colleges, 2007).
Oksanen, J. et al. vegan: Community Ecology Package. R package version 2.4-1, https://CRAN.R-project.org/package=vegan (2007).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. Available at: https://www.R-project.org/. (2016).
Miller, D. L. Distance: Distance Sampling Detection Function and Abundance Estimation. R package version 0.9.6. Available at, https://CRAN.R-project.org/package=Distance (2016).
Bartoń, K. MuMIn: Multi-Model Inference. R package version 1.15.6., https://CRAN.R-project.org/package=MuMIn (2016).
Burnham, K. P. & Anderson, D. R. Model selection and multimodel inference: a practical information-theoretic approach (Springer-Verlag, 2002).
Paradis, E., Claude, J. & Strimmer, K. APE: analyses of phylogenetics and evolution in R language. Bioinformatics 20, (289–290 (2004).
Hijmans, R. J. geosphere: Spherical Trigonometry. R package version 1, 5–5 (2016).
Acknowledgements
Funding for this project was provided by US NSF Grant Number 1265223 (PR, KKK, AC Chhhatre), Oracle (KKK), the US NSF Graduate Research Fellowship (CHC) and Graduate Research Opportunities Worldwide with USAID program (CHC). We gratefully acknowledge support from the Wildlife Conservation Society, Centre for Wildlife Studies, Princeton University, University of Wisconsin-Madison, University of Illinois-UC, and Indian School of Business. We thank Vishnupriya Sankararaman and Shashank Dalvi for leading the field efforts, and volunteers who data. The authors gratefully acknowledge the 187 farm owners who participated in our study, especially B. M. Akarsha, S. Hulikere, H. B. Rajgopal, H. B. Arjun, M. Nanjappa, and A. Goel whose advice and help was invaluable. The authors thank A. Chhatre, K. U. Karanth, N. S. Kumar, U. Srinivasan, N. Deshmukh, A. Srivathsa, V. Tripuraneni, R. Parameshawaran, A. Philip, M. Agarwala, N. Velho, N. Sekar, P. Hannam, P. Elsen, J. D. Nichols, J. Krishnaswamy, N. Page, R. Singal, B. Manjunath, R. Jhambekar, J. Seshadri, D. Jois, T. Gopalaswamy, B. Ramesh, K. Hirdode, S. Nambiar, A. Mendis, P. Modi, S. Prabhu, and A. Jamalabad for helpful discussions and support.
Author information
Authors and Affiliations
Contributions
C.H.C. and K.K.K. designed and executed the analysis, and led the writing of the manuscript; K.K.K. and P.R. conceived the original empirical study design. All authors contributed critically to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chang, C.H., Karanth, K.K. & Robbins, P. Birds and beans: Comparing avian richness and endemism in arabica and robusta agroforests in India’s Western Ghats. Sci Rep 8, 3143 (2018). https://doi.org/10.1038/s41598-018-21401-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-21401-1
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
