
Abstract
Hymenoptera venom allergy can cause severe anaphylaxis in untreated patients. Polistes dominula is an important elicitor of venom allergy in Southern Europe as well as in the United States. Due to its increased spreading to more moderate climate zones, Polistes venom allergy is likely to gain importance also in these areas. So far, only few allergens of Polistes dominula venom were identified as basis for component-resolved diagnostics. Therefore, this study aimed to broaden the available panel of important Polistes venom allergens. The 100 kDa allergen Pol d 3 was identified by mass spectrometry and found to be a dipeptidyl peptidase IV. Recombinantly produced Pol d 3 exhibited sIgE-reactivity with approximately 66% of Polistes venom-sensitized patients. Moreover, its clinical relevance was supported by the potent activation of basophils from allergic patients. Cross-reactivity with the dipeptidyl peptidases IV from honeybee and yellow jacket venom suggests the presence of exclusive as well as conserved IgE epitopes. The obtained data suggest a pivotal role of Pol d 3 as sensitizing component of Polistes venom, thus supporting its status as a major allergen of clinical relevance. Therefore, Pol d 3 might become a key element for proper diagnosis of Polistes venom allergy.
Similar content being viewed by others

Introduction
Stings of hymenoptera of different species can cause life-threatening IgE-mediated anaphylaxis in venom-allergic patients. The most prominent elicitors of venom allergy in Western and Central Europe are honeybees (Apis mellifera) and yellow jackets (Vespula vulgaris)1. Additionally, allergic reactions to paper wasps, especially to Polistes dominula, are common in Southern Europe and the Unites States2,3,4,5,6. Polistes dominula, known to be domestic in Southern Europe, is an invasive species entering the US (1970s) from the north-east to the west coast (1990s)7, South Africa (2008)8 and central Europe (1956)9. Therefore, allergy to Polistes dominula venom (PDV) will most probably gain importance also in other areas.
The only curative treatment for venom allergy is venom-specific immunotherapy (VIT)10,11. To ensure a successful treatment and to avoid the increased risk of side effects, possible de novo sensitizations and higher costs, the correct therapeutic venom must be selected4,12. To accomplish this, a careful anamnesis is important to identify the insect that elicited the allergic reaction. Due to the number and hard to discriminate phenotypes of insects that can induce allergic reactions, many patients and allergy specialists are not able to correctly distinguish between different hymenoptera species such as Vespula spp. and Polistes spp.13. Therefore, clinicians depend on additional diagnostic tests.
The increased knowledge of the composition of hymenoptera venoms has led to major improvements in diagnostic approaches and created the field of molecular or component-resolved diagnostics (CRD) in hymenoptera venom allergy4,14,15. In combination with skin testing and venom extract-based specific IgE (sIgE) diagnostics, CRD has created added clinical value for a proper allergy diagnosis. For CRD of hymenoptera venom allergy commercial allergens are available for different test platforms to determine sIgE serum titers6.
For the diagnosis and discrimination of honeybee venom (HBV)- and yellow jacked venom (YJV)-allergic patients, many commercial allergens are available, allowing for high diagnostic sensitivity and specificity16,17,18,19,20,21,22. However, for the diagnosis of PDV allergy only one allergen (antigen 5, Pol d 5) is commercially available. Furthermore, only three allergens of PDV have been identified in the past, namely phospholipase A1 (Pol d 1)23, protease (Pol d 4)24 and antigen 5 (Pol d 5)25 from which Pol d 4 is a minor allergen with restricted diagnostic importance (unpublished data). Even though, we have recently shown, that Polistes venom is free of cross-reactive carbohydrate determinant-(CCD-)based cross-reactivity26, available extract-based diagnostic approaches to discriminate between PDV und YJV allergy are hampered by extensive protein cross-reactivity2,3,27,28.
An increased knowledge of the allergen composition of PDV and the availability of further important PDV components are likely to generate added clinical benefit for proper and advanced diagnostics. Therefore, our study aimed to identify and immunologically characterise additional major allergens of PDV. The here described allergen Pol d 3 is a homologue of the prominent dipeptidyl peptidase IV (DPP IV) allergens Api m 5 and Ves v 3 from HBV and YJV29. Extensive sIgE reactivity and the ability to activate basophils from allergic patients clearly support the role of Pol d 3 as major allergen of PDV as well as its potential to be a key element for molecular diagnostic approaches.
Results
Identification of Pol d 3
Immunoblots using PDV and pooled sera of PDV-allergic patients (n = 5) revealed sIgE reactivity with several venom components (Fig. 1). Prominent bands at approximately 23 and 31 to 40 kDa were identified by mass spectrometry as the known allergens antigen 5 (Pol d 5), protease (Pol d 4) and phospholipase A1 (Pol d 1), respectively. Moreover, prominent sIgE reactivity was obtained with a high molecular weight protein of approximately 100 kDa. The protein was subjected to de novo sequencing by tandem mass spectrometry. The five identified peptides (Fig. 2) yielded hits in a database search of predicted proteins coded by the recently published genome of Polistes dominula30 and identified the sIgE-reactive protein as venom dipeptidyl peptidase IV (GenBank accession XP_015174445). An additional matrix-assisted laser desorption ionization-time of flight (MALDI-TOF) analysis of the protein band then led to a sequence coverage of approximately 44% with the predicted sequence.

Detection of Pol d 3 in Polistes dominula venom. SDS-PAGE and protein staining (SyproRuby staining) of PDV (left) and sIgE-immunoreactivity of pooled sera from PDV-allergic patients with PDV in Western blot (right). The arrow indicates the 100 kDa band that was subjected to tandem mass spectrometry and MALDI-TOF analyses. Asterisks indicate the band that was identified as Pol d 5 and black bars the area of the gel/blot in which Pol 1 and Pol d 4 were identified. Shown are parts of the gel. Blot and full-length gels are shown in Supplemental Fig. S3. The kDa values correspond to the protein marker (not shown) which can be found in Fig. S3.

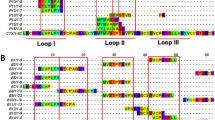
Alignment of Pol d 3 with Ves v 3 and Api m 5. Mature amino acid sequences of Pol d 3 (Polistes dominula), Ves v 3 (Vespula vulgaris) and Api m 5 (Apis mellifera) are shown. Black shaded amino acids are identical between all three proteins, gray shaded amino acids are shared by two proteins and amino acids not shaded are unique to the individual protein. Peptides identified by tandem mass spectrometry are underlined in black and potential N-glycosylation sites in gray. The residues involved in the conserved active center of the enzymes are represented boxed. Overall amino acid identity between the different proteins is stated in percent.
The nucleotide sequence (GenBank accession XM 015318959) of the newly identified allergen codes a mature protein of 751 amino acids and a calculated molecular weight of 86.2 kDa. The discrepancy between the calculated and the apparent molecular mass most likely is due to posttranslational modification by glycosylation as suggested by the presence of 4 or 3 N-linked glycosylation sites based on the binary profile or on average surface accessibility, respectively31. The protein belongs to the dipeptidyl peptidase IV superfamily, known to cleave dipeptides from the N-terminus of peptidic substrates, including many chemokines, neuropeptides, peptide hormones and venom peptides32,33. Therefore, it is a homologue of the well-established allergens Api m 5 (HBV) and Ves v 3 (YJV)29. The identity on protein level with Api m 5 and Ves v 3 is 54.0% and 76.6%, respectively. According to its allergenic properties and the homology to YJV Ves v 3 the new PDV allergen was assigned as Pol d 3.1010 in the WHO/IUIS Allergen Nomenclature Database34.
Recombinant expression and characterisation of Pol d 3
For recombinant expression, the coding region of Pol d 3 was amplified from PDV gland cDNA. Recombinant production was achieved by baculoviral infection of Sf9 insect cells and Ni2+-affinity chromatography yielded soluble recombinant Pol d 3 from culture supernatants with an apparent molecular weight of approximately 100 kDa as shown by Coomassie staining and reactivity with an antibody reacting with the V5-epitope tag added for recombinant expression (Fig. 3). Moreover, correct folding of the allergen was supported by enzymatic DPP IV activity (Supplementary Fig. S1) as well as by circular dichroism (CD) spectroscopy (Supplementary Fig. S2). For comparable analyses the homologous allergens from HBV and YJV, Api m 5 and Ves v 3, were produced as described previously29 (Fig. 3).

Recombinant expression and characterisation of Pol d 3. SDS-PAGE and Western blot analyses of Pol d 3 recombinantly produced in Sf9 insect cells in comparison with the HBV and YJV homologues Api m 5 and Ves v 3 either by Coomassie blue staining or anti-V5 epitope antibody, GNA (Galanthus nivalis agglutinin), anti-HRP antiserum, anti-Api m 5 antiserum and pooled sera of PDV-allergic patients. Shown are parts of one or more gels and blots and full-length gels and blots are given in Supplemental Fig. S3.
Reactivity with Galanthus nivalis agglutinin (GNA) (reactive with terminal 1,2-. 1,3- and 1,6-linked mannose residues) proves the presence of N-linked glycans (Fig. 3). However, staining with rabbit polyclonal horseradish peroxidase (HRP) antiserum, specific for α-1,3-core fucosylation, the structure responsible for CCD-based cross-reactivity, demonstrates the lack of CCDs (Fig. 3), as shown previously for other allergens produced in Sf9 insect cells18,35,36,37.
Furthermore, staining with a polyclonal rabbit Api m 5 antiserum shows reactivity with Api m 5 but also with Ves v 3 and Pol d 3, suggesting cross-reactivity between the three DPP IV hymenoptera allergens. Cross-reactivity was confirmed by sIgE-reactivity of pooled sera of PDV-allergic patients with all three allergens (Fig. 3).
Activation of basophils from venom-allergic patients by Pol d 3
To address the capability of Pol d 3 to cross-link receptor-bound IgE and, thus, to activate effector cells, basophil activation tests (BATs) were performed (Fig. 4) as previously described28,38. First, 13 patients from Spain (from the area of Barcelona) with history of an allergic reaction to PDV and/or YJV were analysed for their reactivity with Pol d 3 and Ves v 3 (Fig. 4a). Three patients showed basophil activation only by Pol d 3 (patients 3, 7, 8), two only by Ves v 3 (patients 1, 11) and four by both allergens (patients 4, 9, 10, 13). Interestingly, for most of the patients these data nicely match the data of skin tests and/or sIgE levels to the venom extracts (i3, i77) and allergen components (i209, i210, i211) (Table 1). Therefore, for most patients BATs with the DPPs IV would have been able to identify the insect(s), the patient most likely shows primary sensitization to. For patient 4, showing stronger basophil activation by Pol d 3 compared to Ves v 3, but higher sIgE levels to YJV compared to PDV, clinical information is too scarce to identify the allergy-eliciting venom. Moreover, patient 13 exhibits basophil activation by both allergens but much higher sIgE level to PDV compared to YJV extract and a negative skin test to YJV. However, for this patient basophil activation by Pol d 3 is much stronger compared to Ves v 3, and Ves v 3-reactivity is most likely due to cross-reactivity. For patients 9 and 10 who show comparable activation patterns by both allergens, also clinical data suggest allergy to both venoms.

Basophil activation tests of (a) PDV- and/or YJV-allergic patients from Spain (area of Barcelona) and (b) HBV- and/or YJV-allergic patients from Germany with recombinant DPP IV allergens Pol d 3, Ves v 3 or Api m 5. Basophils were exposed to different concentrations of the DPP IV allergens. Additionally, stimulation with anti-FcεRI antibody (positive control) and plain stimulation buffer (negative control) is shown. Activation is shown as percentage of CD63+ out of total basophilic cells. The cut-off of the assay (10%) is represented as dotted line.
In order to address cross-reactivity of Pol d 3 in BAT, 15 patients from Germany (from the area of Munich) with allergy to YJV and/or HBV were analysed for their reactivity with Pol d 3, Api m 5 and Ves v 3 (Fig. 4b). Five patients showed basophil activation by Pol d 3 (patients 15, 16, 18, 19, 28) six by Api m 5 (patients 15, 17, 18, 21, 27, 28) and seven by Ves v 3 (15, 18, 19, 20, 24, 26, 28). Again, in most cases activation patterns by Api m 5 and Ves v 3 match sIgE and skin test data, indicating allergy to HBV, YJV or both (Table 1). Three of the Pol d 3-reactive patients, showed basophil activation by all three allergens (patients 15, 18, 28) and one by Pol d 3 and Ves v 3 (patient 19). Only patient 16 showed exclusive basophil activation by Pol d 3, but the overall activation (including the positive control) was very low.
sIgE-reactivity of Pol d 3
The sIgE to the newly identified Pol d 3 was addressed by ELISA. 24/30 of the patients from Spain (from the area of Cordoba) were diagnosed with PDV allergy by a combination of clinical history, skin test and sIgE levels to venom extracts and allergen components (Supplementary Table S1). For the remaining 6 patients (10, 11, 15, 23, 26, 30) diagnostic results were less clear and, hence, allergy to PDV and YJV cannot be excluded. In the group of Spanish patients 20/30 (66.7%) exhibited pronounced sIgE-reactivity with Pol d 3 (Fig. 5a).

sIgE reactivity of individual hymenoptera venom-allergic patients with recombinant DPP IV allergens in ELISA. (a) sIgE immunoreactivity of PDV- (n = 30), HBV- (n = 28) and YJV-allergic patients (n = 20) with Pol d 3. (b) Comparative sIgE immunoreactivity of Pol d 3-positive HBV-allergic patients with Pol d 3 and Api m 5. (c) Comparative sIgE immunoreactivity of Pol d 3-positive YJV-allergic patients with Pol d 3 and Ves v 3. The lower end cut-off of the ELISAs is represented by dotted lines.
In order to address sIgE cross-reactivity of Pol d 3 in HBV- and YJV-allergic patients from Germany (from the area of Munich), reactivity with the recombinant allergen was assessed. The patient groups were selected either for primary sensitization to HBV or YJV by detailed clinical characterisation (Supplementary Table S1). Nevertheless, in these groups allergy to both species cannot be fully excluded. However, since PDV allergy in Germany is virtually not present, primary sensitization to PDV can be excluded with high probability. 9/28 (32.1%) of HBV- and 14/22 (63%) of YJV-allergic patients showed reactivity with Pol d 3, respectively (Fig. 5a). Interestingly, most of the HBV- and YJV-allergic patients with sIgE to Pol d 3 also exhibited sIgE to the homologous allergens from HBV (Api m 5) (Fig. 5b) and YJV (Ves v 3) (Fig. 5c), respectively. Only for one patient with HBV allergy and for 3 YJV-allergic patients with very weak reactivity to Pol d 3, reactivity with the homologous allergens was slightly below the cut-off of the ELISA.
Discussion
Polistes dominula is one of the main elicitors of hymenoptera venom allergy in Southern Europe as well as in parts of the United States. Moreover, Polistes dominula is a very invasive species spreading from the warmer to the more moderate climate zones and is therefore likely to gain importance also in other areas. However, compared to other species such as Apis mellifera or Vepula vulgaris, the knowledge of the composition of important PDV allergens on a molecular level is restricted. Additionally, to date the availability of diagnostic tools for the discrimination between PDV and YJV allergy is very limited. Therefore, our study aimed to identify and immunologically characterise novel important allergens of PDV to extend the available repertoire of venom allergens for analyses on a molecular level.
In this study, we were able to identify the sIgE-reactive 100 kDa allergen of PDV as dipeptidyl peptidase IV and homologue of the well-established DPP IV allergens Api m 5 and Ves v 3 of HBV and YJV, which were shown to represent relevant allergens of clinical importance18,29,39. Protein sequence identity with the corresponding allergens of HBV and YJV is 54% and 76.1%, respectively. Due to its allergenic properties and homology to the YJV allergen Ves v 3 the new PDV allergen was assigned as Pol d 3.1010 to the IUIS/WHO allergen nomenclature database34.
In order to immunologically characterise Pol d 3, the protein was recombinantly produced as soluble and properly folded protein in Sf9 insect cells. Lectin-staining confirmed that Pol d 3 is a glycoprotein, a fact that explains the difference between the calculated molecular mass of the polypeptide chain of 86.2 kDa and the apparent molecular mass of approximately 100 kDa observed in SDS-PAGE and immunoblot analyses. However, recombinant Pol d 3 was devoid of CCD-reactivity as shown previously for other allergens produced in Sf9 insect cells29,35,36,37. Intriguingly, the reactivity of Pol d 3 with a polyclonal Api m 5-specific antiserum40 hinted to a pronounced protein-based cross-reactivity of the DPP IV allergens of the different hymenoptera species. This cross-reactivity was further confirmed by the reactivity of a serum pool from Pol d 3-reactive PDV-allergic patients with the DPP IV allergens Api m 5 and Ves v 3.
To access the relevance of Pol d 3 as allergen, sIgE-reactivity of PDV-allergic patients with this newly identified PDV component was addressed. Thereby, over 66% of PDV-allergic patients exhibited pronounced sIgE to Pol d 3. This is in a comparable range found for the reactivity of either HBV- or YJV-allergic patients with the corresponding DPP IV allergens Api m 5 and Ves v 3, respectively. In previous studies, it was demonstrated that 58.3% to 61.7% of HBV-allergic patients and 57% of YJV-allergic patients show sIgE to Api m 5 and Ves v 3, respectively18,29,39. These data clearly suggest that Pol d 3 represents a major allergen of PDV. As Polistes dominula and Vespula vulgaris coexist in Spain and are difficult to discriminate, systemic reactions due to both insects cannot be excluded in the Spanish patient population.
Additionally, 32% of HBV- and 63% of YJV-allergic patients exhibited sIgE to the new PDV allergen. The majority of these patients additionally showed reactivity with the homologous allergen of HBV (Api m 5) or YJV (Ves v 3), indicating that the sIgE to Pol d 3 is a result of extensive protein-based cross-reactivity. Moreover, the lower degree of cross-reactivity between Pol d 3 and Api m 5 compared to Pol d 3 and Ves v 3 most likely reflects the lower sequence identity between the PDV and HBV allergen and, thus, of less conserved IgE epitopes.
So far phospholipases A1 (Pol d 1 and Ves v 1) and antigens 5 (Pol d 5 and Ves v 5) are well established cross-reactive allergen pairs of PDV and YJV2,3,28,41. Although it is likely that also the hyaluronidases of PDV and YJV are cross-reactive, no reliable data exist. Moreover, at least YJV hyaluronidase seems to be of limited clinical relevance37,42. In this study DPP IV allergens were identified as novel pair of cross-reactive major allergens, responsible for the frequently observed double-positive sIgE test results with PDV and YJV. Additionally, our analyses provide for the first time a molecular basis for the observed cross-reactivity between PDV and HBV43. Therefore, DPP IV allergens might be pan-allergens, present in various hymenoptera venoms. Also, the presence of DPPs IV in many snake venoms44 suggests functions of this enzyme class in the venoms of phylogenetically distinct species.
The capacity of Pol d 3 to activate effector cells and to initiate an allergic response was assessed by basophil activation testing and proved its role as potent allergen. In the Spanish patient group the basophil activation patterns by Pol d 3 and Ves v 3 matched the data of skin testing and sIgE levels to allergen extracts and components. For the German patients, this was also the case for the basophil activation data obtained with Api m 5 and Ves v 3. However, in this patient group Pol d 3 clearly demonstrated to be cross-reactive also in BAT. For these patients PDV allergy can be excluded with high probability since it is virtually not present in Germany. Therefore, Pol d 3-reactivity in BAT in these patients most likely is due to cross-reactivity. This is further supported by the fact that most Pol d 3-reactive patients additionally show basophil activation in response to Ves v 3 (and Api m 5). Only one patient showed weak basophil activation in response to Pol d 3 only but also very low general activation according to the positive control. However, if these results are of clinical relevance is difficult to determine since diagnostic sting provocation testing is ethically not justifiable. Nevertheless, the data demonstrate that BATs with recombinant cross-reactive major allergens represent a helpful tool to identify the allergy-eliciting venom as shown previously28.
To date, in clinical practice the discrimination between allergy to PDV and YJV is quite challenging. In contrast, for the discrimination between allergy to YJV and HBV an extended component-resolved diagnostics has evolved and has shown to provide added benefit for clinical decisions4. It was proposed that PDV and YJV allergy should be discriminated by measurement of the level of sIgE to phospholipases A1 (Pol d 1 and Ves v 1) and antigens 5 (Pol d 5 and Ves v 5)3. However, in addition to the YJV allergens, so far only Pol d 5 is available for routine diagnostics. In a former study, we were able to demonstrate that for most of the patients, for whom the allergy-relevant venom was clearly identified, the sIgE level with the appropriate antigen 5 (Pol d 5 or Ves v 5) indeed was higher28. However, this does not hold true for all patients. Moreover, for many patients the sIgE levels are in a very comparable range, hence, results will be difficult to interpret in many cases.
Of course, it would be of major interest to identify species-specific marker allergens that would allow a reliable and easy discrimination between PDV and YJV allergy. However, all identified major allergens of the two venoms, including phospholipases A1, antigens 5 and dipeptidyl peptidases IV, exhibit a high degree of cross-reactivity. Hence, the development of a component-resolved diagnostic approach, comparable to that for the discrimination between HBV and YJV allergy is difficult to realize, in our opinion. Nevertheless, the extension of the available panel of allergen components also for routine diagnostics of PDV (and YJV) allergy will generate added value for advanced diagnostics. Certainly, the combination of the results of sIgE measurements to more than one major allergen will help to create a clearer diagnostic picture and to facilitate diagnostic decisions also in vespid venom allergy. Therefore, the newly identified major allergen of PDV, Pol d 3, together with its counterparts of YJV and HBV, might become a key element for molecular diagnostics of hymenoptera venom allergy. Moreover, the detailed knowledge of the allergen composition of different insect venoms will help to understand the immunological mechanisms of venom allergy and therapeutic outcome.
Methods
Patients
Blood and/or sera of 108 patients with allergy either to PDV, HBV and/or YJV were analysed. 65 patients were from the area of South Bavaria (Munich, Germany), 30 patients were from the area of Córdoba, Spain and 13 patients were from Barcelona, Spain. As PDV allergy is almost not present in Germany, allergic reactions to this species can be excluded with high probability and the German patients were allergic to YJV and/or HBV. Patients from Córdoba were primarily allergic to PDV and patients from Barcelona were allergic to PDV and/or YJV. As Polistes dominula and Vespula vulgaris coexist in Spain and are difficult to discriminate, systemic reactions due to both insects cannot be excluded.
The diagnosis of venom allergy was based on a combination of clinical history of an allergic sting reaction, a positive intradermal skin test, and/or positive sIgE levels to PDV, YJV and/or HBV (i77, i3, i1) and allergen components (i208, i209, i210, i211) (UniCAP250; Thermo Fisher Scientific, Uppsala, Sweden).
All patients had given informed written consent to draw additional blood samples. The study was approved by the ethics committee of the Faculty of Medicine of the Technical University of Munich (protocol number 5478/12), the ethics committee for clinical research of Reina Sofía University Hospital Cordoba (protocol number BLA-VIT-2015-01) and the ethical committee for clinical investigation of the Hospital Clinic of Barcelona (protocol number 2011/6605). All patients were recruited from clinical routine and the obtained data are not part of another study reported elsewhere. All experiments were performed in accordance with relevant guidelines and regulations.
Protein biochemistry
Pol d 3 was identified by tandem mass spectrometry analyses on a MALDI-TOF instrument. A detailed description is given in the Supplementary Methods.
Cloning and recombinant production of venom dipeptidyl peptidases IV
The coding region of Pol d 3 was amplified from Polistes dominula venom gland cDNA and plasmids coding for Api m 5 and Ves v 3 were generated as described previously29,45. Cloning and recombinant production in Spodoptera frugiperda (Sf9) insect cells is described in detail in the Supplementary Methods.
Immunoreactivity of patient sera with recombinant dipeptidyl peptidases IV
sIgE immunoreactivity of sera with the recombinant venom DPPs IV was assessed by ELISA. A detailed description of the ELISA is given in the Supplementary Methods. The lower end functional cut-off, indicated as dotted lines, was calculated as the mean of the negative controls summed with 3 times the standard deviation (SD) of the mean and additionally 10% of the resulting value.
Basophil activation test
Basophil activation tests were performed in 15 YJV- and/or HBV-allergic patients, and in 13 PDV- and/or YJV-allergic patients as described previously38, using the Flow CAST (Bühlmann Laboratories AG, Schönenbuch, Switzerland). Allergen concentrations were 2, 10, 50 and 250 and 1000 ng/mL. A detailed description is given in the Supplementary Methods.
Other methods
SDS-PAGE, Western blotting, DPP IV activity and CD spectroscopy are described in the Supplementary Methods.
Data availability
All data generated or analysed during this study are included in this published article and its Supplementary Information files.
References
Bilo, B. M. et al. Diagnosis of Hymenoptera venom allergy. Allergy 60, 1339–1349 (2005).
Caruso, B. et al. Wasp venom allergy screening with recombinant allergen testing. Diagnostic performance of rPol d 5 and rVes v 5 for differentiating sensitization to Vespula and Polistes subspecies. Clin Chim Acta 453, 170–173 (2016).
Monsalve, R. I. et al. Component-resolved diagnosis of vespid venom-allergic individuals: phospholipases and antigen 5s are necessary to identify Vespula or Polistes sensitization. Allergy 67, 528–536 (2012).
Ollert, M. & Blank, S. Anaphylaxis to Insect Venom Allergens: Role of Molecular Diagnostics. Curr Allergy Asthma Rep 15, 527 (2015).
Sanchez, F. et al. Comparative study between European and American species of Polistes using sera from European sensitized subjects. Clin Exp Allergy 25, 281–287 (1995).
Schiener, M., Graessel, A., Ollert, M., Schmidt-Weber, C. B. & Blank, S. Allergen-specific immunotherapy of Hymenoptera venom allergy - also a matter of diagnosis. Hum Vaccin Immunother 13, 2467–2481 (2017).
Cervo, R., Zacchi, F. & Turillazzi, S. Polistes dominulus (Hymenoptera, Vespidae) invading North America: some hypotheses for its rapid spread. Insectes Sociaux 47, 155–157 (2000).
Eardley, C., Koch, F. & Wood, A. R. Polistes dominulus (Christ, 1791) (Hymenoptera: Polistinae: Vespidae) newly recorded from South Africa: short communication. African Entomology 17, 226–227 (2009).
Höcherl, N. & Tautz, J. Nesting behavior of the paper wasp Polistes dominula in CentralEurope - a flexible system for expanding into new areas. Ecosphere 6, 1–11 (2015).
Sturm, G. J. et al. EAACI Guidelines on Allergen Immunotherapy: Hymenoptera venom allergy. Allergy (2017).
Golden, D. B. Insect sting anaphylaxis. Immunol Allergy Clin North Am 27, 261–272, vii (2007).
Juarez, C. et al. Specific IgE antibodies to vespids in the course of immunotherapy with Vespula germanica administered to patients sensitized to Polistes dominulus. Allergy 47, 299–302 (1992).
Baker, T. W. et al. Stinging insect identification: Are the allergy specialists any better than their patients? Ann Allergy Asthma Immunol 116, 431–434 (2016).
Antolin-Amerigo, D. et al. Component-resolved diagnosis in hymenoptera allergy. Allergol Immunopathol (Madr) (2017).
Jakob, T., Muller, U., Helbling, A. & Spillner, E. Component resolved diagnostics for hymenoptera venom allergy. Curr Opin Allergy Clin Immunol 17, 363–372 (2017).
Frick, M. et al. rApi m 3 and rApi m 10 improve detection of honey bee sensitization in Hymenoptera venom-allergic patients with double sensitization to honey bee and yellow jacket venom. Allergy 70, 1665–1668 (2015).
Hofmann, S. C., Pfender, N., Weckesser, S., Huss-Marp, J. & Jakob, T. Added value of IgE detection to rApi m 1 and rVes v 5 in patients with Hymenoptera venom allergy. J Allergy Clin Immunol 127, 265–267 (2011).
Kohler, J. et al. Component resolution reveals additional major allergens in patients with honeybee venom allergy. J Allergy Clin Immunol 133, 1383–1389, 1389 e1381–1386 (2014).
Korosec, P. et al. High sensitivity of CAP-FEIA rVes v 5 and rVes v 1 for diagnosis of Vespula venom allergy. J Allergy Clin Immunol 129, 1406–1408 (2012).
Matricardi, P. M. et al. EAACI Molecular Allergology User’s Guide. Pediatr Allergy Immunol 27(Suppl 23), 1–250 (2016).
Mittermann, I. et al. Recombinant allergen-based IgE testing to distinguish bee and wasp allergy. J Allergy Clin Immunol 125(1300–1307), e1303 (2010).
Muller, U., Schmid-Grendelmeier, P., Hausmann, O. & Helbling, A. IgE to recombinant allergens Api m 1, Ves v 1, and Ves v 5 distinguish double sensitization from crossreaction in venom allergy. Allergy 67, 1069–1073 (2012).
Moawad, T. I., Hoffman, D. R. & Zalat, S. Isolation, cloning and characterization of Polistes dominulus venom phospholipase A1 and its isoforms. Acta Biol Hung 56, 261–274 (2005).
Hoffman, D. R., Severino, M. G., Campi, P., Turillazzi, S. & Zerboni, R. Protease is an important allergen in European Polistes wasp venom allergy. J Allergy Clin Immunol 101, S33 (1998).
Pantera, B. et al. Characterization of the major allergens purified from the venom of the paper wasp Polistes gallicus. Biochim Biophys Acta 1623, 72–81 (2003).
Blank, S. et al. Polistes species venom is devoid of carbohydrate-based cross-reactivity and allows interference-free diagnostics. J Allergy Clin Immunol 131, 1239–1242 (2013).
Caruso, B. et al. Evaluation of the IgE cross-reactions among vespid venoms. A possible approach for the choice of immunotherapy. Allergy 62, 561–564 (2007).
Schiener, M. et al. Application of recombinant antigen 5 allergens from seven allergy-relevant Hymenoptera species in diagnostics. Allergy 72, 98–108 (2017).
Blank, S. et al. Identification, recombinant expression, and characterization of the 100 kDa high molecular weight hymenoptera venom allergens Api m 5 and Ves v 3. J Immunol 184, 5403–5413 (2010).
Standage, D. S. et al. Genome, transcriptome and methylome sequencing of a primitively eusocial wasp reveal a greatly reduced DNA methylation system in a social insect. Mol Ecol 25, 1769–1784 (2016).
Chauhan, J. S., Rao, A. & Raghava, G. P. In silico platform for prediction of N-, O- and C-glycosites in eukaryotic protein sequences. PLoS One 8, e67008 (2013).
Aertgeerts, K. et al. Crystal structure of human dipeptidyl peptidase IV in complex with a decapeptide reveals details on substrate specificity and tetrahedral intermediate formation. Protein Sci 13, 412–421 (2004).
Kreil, G., Haiml, L. & Suchanek, G. Stepwise cleavage of the pro part of promelittin by dipeptidylpeptidase IV. Evidence for a new type of precursor-product conversion. Eur J Biochem 111, 49–58 (1980).
Radauer, C. et al. Update of the WHO/IUIS Allergen Nomenclature Database based on analysis of allergen sequences. Allergy 69, 413–419 (2014).
Blank, S. et al. Evaluation of different glycoforms of honeybee venom major allergen phospholipase A2 (Api m 1) produced in insect cells. Protein Pept Lett 18, 415–422 (2011).
Blank, S. et al. Api m 10, a genuine A. mellifera venom allergen, is clinically relevant but underrepresented in therapeutic extracts. Allergy 66, 1322–1329 (2011).
Seismann, H. et al. Dissecting cross-reactivity in hymenoptera venom allergy by circumvention of alpha-1,3-core fucosylation. Mol Immunol 47, 799–808 (2010).
Eberlein, B., Krischan, L., Darsow, U., Ollert, M. & Ring, J. Double positivity to bee and wasp venom: improved diagnostic procedure by recombinant allergen-based IgE testing and basophil activation test including data about cross-reactive carbohydrate determinants. J Allergy Clin Immunol 130, 155–161 (2012).
Frick, M. et al. Predominant Api m 10 sensitization as risk factor for treatment failure in honey bee venom immunotherapy. J Allergy Clin Immunol 138(1663–1671), e1669 (2016).
Blank, S. et al. Component-resolved evaluation of the content of major allergens in therapeutic extracts for specific immunotherapy of honeybee venom allergy. Hum Vaccin Immunother 13, 2482–2489 (2017).
Hemmer, W. [Cross reactions between Hymenoptera venoms from different families, genera and species]. Hautarzt 65, 775–779 (2014).
Jin, C. et al. Reassessing the role of hyaluronidase in yellow jacket venom allergy. J Allergy Clin Immunol 125(184–190), e181 (2010).
Grant, J. A. et al. Diagnosis of Polistes wasp hypersensitivity. J Allergy Clin Immunol 72, 399–406 (1983).
Aird, S. D. Snake venom dipeptidyl peptidase IV: taxonomic distribution and quantitative variation. Comp Biochem Physiol B Biochem Mol Biol 150, 222–228 (2008).
Position paper: Allergen standardization and skin tests. The European Academy of Allergology and Clinical Immunology. Allergy 48, 48–82 (1993).
Ring, J. & Messmer, K. Incidence and severity of anaphylactoid reactions to colloid volume substitutes. Lancet 1, 466–469 (1977).
Acknowledgements
We gratefully acknowledge the technical assistance of Stefanie Etzold, Franziska Martin, Kyra Swiontek and Stephanie Kler.
Author information
Authors and Affiliations
Contributions
M.S. planned and performed the experiments, analysed the data and wrote the manuscript. B.E. coordinated the recruitment of hymenoptera venom-allergic patients and supervised basophil activation tests. M.P. performed basophil activation tests and collected data. C.H., A.K., D.R. and S.P. performed M.S. experiments, analysed the data and revised the manuscript. G.P. recruited HBV- and YJV-allergic patients and collected data. P.S. recruited PDV-allergic patients and collected data. C.M.-A. coordinated the recruitment of PDV-allergic patients and collected and analysed the data. FdlR recruited PDV- and YJV-allergic patients and collected data. T.B. contributed to the interpretation of data and revised the final version of the manuscript. U.D. was responsible for the diagnostic work-up of hymenoptera venom- allergic patients. C.S.-W. supervised the study, contributed to the interpretation of data, and revised the final version of the manuscript. M.O. supervised the study, analysed the data, and wrote the manuscript. S.B. initiated and supervised the study, analysed the data and wrote the manuscript.
Corresponding author
Ethics declarations
Competing Interests
MS has received travel support from ALK-Abelló and Bencard. BE has received research funding from Bühlmann Laboratories. UD has been speaker, investigator and /or been a member of advisory boards for Allergopharma, ALK-Abelló, Bencard, GSK, Hermal, MEDA, Novartis Pharma, Stallergenes, Stiefel. TB has received research funding, speaker’s honorarium and consultancy fees from Thermo Fisher Scientific, has received research support from DFG, Novartis and Thermo Fisher Scientific, has received lecture fees from MSD, Novartis, HIPP GmbH & Co, ALK-Abelló, MedComms Ltd and Astellas Pharma GmbH. CBS-W has received grants from Allergopharma, Leti, PLS-Design and Regeneron; is member of the scientific advisory board of Leti and Bencard; has received consultancy fees from Leti, GLG Consultancy, Allergopharma and Bencard. MO has received consultancy fees from Siemens Healthcare, Hitachi Chemical Diagnostics and Bencard; has received lecture fees from Thermo Fisher Scientific, Bencard and Siemens Healthcare; is co-founder of PLS-Design GmbH. SB has received speaker’s honorarium and/or travel support from ALK-Abelló, Bencard and Thermo Fisher Scientific; has received consultancy fees as an advisory board member and research support from Bencard and Allergy Therapeutics. The other authors declare that they have no conflict of interest.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schiener, M., Hilger, C., Eberlein, B. et al. The high molecular weight dipeptidyl peptidase IV Pol d 3 is a major allergen of Polistes dominula venom. Sci Rep 8, 1318 (2018). https://doi.org/10.1038/s41598-018-19666-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-19666-7
This article is cited by
-
An integrated transcriptomic and proteomic approach to identify the main Torymus sinensis venom components
Scientific Reports (2021)
-
Marker allergens in Hymenoptera venom allergy — Characteristics and potential use in precision medicine
Allergo Journal International (2021)
-
Protein changes as robust signatures of fish chronic stress: a proteomics approach to fish welfare research
BMC Genomics (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
