
Abstract
The commonality of facial expressions of emotion has been studied in different species since Darwin, with most of the research focusing on closely related primate species. However, it is unclear to what extent there exists common facial expression in species more phylogenetically distant, but sharing a need for common interspecific emotional understanding. Here we used the objective, anatomically-based tools, FACS and DogFACS (Facial Action Coding Systems), to quantify and compare human and domestic dog facial expressions in response to emotionally-competent stimuli associated with different categories of emotional arousal. We sought to answer two questions: Firstly, do dogs display specific discriminatory facial movements in response to different categories of emotional stimuli? Secondly, do dogs display similar facial movements to humans when reacting in emotionally comparable contexts? We found that dogs displayed distinctive facial actions depending on the category of stimuli. However, dogs produced different facial movements to humans in comparable states of emotional arousal. These results refute the commonality of emotional expression across mammals, since dogs do not display human-like facial expressions. Given the unique interspecific relationship between dogs and humans, two highly social but evolutionarily distant species sharing a common environment, these findings give new insight into the origin of emotion expression.
Similar content being viewed by others


Introduction
The common origin of emotions has long been a subject of scientific interest1 with different emotional responses producing a diverse range of communicative elements, especially through the face. Facial expressions are also correlates of internal state in both humans2 and other animals3,4,5,6 and so may be used, in part, to infer emotion alongside other component processes, such as physiological activation and behavioural tendencies7. Many studies (e.g.8,9) use an holistic approach (i.e. categorizing the whole face as angry, happy, etc.) to classify the target facial expressions, which reflects the way the human brain processes faces7,10, but can be problematic when examining the underlying mechanism of emotion perception across species. For instance, there is a diverse range of smiling faces with different visual characteristics and different emotional meanings in humans11,12. As a classic example, the Duchenne smile (felt, true enjoyment) differs by one muscle contraction from the non-Duchenne smile (unfelt, usually produced in formal greetings). Moreover, during laughter and depending on the context, a blend of both Duchenne and non-Duchenne smiles is often observed13. Hence, simply classifying a facial expression as “happy” is too simplistic and less meaningful for cross-species comparison. Furthermore, the same ‘holistic’ facial morphological configuration could have different functional meanings (i.e. result in distinctly different behavioural consequences) depending on the species14,15. For example, the Play Face (PF) and the Full Play Face (FPF) are variants of the same facial expression, where the former presents an open mouth with lower teeth exposed, and the latter incorporates visible upper teeth. Both the PF and the FPF represent different degrees of playful expression in great apes (humans included)16,17,18. Conversely, in crested macaques, mandrills and geladas, the FPF is not just a more intense version of the PF, but instead is derived from convergence between the PF and the silent-bared teeth display (SBT), a facial expression observed in affiliative settings such as grooming19. Additionally, the SBT indicates submission and appeasement in Barbary macaques14, signals affinity and benign intentions in humans, and, in chimpanzees, is present in a range of situations from response to aggression to affinity contexts15.
As an alternative to an holistic descriptive approach, the decomposition and objective description of distinct anatomical regions of facial features, such as occurs with the Facial Action Coding System (FACS20), has been the golden standard to study human facial expressions of emotion across individuals of different races and cultures for several decades21,22. Each of the discrete facial movements identified (Action Units, AUs) is the result of an independent facial muscle contraction that can produce several changes in appearance to the face, which in turn are used to identify which AUs are activated. Thus, FACS codes facial movements from a purely anatomical basis, avoiding circular reasoning or a priori assumptions of emotion meaning. Recently, FACS has been adapted to several non-human species23,24,25,26,27,28,29,30, such as chimpanzees and orangutans, following the original methodology20 and has proven to be a successful tool for objectively investigating and comparing facial expressions of closely related species31,32. For example, chimpanzees and humans share an identical facial muscle plan (differing by only one muscle)20,33, but chimpanzees display both homologous (e.g. play face and human laugh) and species-specific expressions (e.g. pant-hoot)32,34.
While the human prototypical facial expressions of emotion are well established, little is known about the quantitative and empirical nature of the emotional facial displays of the domestic dog, an evolutionarily remote, but socially complex species which often shares the human social environment and frequently engages in interspecific communication with an emotional content (e.g.35). To date, functional facial expressions in dogs have been largely discussed holistically in relation to their approach-avoidance value, for example, the “threat gape” in fight-flight situations3, and the PF or the Relaxed Open Mouth (ROM) as a social communicative signal for play solicitation and within play bouts3,36,37. With the development of the FACS for the domestic dog24, it becomes possible to apply a bottom-up technique to investigate the composition and meaning of dogs’ facial expressions and, more importantly, to establish possible analogies with humans, with whom they socially interact.
Dogs and humans, like other mammals, have a homologous facial anatomy plan38,39 even though they belong to phylogenetically distant groups. Additionally, both species share a common mammalian neuroanatomy for the basic emotions such as fear and happiness40,41,42,43,44, typically live in a common social and physical environment, are very facially expressive (e.g.2,3,20,24), and respond to the same or similar conspecific and heterospecific social cues (e.g.45). Consequently, the facial cues and expression of emotion in home-dwelling pet dogs provide a unique comparative model for the study of phylogenetic inertia (i.e. absence of expected change and/or adaptation to an optimal state given specific selection pressures in the current environment)46,47,48 versus evolutionary divergence (i.e. a set of changes brought about by selection pressures from a common ancestor resulting in homologies)49,50 versus evolutionary convergence (i.e. a set of changes from selection pressures acting in independent lineages to create similarity in the resulting analogies)47,50.
Here, we investigated the mechanistic basis of facial expressions in humans and dogs, by objectively measuring their video recorded facial actions during immediate reactions to emotionally-competent stimuli. The FACS20 and the DogFACS24 were applied in a range of contexts associated with four categories of emotional responses: a) happiness, b) positive anticipation, c) fear, and d) frustration (Table 1). Instead of selecting the basic emotions that are known to produce universal facial signals in humans51, we focused on emotions that are defined by evolutionary and biologically consistent criteria: 1) essential for solving adaptive problems in mammals (e.g. fear of a threat prompts flight increasing survival)52, 2) arise from corresponding physiological markers (e.g. elevated opioid levels can increase playfulness53), and 3) correlate with specific neuroanatomical regions (e.g. Nucleus accumbens neurons activate before a positive event leading to positive anticipation44,54). This approach reduces anthropomorphic and anthropocentric bias in the selection and comparison of emotions, i.e. instead of trying to identify stereotypically human emotions in dogs, we focused on examining shared underlying mammalian homologies55. Furthermore, for each category of emotion (e.g. fear), we used a range of contexts to generate the emotional response (thunderstorms, specifically avoided objects, etc.). This increased the likelihood of identifying the general facial responses to the emotional category of stimulus (e.g. general facial actions of fear), instead of behavioural motivations (e.g. facial actions displayed for thunderstorms, but not in other fear contexts). We only analysed spontaneous emotional reactions because posed responses could differ from spontaneous ones in duration, intensity, symmetry and form56,57,58.
Given the common origin of mammalian facial musculature59 and its link to emotion2, and the nature of the long association between humans and dogs, it is plausible that similar emotional reactions might share common facial correlates in these two species (i.e. that the same emotions are closely associated with the same core facial muscle movements). Therefore, we tested two hypotheses: 1) Do dogs produce specific facial actions in response to different categories of emotionally-competent stimuli (i.e. stimuli that produce an emotion cascade resulting in a distinct brain response60,61)? If so, this would provide evidence that dogs possess an adaptive facial behavioural repertoire (sensu Schmidt and Cohn62) with expressive and/or communicative functions associated with the underlying emotional response, as a result of evolutionary pressures. This is a precondition for the main hypothesis: 2) Do dogs use similar facial muscles to humans to express similar emotions? A lack of significant differences between humans and dogs would potentially reflect phylogenetic inertia and be consistent with the shared basic origin of emotional expression as proposed by Darwin1 or reflect convergent evolution. On the other hand, widespread significant differences would indicate that facial expressions of emotion are not highly conserved features across different species.
Results
Human facial actions of emotion
Our study showed convergent validity with previous studies (Table 2) for the core AUs63 associated with each emotion (Table 1). Humans showed a differential use of their facial musculature to produce a higher rate (i.e. in comparison to the relaxed condition representing neutral facial expression) of specific prototypical facial actions during an emotional response, while using various AUs flexibly between contexts. The comparable core AUs identified in our study included AU5 and AU20 for fear, AU14, AU17 and AU24 for frustration, and AU6 for happiness, confirming our human videos as a suitable baseline to compare with the dog videos. This also allowed us to verify that the naturalistic videos used to extract our data can still produce robust results, with facial signals strongly linked to corresponding emotional responses. Additionally, the human FACS coding created a baseline to compare with the dogs’ responses, based on human-dog muscular homologies.
Specifically, our analysis showed that compared to the relaxed condition, humans displayed a significantly higher rate of specific facial actions, in response to stimuli associated with happiness, fear and frustration, but not positive anticipation. For the fear context, the rate of AU5, AU7, AU20 and AD38 were significant; for the frustration context, the rate of AU14, AU17, AU24, AU28 and AD84 were significant; for the happiness context, the rate of AU6 was significant. In the Supplementary Table S4, the facial actions that humans displayed during two or more emotions, but not during the relaxed context are also reported.
Dog facial actions of emotion
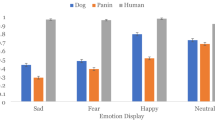
In support of our first hypothesis, we found significant differences between each emotional response and relaxation for particular facial and ear actions: Dogs consistently showed a higher rate of AD126 during fear contexts; AD37, AD137 and EAD102 during positive anticipation; and AU27 during happiness. However, frustrated dogs did not display higher rates of any of the facial actions (Fig. 1, Table 3). The higher rates of these specific facial actions are thus strongly linked to the respective emotional contexts. In the Supplementary Table S4, we report the display of significantly higher rates of those facial actions common between emotions (but absent during relaxation); like humans, dogs made a clear, flexible use of their facial musculature.

Examples of visual representations of unique dog facial actions displayed during emotional reactions. (Individual images composed for illustration of muscular movements only, found in Pixabay 2016, free for commercial use, https://pixabay.com/en/herder-action-dog-animal-dog-plays-1726406/, https://pixabay.com/en/dogs-cute-pet-animal-canine-1181868/, https://pixabay.com/en/animal-canine-cute-dog-friendly-1837003/).
Comparison of human and dog facial actions of emotion
To investigate our second and main hypothesis, direct comparison between human and dog facial actions for each emotion revealed significant differences in specific actions for all the examined emotions (Table 4), demonstrating the differential use of facial musculature between humans and dogs when facing equivalent emotional situations. When compared with humans, dogs displayed higher rates of AD19 in a fearful situation, AU45 during frustration, AD19 and AD37 during positive anticipation, AD19 during happiness, and AU1 in relaxed situations. We also report in Table 4 the facial movements that were significantly higher for humans compared with dogs in the same context. These results show that, in equivalent contexts, dogs and humans mostly activate different facial actions. Out of all the facial actions entered in the analysis, only two Action Units (AU25, AU26) and two Action Descriptors (AD33 and AD40) showed no significant differences between the two species in any of the contexts. This indicates that the majority of facial actions which both dogs and humans are able to produce were used differently for all emotional contexts. Effect sizes were mostly intermediate to large and are reported in the Supplementary Table S5.
Control variables
We found no significant differences between different categories of cephalic type, ear morphology and breed for any of the significant facial actions in our main analysis of dogs, i.e. these variables had no significant impact on the production of facial actions in the emotional contexts analysed in this study. For jowl length, the rate of AU10 during happiness was indistinguishable in dogs with short and long jowls, but was higher than in dogs with no jowls (Kruskal-Wallis, with Dunn-Bonferroni correction, H2 = 6.736, p = 0.050); furthermore AU12 had a significantly higher rate in dogs with short jowls than in dogs with long jowls (Kruskal-Wallis, with Dunn-Bonferroni correction, H2 = 9.889, p = 0.036). However, this observation might be a coding artefact as long or no jowls make these AU movements less conspicuous and thus harder to recognise. Finally, we found significantly higher levels of arousal for all emotions when compared with the relaxed referent (Kruskal-Wallis, with Dunn-Bonferroni correction, Fear: H4 = 45.20, p = 0.0001, frustration: H4 = 69.650, p = 0.0001, positive anticipation: H4 = 69.857, p = 0.0001, happiness: H4 = 96.500, p = 0.0001) and for happiness when compared with fear (Kruskal-Wallis, with Dunn-Bonferroni correction, H4 = 51.300, p = 0.0001). This supports the validity of the relaxed context as a point of reference, since this represents, by definition, absence of/very low arousal.
Discussion
This study provides the first empirical evidence to address two important research questions in comparative emotion and interspecific emotion understanding: dogs display specific discriminatory facial movements in response to different emotional stimuli, but do not display similar facial movements to humans when reacting to emotionally comparable contexts.
Dogs tended to make more frequent use of AD126 in a fearful context; AD37, AD137 and EAD102 during positive anticipation; and AU27 in a happiness context (Fig. 1). Such observation is in agreement with a widely held belief that dogs facially react in a common way as a species when confronted with particular challenges or situations in their environment (e.g. when fearful64,65). This also supports Darwin’s suggestions that facial expressions are correlates of internal states in animals1, and are based on homologous facial musculature with the potential to produce similar facial information for a given species. Given previous studies mentioning varied facial cues in frustrated dogs (also described as displacement or conflict behaviours35,66), it was surprising that we did not observe distinctive facial actions for frustration in dogs. However, it might be that the frustration signals previously reported are not specific to frustration and are instead used flexibly in association with other behaviours and/or in response to different contexts (e.g. stress67, discomfort68, fear65), and/or that facial expressions of frustration are more context specific, without a common dynamic anatomical structure. Analysing more videos or featuring a wider range of contexts, to account for potential flexibility, might identify specific facial actions for this emotion in certain common contexts.
Regarding our second hypothesis, our results revealed that humans and dogs produced distinct facial expressions of emotion and little commonality was found in the facial actions triggered, i.e., dogs did not show human-like facial expressions. This observation refutes the idea of continuity of emotional expression between different species. Given the clear facial muscular homology and comparable anatomical facial plan between humans and dogs38,39, their shared social environment and long history of intensive mutual social interaction69,70, it might have been expected that dogs and humans would at least share some facial responses within a comparable emotion context.
Most of the basic facial muscles are similar between humans and dogs, and share primary functions unrelated to facial expression (e.g. the orbicularis oculi muscle that closes the eye to protect the eye or the zygomaticus major that draws the lips back to facilitate feeding). However, when it comes to emotional expression, dogs do not seem to make use of their muscles in the same way that humans do. This might be due to the inability of the dog’s muscles to produce particular movements present in humans because of their different facial morphology (e.g. lack of localised fat deposits). This is the case, for example, with AU6 (produced by the orbicularis oculi muscle), which is a fundamental AU present in all Duchenne happy faces in humans, but in dogs it was never observed even though the same muscle is present and functional. The human face has a more complex facial musculature, i.e. a few muscles are not present in dogs, such as the levator palpebrae that produces AU5 or the risorius that produces AU20. Both AU5 and AU20 (i.e. eyes widely open, lips stretched laterally and downwards) are critical facial actions of a fearful face in humans, but it is impossible for dogs to produce the equivalent facial change.
Another example of muscular variance that might reflect the reported difference in the displayed facial actions relates to the human-dog frustration comparison. Dogs lack a zygomaticus minor muscle, which produces AU14 in humans. This specific movement is one of the core action units of human frustration facial expression. Given the lack of these specific muscles in dogs, it is possible that other muscles could be activated to produce reliable fear or frustration cues in this species. This appears to be the case for fear, since AD126 production had unique characteristics in this context. However, we did not identify an equivalent in the case of frustration, as discussed earlier.
Given the low number of specific facial actions produced in association with each emotion, we suggest that dogs do not display a composed facial expression with several facial actions being integrated in a stereotypical display, as is observed in humans. Instead, dogs seem to produce isolated actions in response to specific emotionally-competent stimuli.
Due to well-known human perception biases (e.g. anthropomorphic tendencies71), it is essential to determine in an objective and standardized way exactly how dogs produce emotional cues. The results in relation to our second hypothesis illustrate the error of broad descriptions and over-reliance on holistic categorization of facial behaviour, which can lead to biases in the perception of facial expressions of a different species (e.g.14,15). It has been argued that an alternative explanation for why humans perceive non-human animals’ emotions as if conspecifics is because of evolutionary homologies between the species, based on shared mammalian brain structures and behavioural expression patterns42,71. However, our study refutes this hypothesis, as the homologies between humans and dogs seem to be limited to the underlying musculature, rather than their activation. In other domestic animals, such as horses72 and sheep73, the basic facial musculature also appears to be well conserved26,74, but the emotional displays appear to diverge from what is observed in humans. It is worth noting that in both of these species (and arguably most domestic animals) the ears appear to be widely used as part of emotional signals, while in humans the ears have more vestigial muscles. Close analysis of the aural signals may therefore be a fruitful line of enquiry in the future. In this respect, it is also worth noting that greater similarities are seen between different species of primates’ (including human) facial expressions75, with them showing a continuity not only at a muscular level but also at the level of expression production. In the case of the dog-human comparison, and unlike the chimpanzee-human comparison, facial expressions seem to be exclusive to the species. Phylogenetic inertia was likely involved in the development of the well-conserved facial musculature of dogs and humans47, but there are clearly differences in the way this musculature is used to produce facial expressions associated with the intentional or unintentional communication of emotion.
Our findings of humans and dogs displaying distinctively different facial expressions have important theoretical and practical implications on human-dog emotion understanding and its underlying cognitive mechanisms. There is little theoretical understanding of interspecific emotional appraisal, but we examine the extension of two competing theories of human-human emotion perception into the field of human-dog emotion perception: “The mental simulation strategy” (i.e. if the same facial movements are produced in response to the same emotional stimuli in both species, individuals could form an understanding of the emotions of others through their own facial actions), and “The implicit social learning strategy” (i.e. if facial movements produced are different between species, an associative learning between external events and motivational states is likely needed for understanding the emotions of other species)76,77. Given that in our study facial movement production differed between humans and dogs, it is unlikely that a mental simulation strategy could be useful or adopted by both species when interpreting heterospecific expressive facial signals. Instead, an implicit social learning strategy would be a more meaningful way to establish a functional human-dog emotion understanding. This would at least partly explain why untrained humans do not seem proficient in reading dogs’ facial and body language78,79,80, particularly subtle cues such as head turning or nose licking81. This is further supported by the neurocognitive evidence that people read dogs’ and humans’ social cues using overlapping brain areas82,83 and similar cognitive mechanisms76,84. Indeed, humans represent non-human animals’ affective space similarly to that of conspecifics’71,85 and incorrectly identify emotions in dogs that have been shown to be a direct result of anthropomorphic subjective judgements (e.g. guilt86). In our study, most emotional facial actions produced by dogs were in the mouth area, using the tongue or the opening of the mouth, and none were in the eye area, which is an important difference from humans that produce eye region AUs in many of the prototypical facial expressions. Thus this might be another reason why humans seem to find it very hard to read dogs’ communication.
The preceding studies strongly suggest that humans read dog communication as they would read other humans (own-species bias87), and our results indicate this is potentially problematic when it comes to evaluation of the emotional content of these signals. This leads to another important issue: all studies to date on human perception of dogs are missing bias-free empirical validation (e.g.88,89,90). For example, when showing a “happy dog” static picture, the only “validation” is the agreement of other subjective human judges, without ever specifying why a “happy dog” is identified as happy, leading to subjective anthropomorphic assessments and circular reasoning, i.e. different human dog “experts” agree with each other on what a “happy dog” should look like, instead of examining what does a “happy dog” actually look like. For the first time, our study demonstrates how to validate dogs’ facial expressions by applying an anatomical and bias-free method based on the presence of empirical stimuli of relevance.
It remains to be seen to what extent the current findings can be generalised across all dog breeds and other emotional contexts, given the variation in facial morphology and possibly muscular functionality in different breeds. Nonetheless, we did establish that features like variation in general head shape may be less important than changes in jowl structure, given the location of key AUs used to express the emotions studied. Despite this study’s focus on facial expressions alone, it is important to mention that emotional expression is clearly multimodal in many species including those studied here (e.g. vocalisations, body posture, and odours may be important moderators of the message sent). While the stimuli were selected for their fundamental emotional equivalence between the two species, the resulting expression might have been moderated by other factors associated with their processing, which were not controlled for (e.g. social vs non-social stimulus source91). Nevertheless, the critical feature is that they should largely share a final common emotional process, whose expression we set out to measure. Thus, future studies might aim to compare dogs of different breeds and morphologies, as well as incorporate the multimodality of emotional signals, and test these hypotheses in different settings (e.g. the more controlled laboratory) to expand on these conclusions.
Methods
Video source and individuals
The videos used in this study were selected from online databases (www.youtube.com and AM-FED database92). Only one individual per video was coded. Whenever there was more than one individual present in the same video that fulfilled all the necessary criteria (see section b), one of the individuals was randomly selected and coded.
The sample consisted of 150 individuals in total, 50 humans and 100 family dogs, distributed equally between emotional contexts (happiness, positive anticipation, fear, frustration and relaxation, Table 1). For detailed information on individuals, please see Supplementary material S1.
Criteria for video selection
The four categories of emotion were defined according to literature sources (Table 1). As a control, we selected videos of neutral/relaxed individuals, i.e., where any specific emotionally-triggering stimuli or overt behavioural reactions were absent. The videos were chosen on the basis of stimulus quality (e.g. source) and its clear association with an evoked response, with different stimuli selected for humans and dogs to allow functional equivalence93,94 (e.g., most humans are not afraid of thunderstorms, while this is a common fear trigger in dogs95,96,97). Videos of dog training, clinical/veterinary settings or with music covering the audio were excluded. Only videos with minimum editing, high image quality (at least 720p), good lighting and visibility of faces were selected. The putative emotion eliciting stimulus had to be apparent and clearly identifiable for at least part of the video. By including homemade/amateur videos with naturalistic and spontaneous behaviour we ensured that the responses were ecologically valid, less constricted, and more robust, especially when compared to laboratory studies on emotions98.
To ensure that the emotionally-competent stimuli were present in the dog videos and were impacting their behaviour, the first author of this study (CC) selected, blinded and randomised the video order. Another author (DM, specialist in veterinary behavioural medicine) relabelled all videos according to the emotion likely triggered by the stimulus99. Only videos that had full agreement were included in the study.
FACS, control variables and reliability coding
All human and dog videos were coded with FACS20 and DogFACS24, respectively, by a certified coder (CC) via the open source software BORIS (Behavioural Observation Research Interactive Software) V2.96100. Each video duration was determined by the individual’s emotional response to the trigger, i.e. from the starting of the behavioural display to the ending time point (neutral behaviour). For example, for the fear trigger of a thunderstorm, the trigger was present throughout the video, for all thunderstorm videos analysed, and the duration of the video was considered while the response (flight/freeze, etc.) of the individual was observed. Before starting the FACS coding, one or more neutral face frames were selected for all individuals. The number of facial actions was identified by watching the videos frame-by-frame, logged in BORIS and extracted for analysis. During speech/vocalizations, facial actions were coded only if they were above level A intensity (as per FACS investigator guide recommendation101). To ensure an unbiased coding, an independent DogFACS certified coder, unaware of the aim of the study or the emotional categorization of the videos, re-coded a random 30% of the dog videos. Inter-coder reliability was thus obtained with 84% mean agreement in the Intraclass Correlation Coefficient (ICC(3,k), CI range of 0.80–0.87).
Since morphological differences could potentially impact upon the production of facial actions in dogs, we used cephalic type (brachycephalic, mesaticephalic, dolichocephalic), jowls length (none, short, long), ear morphology (floppy, partially erect, erect) and breed (as per the British Kennel Club) as control variables. We only analysed the effects of these control variables in the facial actions that were significant in our main analysis. As an additional control, we categorized all the videos for the perceived level of arousal (five point Likert scale: 1 – very low, 2 – low, 3 – moderate, 4 – high, 5 – very high) to ensure arousal was not a confounding variable for emotion. These control variables allowed us to increase the internal validity of our study, since it excludes alternative explanations for the causal relationship between independent and dependent variables102. Different blinded coders recoded the whole sample for each of the control variables to assess mean agreement of inter-rater reliability (ICC(3,k)), obtaining 94% (CI range of 0.91–096) for ears, 85% (CI range of 0.78–0.90), for jowls, and 87% (CI range of 0.81–0.90) for arousal. Human videos were not coded for control or reliability purposes, instead, our results were compared with the respective facial movements reported in the established literature.
Statistical analysis
Because the videos varied in duration (62 ± 45 s, mean ± s.d.) due to differences in the individuals’ responses to the triggers and the emotions examined, the number of facial actions (4.59 ± 8.97, mean ± s.d.) was normalised into rates of visible time, i.e., the number of each AU was divided by the duration of the video where the face is visible. Variables with very low rates (<0.001) were excluded from analysis (Supplementary material S3). Thus, we included 30 human facial actions and 22 dog facial actions in the statistical analysis (Supplementary Table S2). We performed Kolmogorov–Smirnov tests for normality and non-parametric Levene’s test for homoscedasticity. Since all variables violated both assumptions in at least one of the groups, non-parametric tests were used throughout the analysis. To investigate if dogs produced differential facial actions in response to specific emotional triggers, we compared the facial actions displayed between emotions and between each emotion and the control relaxation videos with Kruskal-Wallis (adjusted for ties) followed by post-hoc pairwise multiple comparisons tests, with Dunn-Bonferroni correction. For our second hypothesis, where we were interested in assessing if dogs were using the same facial actions as humans in response to an equivalent emotional trigger, we performed Mann-Whitney tests, with exact p-value for the homologous facial actions between species. For both hypotheses, the rate of facial movements was used as the dependent variable and the emotion category was the independent variable. Additionally, we calculated effect sizes to consider and minimize the risk of type 2 error in our interpretation of the results.
To analyse the potential effect of the control variables cephalic type, ear morphology, jowl length, breed, and arousal level on the facial actions, we used Kruskal-Wallis tests (adjusted for ties) and post-hoc pairwise multiple comparisons tests, with Dunn-Bonferroni correction. Here, we compared the rate of each facial action (dependent variable) between the categories (independent variables) of the control variables, in order to ensure the distribution of facial actions was the same regardless of the dogs’ morphological variation. For ear morphology and jowl length, we only performed the analysis for the relevant facial actions (e.g. jowl length only analysed for lower face actions). All statistical analyses were performed using IBM SPSS v22, except for the effect sizes that were computed with GPower v3.1.
Ethics
This study was approved by the School of Psychology Research Ethics Committee and follows the Ethics Guidelines for Internet-mediated Research by the British Psychological Society.
Data availability
The datasets generated and analysed during the current study are available from the corresponding author on request.
Change history
05 July 2018
A correction to this article has been published and is linked from the HTML and PDF versions of this paper. The error has been fixed in the paper.
References
Darwin, C. The expression of the emotions in man and animals. (D. Appleton and Company, 1896).
Ekman, P. & Oster, H. Facial expressions of emotion. Annu. Rev. Psychol. 30, 527–554 (1979).
Fox, M. W. A comparative study of the development of facial expressions in canids: wolf, coyote and foxes. Behaviour 36, 49–73 (1970).
Chevalier-Skolnikoff, S. Facial expression of emotion in nonhuman primates. In Darwin and Facial Expression: A Century of Research In Review (ed. Ekman, P.) (Academic Press, 1973).
Tate, A. J., Fischer, H., Leigh, A. E. & Kendrick, K. M. Behavioural and neurophysiological evidence for face identity and face emotion processing in animals. Philos. Trans. R. Soc. B Biol. Sci. 361, 2155–2172 (2006).
Leopold, D. A. & Rhodes, G. A comparative view of face perception. J. Comp. Psychol. 124, 233–251 (2010).
Scherer, K. R. What are emotions? And how can they be measured? Soc. Sci. Inf. 44, 695–729 (2005).
Dimberg, U. & Petterson, M. Facial reactions to happy and angry facial expressions: Evidence for right hemisphere dominance. Psychophysiology 37, 693–696 (2000).
Surguladze, S. A. et al. Recognition accuracy and response bias to happy and sad facial expressions in patients with major depression. Neuropsychology 18, 212–218 (2004).
Etcoff, N. L. & Magee, J. J. Categorical perception of facial expressions. Cognition 44, 227–240 (1992).
Ekman, P. & Friesen, W. V. Felt, false, and miserable smiles. J. Nonverbal Behav. 6, 238–252 (1982).
Gervais, M. & Wilson, D. S. The evolution and functions of laughter and humor: A synthetic approach. Q. Rev. Biol. 80, 395–430 (2005).
Harris, C. & Alvarado, N. Facial expressions, smile types, and self-report during humour, tickle, and pain. Cogn. Emot. 19, 655–669 (2005).
Preuschoft, S. “Laughter” and “Smile” in Barbary Macaques (Macaca sylvanus). Ethology 91, 220–236 (1992).
Waller, B. M. & Dunbar, R. I. M. Differential behavioural effects of silent bared teeth display and relaxed open mouth display in chimpanzees (Pan troglodytes). Ethology 111, 129–142 (2005).
Waller, B. M. & Cherry, L. Facilitating play through communication: Significance of teeth exposure in the gorilla play face. Am. J. Primatol. 74, 157–164 (2012).
Palagi, E. Social play in bonobos (Pan paniscus) and chimpanzees (Pan troglodytes): Implications for natural social systems and interindividual relationships. Am. J. Phys. Anthropol. 129, 418–426 (2006).
Palagi, E., Antonacci, D. & Cordoni, G. Fine-tuning of social play in juvenile lowland gorillas (gorilla gorilla gorilla). Dev. Psychobiol. 49, 433–445 (2007).
Palagi, E. & Mancini, G. Playing with the face: Playful facial “chattering” and signal modulation in a monkey species (Theropithecus gelada). J. Comp. Psychol. 125, 11–21 (2011).
Ekman, P., Friesen, W. & Hager, J. Facial Action Coding System (FACS): manual. (Research Nexus, 2002).
Ekman, P. & Friesen, W. V. A new pan-cultural facial expression of emotion. Motiv. Emot. 10, 159–168 (1986).
Ekman, P. & Rosenberg, E. L. What the face reveals: Basic and applied studies of spontaneous expression using the Facial Action Coding System (FACS). (Oxford University Press, 1997).
Caeiro, C. C., Waller, B. M., Zimmermann, E., Burrows, A. M. & Davila-Ross, M. OrangFACS: A muscle-based facial movement coding system for orangutans (Pongo spp.). Int. J. Primatol. 34, 115–129 (2013).
Waller, B. M. et al. Paedomorphic facial expressions give dogs a selective advantage. PLOS ONE 8, e82686 (2013).
Caeiro, C. C., Burrows, A. & Waller, B. M. Development and application of CatFACS: Are human cat adopters influenced by cat facial expressions? Appl. Anim. Behav. Sci. 189, 66–78 (2017).
Wathan, J., Burrows, A. M., Waller, B. M. & McComb, K. EquiFACS: The Equine Facial Action Coding System. PLOS ONE 10, e0131738 (2015).
Waller, B. M., Lembeck, M., Kuchenbuch, P., Burrows, A. M. & Liebal, K. GibbonFACS: A muscle-based facial movement coding system for hylobatids. Int. J. Primatol. 33, 809–821 (2012).
Parr, L. A., Waller, B. M., Burrows, A. M., Gothard, K. M. & Vick, S. J. Brief communication: MaqFACS: A muscle-based facial movement coding system for the rhesus macaque. Am. J. Phys. Anthropol. 143, 625–630 (2010).
Julle-Danière, É. et al. MaqFACS (Macaque Facial Action Coding System) can be used to document facial movements in Barbary macaques (Macaca sylvanus). PeerJ 3, e1248 (2015).
Vick, S. J., Waller, B. M., Parr, L. A., Pasqualini, M. C. S. & Bard, K. A. A cross-species comparison of facial morphology and movement in humans and chimpanzees using the Facial Action Coding System (FACS). J. Nonverbal Behav. 31, 1–20 (2007).
Waller, B. M., Misch, A., Whitehouse, J. & Herrmann, E. Children, but not chimpanzees, have facial correlates of determination. Biol. Lett. 10, 20130974–20130974 (2014).
Parr, L. A., Waller, B. M. & Vick, S. J. New developments in understanding emotional facial signals in chimpanzees. Curr. Dir. Psychol. Sci. 16, 117–122 (2007).
Burrows, A. M., Waller, B. M., Parr, L. A. & Bonar, C. J. Muscles of facial expression in the chimpanzee (Pan troglodytes): descriptive, comparative and phylogenetic contexts. J. Anat. 208, 153–167 (2006).
Parr, L. A., Waller, B. M., Vick, S. J. & Bard, K. A. Classifying chimpanzee facial expressions using muscle action. Emotion 7, 172–181 (2007).
Kuhne, F., Hößler, J. C. & Struwe, R. Emotions in dogs being petted by a familiar or unfamiliar person: Validating behavioural indicators of emotional states using heart rate variability. Appl. Anim. Behav. Sci. 161, 113–120 (2014).
Bekoff, M. Social play in coyotes, wolves, and dogs. BioScience 24, 225–230 (1974).
Cordoni, G., Nicotra, V. & Palagi, E. Unveiling the “secret” of play in dogs (Canis lupus familiaris): Asymmetry and signals. J. Comp. Psychol. 130, 278–287 (2016).
Bolwig, N. Facial expression in primates with remarks on a parallel development in certain carnivores (a preliminary report on work in progress). Behaviour 22, 167–192 (1964).
Diogo, R., Wood, B. A., Aziz, M. A. & Burrows, A. On the origin, homologies and evolution of primate facial muscles, with a particular focus on hominoids and a suggested unifying nomenclature for the facial muscles of the Mammalia. J. Anat. 215, 300–319 (2009).
Lang, P. J., Davis, M. & Öhman, A. Fear and anxiety: animal models and human cognitive psychophysiology. J. Affect. Disord. 61, 137–159 (2000).
Berridge, K. C. Comparing the emotional brains of humans and other animals. In Handbook of Affective Sciences (eds. Davidson, R. J., Scherer, K. R. & Goldsmith, H. H.) 25–51 (Oxford University Press, 2003).
Panksepp, J. Affective consciousness: Core emotional feelings in animals and humans. Conscious. Cogn. 14, 30–80 (2005).
Phelps, E. A. & LeDoux, J. E. Contributions of the amygdala to emotion processing: From animal models to human behavior. Neuron 48, 175–187 (2005).
Panksepp, J. The basic emotional circuits of mammalian brains: Do animals have affective lives? Neurosci. Biobehav. Rev. 35, 1791–1804 (2011).
Guo, K., Meints, K., Hall, C., Hall, S. & Mills, D. Left gaze bias in humans, rhesus monkeys and domestic dogs. Anim. Cogn. 12, 409–418 (2009).
Hansen, T. F., Pienaar, J. & Orzack, S. H. A comparative method for studying adaptation to a randomly evolving environment. Evolution 1965–1977, https://doi.org/10.1111/j.1558-5646.2008.00412.x (2008).
Shanahan, T. Phylogenetic inertia and Darwin’s higher law. Stud. Hist. Philos. Sci. Part C Stud. Hist. Philos. Biol. Biomed. Sci. 42, 60–68 (2011).
Blomberg, S. P. & Garland, T. Tempo and mode in evolution: phylogenetic inertia, adaptation and comparative methods. J. Evol. Biol. 15, 899–910 (2002).
Darwin, C. On the origin of species. (John Murray, 1859).
Hall, B. K. Descent with modification: the unity underlying homology and homoplasy as seen through an analysis of development and evolution. Biol. Rev. 78, 409–433 (2003).
Ekman, P. An argument for basic emotions. Cogn. Emot. 6, 169–200 (1992).
Panksepp, J., Fuchs, T. & Iacobucci, P. The basic neuroscience of emotional experiences in mammals: The case of subcortical FEAR circuitry and implications for clinical anxiety. Appl. Anim. Behav. Sci. 129, 1–17 (2011).
Trezza, V., Baarendse, P. J. J. & Vanderschuren, L. J. M. J. The pleasures of play: pharmacological insights into social reward mechanisms. Trends Pharmacol. Sci. 31, 463–469 (2010).
Berridge, K. C. Reward learning: Reinforcement, incentives, and expectations. Psychol. Learn. Motiv. 40, 223–278 (2000).
Charland, L. C. The natural kind status of emotion. Br. J. Philos. Sci. 53, 511–537 (2002).
Ekman, P., Hager, J. C. & Friesen, W. V. The symmetry of emotional and deliberate facial actions. Psychophysiology 18, 101–106 (1981).
Cohn, J. F. & Schmidt, K. L. The timing of facial motion in posed and spontaneous smiles. Int. J. Wavelets Multiresolution Inf. Process. 2, 121–132 (2004).
Raheja, J. L. & Gupta, J. Distinguishing Facial Expressions: Genuine Vs Fake. Int. J. Recent Trends Eng. Technol. 3 (2010).
Diogo, R. et al. The head and neck muscles of the serval and tiger: Homologies, evolution, and proposal of a mammalian and a veterinary muscle ontology. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 295, 2157–2178 (2012).
Damasio, A. Emotions and feelings: A neurobiological perspective. In Feelings and Emotions: The Amsterdam Symposium (eds. Manstead, A. S. R., Frijda, N. & Fischer, A.) 49–57 (Cambridge University Press, 2004).
Damasio, A. Fundamental feelings. Nature 413, 781 (2001).
Schmidt, K. L. & Cohn, J. F. Human facial expressions as adaptations: Evolutionary questions in facial expression research. Am. J. Phys. Anthropol. 116, 3–24 (2001).
Waller, B. M., Cray, J. J. & Burrows, A. M. Selection for universal facial emotion. Emotion 8, 435–439 (2008).
Beerda, B., Schilder, M. B., van Hooff, J. A., de Vries, H. W. & Mol, J. A. Behavioural, saliva cortisol and heart rate responses to different types of stimuli in dogs. Appl. Anim. Behav. Sci. 58, 365–381 (1998).
Stellato, A. C., Flint, H. E., Widowski, T. M., Serpell, J. A. & Niel, L. Assessment of fear-related behaviours displayed by companion dogs (Canis familiaris) in response to social and non-social stimuli. Appl. Anim. Behav. Sci. 188, 84–90 (2017).
Lund, J. D. & Jørgensen, M. C. Behaviour patterns and time course of activity in dogs with separation problems. Appl. Anim. Behav. Sci. 63, 219–236 (1999).
Beerda, B., Schilder, M. B., van Hooff, J. A. & de Vries, H. W. Manifestations of chronic and acute stress in dogs. Appl. Anim. Behav. Sci. 52, 307–319 (1997).
Hecht, J. & Horowitz, A. Introduction to dog behaviour. In Animal Behavior for Shelter Veterinarians and Staff (eds. Weiss, E., Heather Mohan-Gibbons & Stephen Zawistowski) (Wiley-Blackwell, 2015).
Vilà, C. et al. Multiple and ancient origins of the domestic dog. Science 276, 1687–1689 (1997).
Hare, B. & Tomasello, M. Human-like social skills in dogs? Trends Cogn. Sci. 9, 439–444 (2005).
Konok, V., Nagy, K. & Miklósi, Á. How do humans represent the emotions of dogs? The resemblance between the human representation of the canine and the human affective space. Appl. Anim. Behav. Sci. 162, 37–46 (2015).
Hintze, S., Smith, S., Patt, A., Bachmann, I. & Würbel, H. Are eyes a mirror of the soul? What eye wrinkles reveal about a horse’s emotional state. PLOS ONE 11, e0164017 (2016).
Boissy, A. et al. Cognitive sciences to relate ear postures to emotions in sheep. Anim. Welf. 20, 47 (2011).
Swielim, G. E. A. Atlas - Anatomy of sheep. (The Academic Bookshop, Egyptian Joint-Stock Co., 2006).
Parr, L. A. & Waller, B. M. Understanding chimpanzee facial expression: insights into the evolution of communication. Soc. Cogn. Affect. Neurosci. 1, 221–228 (2006).
Keysers, C. & Perrett, D. I. Demystifying social cognition: a Hebbian perspective. Trends Cogn. Sci. 8, 501–507 (2004).
Bruce, V. & Young, A. W. Face perception. (Psychology Press, 2012).
Tami, G. & Gallagher, A. Description of the behaviour of domestic dog (Canis familiaris) by experienced and inexperienced people. Appl. Anim. Behav. Sci. 120, 159–169 (2009).
Colombo, E. S. & Prato-Previde, E. Empathy and recognition of dogs’ (canis familiaris) emotions: a pilot focusing on vets. J. Vet. Behav. Clin. Appl. Res. 9, e18 (2014).
Kerswell, K. J., Bennett, P., Butler, K. L. & Hemsworth, P. H. Self-Reported Comprehension Ratings of Dog Behavior by Puppy Owners. Anthrozoos Multidiscip. J. Interact. People Anim. 22, 183–193 (2009).
Mariti, C. et al. Perception of dogs’ stress by their owners. J. Vet. Behav. Clin. Appl. Res. 7, 213–219 (2012).
Stoeckel, L. E., Palley, L. S., Gollub, R. L., Niemi, S. M. & Evins, A. E. Patterns of brain activation when mothers view their own child and dog: An fMRI study. PLoS ONE 9, e107205 (2014).
Törnqvist, H. et al. Comparison of dogs and humans in visual scanning of social interaction. R. Soc. Open Sci. 2, 150341 (2015).
Kujala, M. V., Kujala, J., Carlson, S. & Hari, R. Dog Experts’ Brains Distinguish Socially Relevant Body Postures Similarly in Dogs and Humans. PLoS ONE 7, e39145 (2012).
Morris, P. H., Doe, C. & Godsell, E. Secondary emotions in non-primate species? Behavioural reports and subjective claims by animal owners. Cogn. Emot. 22, 3–20 (2008).
Horowitz, A. Disambiguating the “guilty look”: Salient prompts to a familiar dog behaviour. Behav. Processes 81, 447–452 (2009).
Pascalis, O. & Kelly, D. J. The origins of face processing in humans: Phylogeny and ontogeny. Perspect. Psychol. Sci. 4, 200–209 (2009).
Rugaas, T. On Talking Terms with Dogs: Calming Signals. (Dogwise Publishing, 2005).
Shepherd, K. Development of behaviour, social behaviour and communication in dogs. In BSAVA Manual of Canine and Feline Behavioural Medicine (eds. Horwitz, D. & Mills, D. S.) 13–16 (2009).
Bloom, T. & Friedman, H. Classifying dogs’ (Canis familiaris) facial expressions from photographs. Behav. Processes 96, 1–10 (2013).
Davidson, J. R. Use of benzodiazepines in social anxiety disorder, generalized anxiety disorder, and posttraumatic stress disorder. J. Clin. Psychiatry 65, 29–33 (2004).
McDuff, D. et al. Affectiva-MIT facial expression dataset (AM-FED): Naturalistic and spontaneous facial expressions collected ‘in-the-wild’. In 2013 IEEE Conference on Computer Vision and Pattern Recognition Workshops 881–888 https://doi.org/10.1109/CVPRW.2013.130 (2013).
Tomasello, M. & Call, J. Assessing the validity of ape-human comparisons: A reply to Boesch (2007). J. Comp. Psychol. 122, 449–452 (2008).
Green, G. & Saunders, R. Stimulus equivalence. In Handbook of Research Methods in Human Operant Behavior (eds. Lattal, K. & Perone, M.) (Springer Science & Business Media, 2013).
McCobb, E., Brown, E., Damiani, K. & Dodman, N. Thunderstorm phobia in dogs: an Internet survey of 69 cases. J. Am. Anim. Hosp. Assoc. 37, 319–324 (2001).
Overall, K. L. & Love, M. Dog bites to humans—demography, epidemiology, injury, and risk. J. Am. Vet. Med. Assoc. 218, 1923–1934 (2001).
Blackwell, E. J., Bradshaw, J. W. S. & Casey, R. A. Fear responses to noises in domestic dogs: Prevalence, risk factors and co-occurrence with other fear related behaviour. Appl. Anim. Behav. Sci. 145, 15–25 (2013).
Fernández-Dols, J.-M. & Crivelli, C. Emotion and expression: Naturalistic studies. Emot. Rev. 5, 24–29 (2013).
Mills, D. S., Dube, M. B. & Zulch, H. Stress and pheromonatherapy in small animal clinical behaviour. (John Wiley & Sons, 2012).
Friard, O. & Gamba, M. BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 7, 1325–1330 (2016).
Ekman, P., Friesen, W. & Hager, J. FACS investigator’s guide. (Research Nexus, 2002).
Brewer, M. B. & Crano, W. D. Research design and issues of validity. in Handbook of Research Methods in Social and Personality Psychology (eds. Reis, H. T. & Judd, C. M.) 11–26 (Cambridge University Press, 2014).
Broom, D. M. & Johnson, K. G. Stress and Animal Welfare. (Springer Science & Business Media, 1993).
O’Doherty, J. P., Deichmann, R., Critchley, H. D. & Dolan, R. J. Neural responses during anticipation of a primary taste reward. Neuron 33, 815–826 (2002).
Boissy, A. et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 92, 375–397 (2007).
Held, S. D. E. & Špinka, M. Animal play and animal welfare. Anim. Behav. 81, 891–899 (2011).
Acknowledgements
Mónica Costa, Satoko Matsubara, Pearl Gillum and Jane Harris for inter-reliability coding. School of Psychology technicians for software support. Daniel Pincheira Donoso for his comments and discussion of points in earlier versions of the manuscript. This study was funded by a University of Lincoln Research Investment Fund studentship, awarded to C.C.C.
Author information
Authors and Affiliations
Contributions
C.C., K.G. and D.M. developed the study concept and design. C.C. and D.M. selected videos. C.C. coded videos and analysed the data. C.C., K.G. and D.M. interpreted the data, and C.C. drafted the initial manuscript. All authors provided critical revisions and approved the final version.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Caeiro, C., Guo, K. & Mills, D. Dogs and humans respond to emotionally competent stimuli by producing different facial actions. Sci Rep 7, 15525 (2017). https://doi.org/10.1038/s41598-017-15091-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-15091-4
This article is cited by
-
Domestication constrains the ability of dogs to convey emotions via facial expressions in comparison to their wolf ancestors
Scientific Reports (2024)
-
Intra and interspecific audience effect on domestic dogs' behavioural displays and facial expressions
Scientific Reports (2024)
-
Advanced techniques for automated emotion recognition in dogs from video data through deep learning
Neural Computing and Applications (2024)
-
Horses (Equus caballus) facial micro-expressions: insight into discreet social information
Scientific Reports (2023)
-
Visual perception of emotion cues in dogs: a critical review of methodologies
Animal Cognition (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
