
Abstract
Alzheimer’s Disease (AD) is the most common form of dementia, characterised by extracellular amyloid deposition as plaques and intracellular neurofibrillary tangles of tau protein. As no current clinical test can diagnose individuals at risk of developing AD, the aim of this project is to evaluate a blood-based biomarker panel to identify individuals who carry this risk. We analysed the levels of 22 biomarkers in clinically classified healthy controls (HC), mild cognitive impairment (MCI) and Alzheimer’s participants from the well characterised Australian Imaging, Biomarker and Lifestyle (AIBL) study of aging. High levels of IL-10 and IL-12/23p40 were significantly associated with amyloid deposition in HC, suggesting that these two biomarkers might be used to detect at risk individuals. Additionally, other biomarkers (Eotaxin-3, Leptin, PYY) exhibited altered levels in AD participants possessing the APOE ε4 allele. This suggests that the physiology of some potential biomarkers may be altered in AD due to the APOE ε4 allele, a major risk factor for AD. Taken together, these data highlight several potential biomarkers that can be used in a blood-based panel to allow earlier identification of individuals at risk of developing AD and/or early stage AD for which current therapies may be more beneficial.
Similar content being viewed by others


Introduction
Alzheimer’s Disease (AD) is the most common form of dementia in the elderly, characterised by the accumulation of extracellular senile plaques and intracellular neurofibrillary tangles in the brain1. Senile plaques are mainly composed of amyloid-β (Aβ), which is a product of Amyloid Precursor Protein (APP) that can undergo amyloidogenic cleavage to produce Aβ, or non-amyloidogenic cleavage to produce a p3 fragment2,3. Brain functions decline with disease progression as consequence of synaptic loss and neuronal death, ultimately reducing the brain volume4,5,6,7. Currently, there is no cure for AD and current treatments are aimed at reducing the symptoms and rate of decline, rather than addressing underlying causes of the disease8,9. It has been proposed that earlier interventions for AD may be more effective but it is obvious that the diagnostic process must be improved to detect the disease either before its onset or in its early stages. As of today, definitive diagnosis of the disease can only be achieved post-mortem by examination of the brain tissue. While other analysis are available, they are not suitable for large scale screening. For instance, it is possible to detect amyloid deposition through the use of PET scans using Pittsburgh Compound B (PiB) as the radiotracer, which has proven to be effective in identifying individuals who are at risk before AD onset since the technique has demonstrated that Aβ deposition takes place years before the clinical onset10. However, this analysis is quite expensive, requires specially trained physicians and bulky machinery, which means that it cannot be used to perform large scale testing. An alternative test is the analysis of the cerebrospinal fluid (CSF). While this analysis has provided positive results in discriminating between healthy controls (HC) and AD with several biomarker panels having been proposed11,12,13,14,15, it also requires trained operators and inherently carries more risk. The collection of CSF cannot be performed too often and cannot be routinely performed to screen a large population. Because of the risks and invasiveness, many individuals would refuse to undergo such a procedure, limiting the pool of available samples. Ideally, a blood-based test for AD using serum or plasma would be the best choice, as it would be inexpensive, relatively non-invasive, widely accessible with low associated risk and sampling could be performed almost anywhere. However, despite current efforts, there is no blood test that can diagnose AD. In the recent years several groups have attempted to create a biomarker panel that would predict the transition to disease in healthy individuals or to differentiate between AD and other forms of dementia. In 2007 Ray and collaborators16 suggested a blood-based biomarker panel of 18 proteins that was able to predict the conversion to AD 2–6 years later. As this panel was considered a milestone in the field, other groups have subsequently attempted to confirm these results using different cohorts. In 2008, another report indicated that a panel of 5 proteins from the former 18-protein panel was sufficient to distinguish controls from AD with the same accuracy17. In subsequent studies of the former 18-protein panel, only 3 and 5 proteins were found to be associated with AD, respectively18,19. Our group has recently published an 18-protein panel which was able to distinguish between healthy controls and AD20, while others have performed similar analysis in their cohorts21,22,23,24. Some groups have suggested panels of 3, 4 or 11 proteins that were able to distinguish between AD and controls25,26,27. A common denominator in all these studies is the heterogenetic time-points samples were collected with regards to the disease, the biomarkers evaluated and the statistical analysis used to validate effective panels. In our analysis we evaluated 22 biomarkers at 2 time points, in order to determine their association with the disease both cross-sectionally and longitudinally. This allowed us to examine whether any of these biomarkers were predictive of the conversion from healthy controls to MCI or AD and to determine their association with Aβ deposition in the cognitively healthy brain. Together, this analysis provides additional data which could help in finding a specific biomarker panel to distinguish between healthy and AD participants and/or identify those at risk of developing AD.
Materials and Methods
The AIBL Cohort
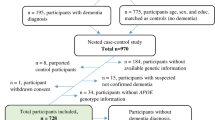
The AIBL study was approved by the ethics committees of St. Vincent’s Health, Hollywood Private Hospital, Austin Health and Edith Cowan University (Australia). All methods were performed in accordance with the relevant guidelines and regulations. A total of 1176 individuals were enrolled in the AIBL study and all volunteers gave written and informed consent before participating in our study. AIBL is a prospective longitudinal study in which healthy controls and patients are evaluated every 18 months. Individuals were evaluated in the morning, after an overnight fast. During each assessment, several body parameters, including weight, blood pressure and pulse rate were recorded, after which blood was drawn and collected in EDTA tubes for subsequent processing and analysis of the plasma28. Cognitive evaluations were then performed29,30. A more detailed description of the recruitment process has been previously described28. The diagnostic classifications were performed in accordance with the NINCDS-ARDA criteria31,32. For this study a total of 665 participants whom had their blood drawn at 18 and 54 months were included.
Blood collection and APOE genotype
Plasma was isolated from whole blood collected in EDTA tubes by centrifugation, aliquot and stored at −80 °C. APOE status was determined by genotyping cells from whole blood as previously described33.
Plasma biomarker Assay
22 analytes (IL-1α, IL-1β, IL-5, IL-6, IL-7, IL-10, IL-12/23p40, IL-13, IL-15, IL-17, EGF, EGFR, Eotaxin-3, Leptin, Angiopoietin-2, MCP-1, MMP-2, MIP-1α, PYY, SCF, TARC and TNF-α) were measured using custom assays (MesoScale Diagnostics, Maryland, USA). Briefly, these analytes were spread across three multiplex assay panels (5-plex: Angiopoietin-2, SCF, EGFR, Leptin and PYY, sample dilution: 2-fold; 8-plex: EGF, IL-1α, IL-12/23p40, IL-13, IL-15, IL-17, MCP-1 and MMP-2, sample dilution: 2-fold; 9-plex: Eotaxin-3, IL-1β, IL-5, IL-6, IL-7, IL-10, MIP-1α, TARC and TNF-α, sample dilution: undiluted) where analytes were grouped together based on their dilution, antibody compatibility and optimal assay condition requirements as specified by the manufacturer. Blocking the plate (if required) was performed with buffer provided by the manufacturer, samples were diluted according to manufacturer’s instructions and incubated at room temperature for 2 hours or overnight at 4 °C, depending on the assay panel. After washing the plates 3 times with PBST (pH 7.4), the specified detection antibodies were added and plates were incubated for a further 1 hour at room temperature, followed by 3 washes with PBST. Read buffer was then added and plates were read on the SECTOR Imager (MSD, Maryland, USA).
PET scan
A selected number of individuals of the AIBL cohort (at 18 months HC PiB−, n = 94 and HC PiB+, n = 27; at 54 months HC PiB−, n = 67 and HC PiB+, n = 26) volunteered for Positron Emission Tomography with the labelled Pittsburgh compound B (11C-PiB−PET) to measure cerebral amyloid load34. PiB score, expressed as Standardized Uptake Value Ratio (SUVR), was calculated by normalizing the Standardized Uptake Value (SUV) to the cerebellar SUV. The common SUVR threshold of 1.5 was used to demarcate individuals with low amyloid deposition (PiB−, SUVR < 1.5) or high amyloid deposition (PiB+, SUVR > 1.5)10.
Statistical analysis
Statistical comparisons of markers at either 18 months or 54 months were carried out using linear models correcting for age, site, APOE ε4 allele status and gender. Results were considered significant when p < 0.05. The combined analysis over both time points simultaneously was performed using mixed modelling to accommodate correlations between observations on the same individual. Logistic regression analysis was used to determine the joint association between multiple biomarkers and group outcomes. All calculations were carried out using TIBCO Spotfire S+ ver 8.2 statistical software (TIBCO Software, Inc., Boston, MA). Note that while data in the tables are presented as pg/ml, statistical analysis was performed using the log10 transformation of the raw data to better approximate normality. For IL-5, IL-6 and IL-17 the log10 transformation was performed on the (raw data + 1) in order to accommodate the 0 values in the dataset. ROC analysis for prediction of high or low SUVR values was carried out via logistic regression comparing a base model which included age + sex + APOE genotype and an extended model including age + sex + APOE genotype + log(IL-10*IL-12/23).
Results
Demographics
The basic demographic of this study are summarized in Table 1. A total of 559 healthy controls (HC), 39 mild cognitive impaired (MCI) and 67 Alzheimer patients (AD) were evaluated. Plasma samples were analysed for all biomarkers listed. However, in spite of analysing our biomarker panel in all individuals, the difference in available data for each biomarker are due to the difference in CV across duplicates, which lead us to eliminate duplicate values with high CV. The elimination rate was different for each biomarker analysed. We used a 30% CV cut-off above which the calculated value was considered not suitable for analysis. While this value may appear high, it should be noted that cytokines levels were extremely low and often fell in the lowest section of the standard curve, where small differences in the raw value resulted in a much larger difference in the calculated result. Hence, while a low percent CV of the raw value, which would normally indicate good duplicates in the assay may translate into high CV of the calculated result. In order to keep the analysis consistent, when a sample at either 18 or 54 months was not available, its counterpart at the other time point was also removed, even if the sample fell in the readable range. Of the 22 biomarkers evaluated, IL-13 was excluded as duplicates displayed inconsistent reproducibility while IL-1α, IL-1β and MIP-1α levels were below the detectible range for most of the samples. In Table 2 we report the valid cases for each individual biomarker at both time points. In order to demonstrate that the removal of samples with high CV did not affect the average biomarker values, all samples regardless of their CV were tabulated (Supplementary Table 1). As shown, the overall averages of all samples (Supplementary Table 1) are similar to the average values of the valid cases (Table 3). Therefore the exclusion of these samples did not affect the average values of these biomarkers.
Longitudinal assessment of biomarkers
Table 3 illustrates changes in biomarkers at 2 different time points, 18 months and 54 months in non-converters, with p-values corrected for age, gender, site and APOE ε4 carriage. PYY levels were significantly higher in AD at 18 months (p = 0.02) but not at 54 months (p = 0.09). In contrast Eotaxin-3 levels were significantly higher in AD at 54 months (p = 0.03) but not at 18 months (p = 0.39). EGF and TARC levels showed increased levels in AD at 54 months (p = 0.08 for both) but not at 18 months (p = 0.48 and p = 0.62, respectively). A mixed model analysis combining the two time points (Table 4) found that only Eotaxin-3, TARC and PYY levels showed a trend (p = 0.07, p = 0.06 and p = 0.07, respectively). The p-values reported for these were for single analyte analysis, and are not significant upon Bonferroni’s correction for multiple comparisons (data not shown).
Eotaxin-3, Leptin and PYY show altered levels in a subset of AD patients (APOE ε4 carrier)
In Tables 5 and 6 we evaluated 5 biomarkers (IL-12/23p40, Eotaxin-3, Angiopoietin-2, Leptin and PYY) that had shown differences based on their APOE genotype by subdividing the non-converters group into APOE ε4− and APOE ε4+. In the APOE ε4− group (Table 5) IL-12/23p40 is slightly lower in AD at 18 months only (p = 0.08), while Leptin levels tend to be lower, although not significantly, in AD versus HC at both time points (p = 0.13 and p = 0.17, respectively). However, when evaluating the same biomarkers in the APOE ε4+ group (Table 6), Eotaxin-3, Leptin and PYY levels are higher in AD patients at both assessments. Although the mixed model analysis did not show any statistical significance overall (Table 7), both Leptin and PYY levels are significantly higher in AD/APOE ε4+ at 54 months compared to the other groups (Leptin p = 0.035, PYY p = 0.0013). We also performed the General Linear Model analysis in the AD group only comparing Leptin and PYY in APOE ε4− vs APOE ε4+ at both time points. The analysis did not show significance for Leptin (p = 0.18 and p = 0.13 at 18 and 54 months, respectively), neither for PYY (p = 0.77 and p = 0.33 at 18 and 54 months, respectively). However, the analysis itself may have been affected by the low number of cases (PYY: n = 16 and 33 for APOE ε4− and APOE ε4+, respectively and Leptin: n = 19 and 47 for APOE ε4− and APOE ε4+, respectively). Although it would have been interesting to examine the effect of participants being heterozygous or homozygous for the APOE ε4 allele, due to the very small sample size of APOE ε4 homozygous participants (for PYY n = 26 of APOE ε4 heterozygous and n = 7 of APOE ε4 homozygous; for Leptin n = 36 of APOE ε4 heterozygous and n = 11 of APOE ε4 homozygous) in the AD group, undertaking such an analysis was not feasible.
IL-10 and IL-12/23p40 are jointly associated as predictors of β–amyloid load
In order to determine whether these biomarkers can be useful diagnostically or predictively for development of AD, we analysed the biomarker levels in all HC (non-converters and converters) who had undergone amyloid imagining and stratified them into PiB− (SUVR < 1.5) or PiB+ (SUVR > 1.5). The results in Table 8 illustrate that only IL-12/23 p40 levels at 54 months were significantly higher in HC PiB+ versus HC PiB− (p = 0.023), while their increase at 18 months was not (p = 0.11). Similarly, IL-10 also showed a trend for increased levels in HC PiB+ at both time points (p = 0.13 and p = 0.15). No altered levels of any other biomarkers were associated with HC in which amyloid deposition is present (PiB+). The p-values reported here are performed for single analyte analysis and they are not significant upon Bonferroni’s correction for multiple comparisons (data not shown). Logistic regression analysis of the two markers jointly with PiB+ level as the outcome indicated that they were jointly significant with the effect restricted to the (log) product of the two markers (Table 9, p = 0.039 at 18 months and p = 0.017 at 54 months). ROC analysis performed at 54 months indicated an increased area under curve (AUC = 0.805) when including the IL-10*IL-12/23p40 to the base model (age + sex + APOE ε4) for which the AUC = 0.779 (Fig. 1). Similar results were obtained when evaluating our HC at 18 months (Base model AUC = 0.787; Base model + log(IL-10*IL-12/23p40) AUC = 0.802, data not shown).

ROC analysis performed at 54 months. The black line represent the base model (age + sex + APOE ε4), while the red line represents the base model + log(IL-10*IL-12/23p40).
We also evaluated the levels of the same biomarkers in all HC at 18 months based on their classification at 54 months, comparing the non-converter participants, who did not develop AD at 54 months, versus those who did convert to MCI or AD (data not shown). Here, at 18 months IL-12/23p40 and IL-10 levels tended to be higher in the converter group versus the non-converters. However, the low sample size of converters may be a constraint to the statistical analysis.
Discussion
AD is the leading cause of dementia in the elderly, but current methods of ante-mortem diagnosis lack accuracy and specificity. Developing a biomarker panel that can provide early diagnosis of AD or indicate level of risk in healthy individuals remains a considerable challenge. Many studies have reported biomarker panels of 10 or more biomarkers but tended to be cross-sectional. Our study is longitudinal with 22 biomarkers. It was performed on the well characterised AIBL cohort, using biomarkers that have previously been associated with AD in previous studies. In order to provide a more detailed evaluation, we have assessed biomarkers at 2 different time points to determine whether biomarker changes were consistent over time, or specific to the early or late stages of the disease.
We evaluated biomarkers in non-converters, individuals whose cognition was unchanged at both time points. As shown in Table 3, only 4 biomarkers displayed altered levels between HC and AD. We found that PYY was significantly higher in AD individuals at 18 months and follows a similar trend at 54 months. PYY is a molecule that belongs to the pancreatic polypeptide family and has been linked to aluminium metabolism in AD35. Previous work did not show any differences in blood levels of PYY between controls and AD36.
Similarly, Eotaxin-3 significantly increased in AD at 54 months but not at 18 months. Other studies have reported that higher levels of Eotaxin-3 in plasma and CSF were observed and were associated with prodromal AD11,24,37. In our biomarker panel, 2 more analytes (EGF and TARC) appeared to be associated with AD. For instance, a decrease in EGF levels has previously been associated with AD in a study with a biomarker panel of 18 proteins to predict the conversion from MCI to AD16. However, subsequent studies using the same 18-protein panel failed to validate it as only 3 and 5 of those proteins were significantly associated with AD18,19. While EGF was significantly associated with AD in both studies, its levels in the AD participants were increased rather than decreased. These data corroborates with another study that also showed increased EGF levels in AD27. Our data indicated a trend towards increased EGF at 54 months but not at 18 months. However, this difference was mostly due to a more severe drop of EGF levels in the HC, rather than an increase in the AD group (Table 3).
TARC, also known as CCL17, is a chemokine constitutively expressed in thymus whose natural ligand is CCR438. Similar to EGF, our data indicated a significant increase of TARC at 54 month but not at 18 months. While it can be argued that these biomarkers are likely more related to later stage AD, we have also observed that age and site were affecting biomarker levels. However, all our analysis were corrected for age, gender, APOE genotype and site. The combined mixed model analysis (Table 4), which also accounted for longitudinal changes showed that Eotaxin-3, TARC and PYY may represent a suitable stable biomarker for differentiating between HC and AD (p = 0.07, p = 0.06 and p = 0.07, respectively). While the overall analysis provides a broad picture that can be used to find a biomarker panel with stable analytes over time for the detection of AD, it is always important to remember that APOE genotype is currently a major risk factor for AD, therefore the presence of the ε4 allele must always be considered when evaluating biomarkers.
We have therefore evaluated 5 biomarkers that demonstrated clear differences according to APOE ε4 status. The data show consistent increases in plasma Eotaxin-3 levels in AD individuals carrying APOE ε4 at both time points. However, a large variability was also observed amongst these individuals, suggesting that the presence of the ε4 allele could increase Eotaxin-3, but was not sufficient by itself to cause this increase. Other factors that may be involved in some but not all of the AD/APOE ε4+ participants might also play a role in altering Eotaxin-3 levels. In parallel, two other biomarkers, Leptin and PYY, have shown increased levels in the AD/APOE ε4+ subgroup at both time points, strongly suggesting that in the AD/APOE ε4+ subgroup, these analytes may represent a stable biomarker alternative in the APOE ε4− carrier individuals, while seeking for a biomarker panel sufficient for AD detection. It is interesting to note that both Leptin and PYY perform similar functions as they affect the metabolism by reducing appetite after a meal. A study in APOE ε4 knock-in mice has shown that Leptin receptors are specifically reduced in the hippocampal neurons, similar to the levels observed in Tg2576 mice, but not in APOE ε3 knock-in mice39. This suggests that APOE ε4 carriers may develop some resistance to Leptin signalling and that increased levels of Leptin could be a consequence of this altered metabolism. Similarly, NPY receptor levels (the receptor for PYY) were reportedly lower in the cortex and in the hippocampus of AD brains40, although it is not known whether their levels are affected by APOE genotype.
While finding a biomarker panel that can differentiate between HC and AD remains a primary target, another important goal is to predict the development of AD. This would allow interventions before onset of the disease. In order to determine if this can be achieved with our approach, we evaluated our biomarkers with respect to the Aβ deposition in the brains of HC participants. Our analysis therefore tested the association of biomarker levels with the neocortical SUVR score rather than specific areas. Since the neocortical regions are the most commonly affected by Aβ deposition in AD41, the detection of Aβ using this approach should be more consistent.
Because it is understood that this deposition starts years before the clinical manifestation of the disease and it is accepted that HC with high amyloid load are more likely to progress to AD in the future10, we subdivided this group into PiB− (SUVR < 1.5, low amyloid deposition) and PiB+ (SUVR > 1.5, high amyloid deposition). When the biomarker levels were analysed with this stratification we found that at 54 months, IL-12/23p40 levels were significantly higher in PiB+ HC than the PiB−, while the earlier 18 month assessment only exhibited a trend. IL-10 levels also followed this trend at both time points in PiB+ compared to PiB−, while all other biomarkers did not display any association with amyloid load.
Consistent with studies in mice, we found that IL-10 and IL-12/23p40 are jointly associated with the amyloid deposition in the brain, indicating that several factors are involved in this process42,43. IL-12 is a heterodimeric cytokine produced by a number of cells associated with the immune system with a broad range of activities and acts on T- and natural killer cells. It shares the p40 subunit which is also common to IL-2344 and has been previously associated with AD. IL-12 is expressed by activated macrophages which serves as an inducer for T-helper cell (Th1) development45, which have important roles in the adaptability of the immune system and its regulation. At present, the connection of the IL-12/23p40 subunit with AD is not well understood. However, polymorphisms of the gene encoding it have been associated with autoimmune diseases46,47,48 and susceptibility for multiple sclerosis49,50,51,52,53,54, particularly those that increase its production55.
Reduced levels seems to lower the inflammatory response, as seen with mice deficient in the p40 subunit which developed less severe forms of autoimmune diseases56. It is not known if this is consistent in the brain, but it was demonstrated in other mouse models that inhibition of IL-12/23 p40 was associated with reduced Aβ levels57,58. This seems to agree with a recent small scale study using CSF from human participants that showed a positive correlation of IL-12/23 p40 with CSF Aβ1-4259, and our own current work here. This evidence strengthens the connection between brain amyloid load and the immune system.
While there is still clearly much to learn about this link, a clue may lie with the cytokines sharing the p40 subunit. IL-12 production from whole blood after mitogen stimulation was found to be lower in cells from AD patients compared to healthy controls60. Another study showed IL-12 levels were initially elevated in mild AD and decreased as the disease progressed61. These findings indicate that while IL-12 and other cytokines may play an early role in development of AD, the capacity of cells to produce these cytokines is diminished with disease progression, indicating a reduced immune response in AD62. This reduction of IL-12 during disease progression appears to be generalised and not confined to the blood and periphery, since IL-12 levels were also found to be lower in the CSF of AD patients63.
IL-23, the other heterodimeric cytokine which shares the p40 subunit is important in the inflammatory response during infection, affecting both innate and adaptive immune system functions64,65. Inflammation is considered a feature of AD pathology and this cytokine is known to promote inflammatory responses, such as upregulation of the matrix metalloprotease MMP956. It was even considered to be a primary cytokine for autoimmune inflammation in the brain66.
Like IL-12, not much is actually known about its primary role in the brain. However, many studies have reported associations with AD. IL-23 levels appeared to be increased in AD67, which may be a direct consequence of the increased numbers of IL-23-producing cells in AD68,69. As IL-12 and IL-23 displayed different modulation in AD, it brings into question the roles of the other subunits of these two interleukins, namely p35 and p19 for IL-12 and IL-23, respectively. Regardless, both IL-10 and IL-12/23p40 may be considered as viable biomarkers in a broader panel to help identify individuals at risk of developing AD, where early therapeutic intervention may delay the onset of the disease. Overall, IL-12 and IL-23 are considered pro-inflammatory cytokines mainly secreted by antigen-presenting cells, such as dendritic cells and macrophages, with extensive roles in in the immune response and the differentiation of Th1 and Th17 subset of T-helper cells44,66,70,71,72,73. Conversely, IL-10 is one of the more potent anti-inflammatory cytokines which inhibits the secretion of Th1 cytokines from stimulated cells74,75,76,77.
A problem that may arise from this kind of analysis is the extreme heterogeneity of the studies. Various biomarker panels have given different results based on the different proteins and methods used in the evaluation. Even when the same proteins were examined in different studies, their involvement in AD was inconsistent. Furthermore, the sources of these analytes is important, i.e. whether it is CSF or blood-derived specimens. Inconsistencies across different studies may be due to a combination of factors, including different assay platforms but particularly the stage of the disease, as several proteins may be affected during early or late stages in AD. Although blood was typically drawn from fasting individuals, the time of the day is another factor that should be considered as many proteins have a day-night cycle that severely affects their levels over the daily cycle. The statistical analysis used to determine the significance of the biomarker can play a role in determining which analytes are significant and which are not. Finally, the number of times blood is drawn and examined can be a very important factor as it would provide a more detailed and regular analysis, eliminating the risk of false positive/false negative results that can easily occur during a single time point analysis.
However, in our study we tried to address these issues and detected two biomarkers that are jointly associated with brain amyloid deposition in the HC that can be used in a broader biomarker panel for the detection of individuals at higher risk of developing AD. As it is increasingly necessary to identify individuals at risk for AD, such findings would allow for early treatments with the ultimate goal to stop or delay the onset of the disease.
References
Price, J. L. & Morris, J. C. Tangles and plaques in nondemented aging and “preclinical” Alzheimer’s disease. Annals of neurology 45, 358–368 (1999).
Krishnaswamy, S., Verdile, G., Groth, D., Kanyenda, L. & Martins, R. N. The structure and function of Alzheimer’s gamma secretase enzyme complex. Critical reviews in clinical laboratory sciences 46, 282–301, https://doi.org/10.3109/10408360903335821 (2009).
Zhang, H., Ma, Q., Zhang, Y. W. & Xu, H. Proteolytic processing of Alzheimer’s beta-amyloid precursor protein. Journal of neurochemistry 120(Suppl 1), 9–21, https://doi.org/10.1111/j.1471-4159.2011.07519.x (2012).
Arendt, T., Bigl, V., Arendt, A. & Tennstedt, A. Loss of neurons in the nucleus basalis of Meynert in Alzheimer’s disease, paralysis agitans and Korsakoff’s Disease. Acta neuropathologica 61, 101–108 (1983).
Gomez-Isla, T. et al. Profound loss of layer II entorhinal cortex neurons occurs in very mild Alzheimer’s disease. The Journal of neuroscience: the official journal of the Society for Neuroscience 16, 4491–4500 (1996).
West, M. J., Coleman, P. D., Flood, D. G. & Troncoso, J. C. Differences in the pattern of hippocampal neuronal loss in normal ageing and Alzheimer’s disease. Lancet (London, England) 344, 769–772 (1994).
Whitehouse, P. J. et al. Alzheimer’s disease and senile dementia: loss of neurons in the basal forebrain. Science (New York, N.Y.) 215, 1237–1239 (1982).
Rogers, S. L., Farlow, M. R., Doody, R. S., Mohs, R. & Friedhoff, L. T. A 24-week, double-blind, placebo-controlled trial of donepezil in patients with Alzheimer’s disease. Donepezil Study Group. Neurology 50, 136–145 (1998).
Tariot, P. N. et al. Memantine treatment in patients with moderate to severe Alzheimer disease already receiving donepezil: a randomized controlled trial. Jama 291, 317–324, https://doi.org/10.1001/jama.291.3.317 (2004).
Villemagne, V. L. et al. Amyloid beta deposition, neurodegeneration, and cognitive decline in sporadic Alzheimer’s disease: a prospective cohort study. The Lancet. Neurology 12, 357–367, https://doi.org/10.1016/s1474-4422(13)70044-9 (2013).
Craig-Schapiro, R. et al. Multiplexed immunoassay panel identifies novel CSF biomarkers for Alzheimer’s disease diagnosis and prognosis. PloS one 6, e18850, https://doi.org/10.1371/journal.pone.0018850 (2011).
Hu, W. T. et al. Novel CSF biomarkers for frontotemporal lobar degenerations. Neurology 75, 2079–2086, https://doi.org/10.1212/WNL.0b013e318200d78d (2010).
Khan, W. et al. A Subset of Cerebrospinal Fluid Proteins from a Multi-Analyte Panel Associated with Brain Atrophy, Disease Classification and Prediction in Alzheimer’s Disease. PloS one 10, e0134368, https://doi.org/10.1371/journal.pone.0134368 (2015).
Perrin, R. J. et al. Identification and validation of novel cerebrospinal fluid biomarkers for staging early Alzheimer’s disease. PloS one 6, e16032, https://doi.org/10.1371/journal.pone.0016032 (2011).
Simonsen, A. H. et al. Identification of a novel panel of cerebrospinal fluid biomarkers for Alzheimer’s disease. Neurobiology of aging 29, 961–968, https://doi.org/10.1016/j.neurobiolaging.2007.01.011 (2008).
Ray, S. et al. Classification and prediction of clinical Alzheimer’s diagnosis based on plasma signaling proteins. Nature medicine 13, 1359–1362, https://doi.org/10.1038/nm1653 (2007).
Gomez Ravetti, M. & Moscato, P. Identification of a 5-protein biomarker molecular signature for predicting Alzheimer’s disease. PloS one 3, e3111, https://doi.org/10.1371/journal.pone.0003111 (2008).
Bjorkqvist, M., Ohlsson, M., Minthon, L. & Hansson, O. Evaluation of a previously suggested plasma biomarker panel to identify Alzheimer’s disease. PloS one 7, e29868, https://doi.org/10.1371/journal.pone.0029868 (2012).
Marksteiner, J. et al. Five out of 16 plasma signaling proteins are enhanced in plasma of patients with mild cognitive impairment and Alzheimer’s disease. Neurobiology of aging 32, 539–540, https://doi.org/10.1016/j.neurobiolaging.2009.03.011 (2011).
Doecke, J. D. et al. Blood-based protein biomarkers for diagnosis of Alzheimer disease. Archives of neurology 69, 1318–1325, https://doi.org/10.1001/archneurol.2012.1282 (2012).
O’Bryant, S. E. et al. A blood-based screening tool for Alzheimer’s disease that spans serum and plasma: findings from TARC and ADNI. PloS one 6, e28092, https://doi.org/10.1371/journal.pone.0028092 (2011).
O’Bryant, S. E. et al. A blood-based algorithm for the detection of Alzheimer’s disease. Dementia and geriatric cognitive disorders 32, 55–62, https://doi.org/10.1159/000330750 (2011).
Soares, H. D. et al. Identifying early markers of Alzheimer’s disease using quantitative multiplex proteomic immunoassay panels. Annals of the New York Academy of Sciences 1180, 56–67, https://doi.org/10.1111/j.1749-6632.2009.05066.x (2009).
Soares, H. D. et al. Plasma biomarkers associated with the apolipoprotein E genotype and Alzheimer disease. Archives of neurology 69, 1310–1317, https://doi.org/10.1001/archneurol.2012.1070 (2012).
Hu, W. T. et al. Plasma multianalyte profiling in mild cognitive impairment and Alzheimer disease. Neurology 79, 897–905, https://doi.org/10.1212/WNL.0b013e318266fa70 (2012).
Johnstone, D., Milward, E. A., Berretta, R. & Moscato, P. Multivariate protein signatures of pre-clinical Alzheimer’s disease in the Alzheimer’s disease neuroimaging initiative (ADNI) plasma proteome dataset. PloS one 7, e34341, https://doi.org/10.1371/journal.pone.0034341 (2012).
Laske, C. et al. Identification of a blood-based biomarker panel for classification of Alzheimer’s disease. The international journal of neuropsychopharmacology / official scientific journal of the Collegium Internationale Neuropsychopharmacologicum (CINP) 14, 1147–1155, https://doi.org/10.1017/s1461145711000459 (2011).
Ellis, K. A. et al. TheAustralian Imaging, Biomarkers and Lifestyle (AIBL) study of aging: methodology and baseline characteristics of 1112 individuals recruited for a longitudinal study of Alzheimer’s disease. Int Psychogeriatr 21, 672–687, https://doi.org/10.1017/s1041610209009405 (2009).
Folstein, M. F., Folstein, S. E. & McHugh, P. R. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. Journal of psychiatric research 12, 189–198 (1975).
Saxton, J. et al. Normative data on the Boston Naming Test and two equivalent 30-item short forms. The Clinical neuropsychologist 14, 526–534, https://doi.org/10.1076/clin.14.4.526.7204 (2000).
McKhann, G. et al. Clinical diagnosis of Alzheimer’s disease: report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 34, 939–944 (1984).
Winblad, B. et al. Mild cognitive impairment–beyond controversies, towards a consensus: report of the International Working Group on Mild Cognitive Impairment. J Intern Med 256, 240–246, https://doi.org/10.1111/j.1365-2796.2004.01380.x (2004).
Gupta, V. B. et al. Plasma apolipoprotein E and Alzheimer disease risk: the AIBL study of aging. Neurology 76, 1091–1098, https://doi.org/10.1212/WNL.0b013e318211c352 (2011).
Pike, K. E. et al. Beta-amyloid imaging and memory in non-demented individuals: evidence for preclinical Alzheimer’s disease. Brain 130, 2837–2844, https://doi.org/10.1093/brain/awm238 (2007).
Croom, J. & Taylor, I. L. Neuropeptide Y, peptide YY and aluminum in Alzheimer’s disease: is there an etiological relationship? Journal of inorganic biochemistry 87, 51–56 (2001).
Ahmed, R. M. et al. Eating behavior in frontotemporal dementia: Peripheral hormones vs hypothalamic pathology. Neurology 85, 1310–1317, https://doi.org/10.1212/wnl.0000000000002018 (2015).
Westin, K. et al. CCL2 is associated with a faster rate of cognitive decline during early stages of Alzheimer’s disease. PloS one 7, e30525, https://doi.org/10.1371/journal.pone.0030525 (2012).
Imai, T. et al. The T cell-directed CC chemokine TARC is a highly specific biological ligand for CC chemokine receptor 4. The Journal of biological chemistry 272, 15036–15042 (1997).
Maioli, S. et al. Alterations in brain leptin signalling in spite of unchanged CSF leptin levels in Alzheimer’s disease. Aging cell 14, 122–129, https://doi.org/10.1111/acel.12281 (2015).
Martel, J. C., Alagar, R., Robitaille, Y. & Quirion, R. Neuropeptide Y receptor binding sites in human brain. Possible alteration in Alzheimer’s disease. Brain research 519, 228–235 (1990).
Serrano-Pozo, A., Frosch, M. P., Masliah, E. & Hyman, B. T. Neuropathological alterations in Alzheimer disease. Cold Spring Harbor perspectives in medicine 1, a006189, https://doi.org/10.1101/cshperspect.a006189 (2011).
Chakrabarty, P. et al. IL-10 alters immunoproteostasis in APP mice, increasing plaque burden and worsening cognitive behavior. Neuron 85, 519–533, https://doi.org/10.1016/j.neuron.2014.11.020 (2015).
Guillot-Sestier, M. V. et al. Il10 deficiency rebalances innate immunity to mitigate Alzheimer-like pathology. Neuron 85, 534–548, https://doi.org/10.1016/j.neuron.2014.12.068 (2015).
Oppmann, B. et al. Novel p19 protein engages IL-12p40 to form a cytokine, IL-23, with biological activities similar as well as distinct from IL-12. Immunity 13, 715–725 (2000).
Radrizzani, M. et al. IL-12 inhibits apoptosis induced in a human Th1 clone by gp120/CD4 cross-linking and CD3/TCR activation or by IL-2 deprivation. Cellular immunology 161, 14–21, https://doi.org/10.1006/cimm.1995.1003 (1995).
Capon, F. et al. Sequence variants in the genes for the interleukin-23 receptor (IL23R) and its ligand (IL12B) confer protection against psoriasis. Human genetics 122, 201–206, https://doi.org/10.1007/s00439-007-0397-0 (2007).
Cargill, M. et al. A large-scale genetic association study confirms IL12B and leads to the identification of IL23R as psoriasis-risk genes. American journal of human genetics 80, 273–290, https://doi.org/10.1086/511051 (2007).
Glas, J. et al. Analysis of IL12B gene variants in inflammatory bowel disease. PloS one 7, e34349, https://doi.org/10.1371/journal.pone.0034349 (2012).
Alloza, I. et al. Interleukin-12p40 polymorphism and susceptibility to multiple sclerosis. Annals of neurology 52, 524–525, https://doi.org/10.1002/ana.10348 (2002).
Begovich, A. B. et al. The autoimmune disease-associated IL12B and IL23R polymorphisms in multiple sclerosis. Human immunology 68, 934–937, https://doi.org/10.1016/j.humimm.2007.09.005 (2007).
Illes, Z. et al. 3′UTR C2370A allele of the IL-23 receptor gene is associated with relapsing-remitting multiple sclerosis. Neuroscience letters 431, 36–38, https://doi.org/10.1016/j.neulet.2007.11.015 (2008).
Nunez, C. et al. IL23R: a susceptibility locus for celiac disease and multiple sclerosis? Genes and immunity 9, 289–293, https://doi.org/10.1038/gene.2008.16 (2008).
Roos, I. M., Kockum, I. & Hillert, J. The interleukin 23 receptor gene in multiple sclerosis: a case-control study. Journal of neuroimmunology 194, 173–180, https://doi.org/10.1016/j.jneuroim.2007.11.011 (2008).
van Veen, T. et al. Interleukin-12p40 genotype plays a role in the susceptibility to multiple sclerosis. Annals of neurology 50, 275 (2001).
Javan, M. R. et al. An interleukin 12 B single nucleotide polymorphism increases IL-12p40 production and is associated with increased disease susceptibility in patients with relapsing-remitting multiple sclerosis. Neurological research 39, 435–441, https://doi.org/10.1080/01616412.2017.1301623 (2017).
Langowski, J. L. et al. IL-23 promotes tumour incidence and growth. Nature 442, 461–465, https://doi.org/10.1038/nature04808 (2006).
Tan, M. S. et al. IL12/23 p40 inhibition ameliorates Alzheimer’s disease-associated neuropathology and spatial memory in SAMP8 mice. Journal of Alzheimer’s disease: JAD 38, 633–646, https://doi.org/10.3233/jad-131148 (2014).
Vom Berg, J. et al. Inhibition of IL-12/IL-23 signaling reduces Alzheimer’s disease-like pathology and cognitive decline. Nature medicine 18, 1812–1819, https://doi.org/10.1038/nm.2965 (2012).
Johansson, P. et al. Reduced cerebrospinal fluid concentration of interleukin-12/23 subunit p40 in patients with cognitive impairment. PloS one 12, e0176760, https://doi.org/10.1371/journal.pone.0176760 (2017).
Richartz, E. et al. Diminished production of proinflammatory cytokines in patients with Alzheimer’s disease. Dementia and geriatric cognitive disorders 19, 184–188, https://doi.org/10.1159/000083497 (2005).
Motta, M., Imbesi, R., Di Rosa, M., Stivala, F. & Malaguarnera, L. Altered plasma cytokine levels in Alzheimer’s disease: correlation with the disease progression. Immunology letters 114, 46–51, https://doi.org/10.1016/j.imlet.2007.09.002 (2007).
Richartz, E. et al. Decline of immune responsiveness: a pathogenetic factor in Alzheimer’s disease? Journal of psychiatric research 39, 535–543, https://doi.org/10.1016/j.jpsychires.2004.12.005 (2005).
Rentzos, M. et al. Interleukin-12 is reduced in cerebrospinal fluid of patients with Alzheimer’s disease and frontotemporal dementia. Journal of the neurological sciences 249, 110–114, https://doi.org/10.1016/j.jns.2006.05.063 (2006).
Kikly, K., Liu, L., Na, S. & Sedgwick, J. D. The IL-23/Th(17) axis: therapeutic targets for autoimmune inflammation. Current opinion in immunology 18, 670–675, https://doi.org/10.1016/j.coi.2006.09.008 (2006).
Memari, B. et al. Engagement of the Aryl Hydrocarbon Receptor in Mycobacterium tuberculosis-Infected Macrophages Has Pleiotropic Effects on Innate Immune Signaling. Journal of immunology (Baltimore, Md.: 1950) 195, 4479–4491, https://doi.org/10.4049/jimmunol.1501141 (2015).
Cua, D. J. et al. Interleukin-23 rather than interleukin-12 is the critical cytokine for autoimmune inflammation of the brain. Nature 421, 744–748, https://doi.org/10.1038/nature01355 (2003).
Chen, J. M., Jiang, G. X., Li, Q. W., Zhou, Z. M. & Cheng, Q. Increased serum levels of interleukin-18, -23 and -17 in Chinese patients with Alzheimer’s disease. Dementia and geriatric cognitive disorders 38, 321–329, https://doi.org/10.1159/000360606 (2014).
Saresella, M. et al. Increased activity of Th-17 and Th-9 lymphocytes and a skewing of the post-thymic differentiation pathway are seen in Alzheimer’s disease. Brain, behavior, and immunity 25, 539–547, https://doi.org/10.1016/j.bbi.2010.12.004 (2011).
Saresella, M. et al. A complex proinflammatory role for peripheral monocytes in Alzheimer’s disease. Journal of Alzheimer’s disease: JAD 38, 403–413, https://doi.org/10.3233/jad-131160 (2014).
Langrish, C. L. et al. IL-23 drives a pathogenic T cell population that induces autoimmune inflammation. The Journal of experimental medicine 201, 233–240, https://doi.org/10.1084/jem.20041257 (2005).
Langrish, C. L. et al. IL-12 and IL-23: master regulators of innate and adaptive immunity. Immunological reviews 202, 96–105, https://doi.org/10.1111/j.0105-2896.2004.00214.x (2004).
Ma, X. & Trinchieri, G. Regulation of interleukin-12 production in antigen-presenting cells. Advances in immunology 79, 55–92 (2001).
Vignali, D. A. & Kuchroo, V. K. IL-12 family cytokines: immunological playmakers. Nature immunology 13, 722–728, https://doi.org/10.1038/ni.2366 (2012).
Couper, K. N., Blount, D. G. & Riley, E. M. IL-10: the master regulator of immunity to infection. Journal of immunology (Baltimore, Md.: 1950) 180, (5771–5777 (2008).
Joss, A., Akdis, M., Faith, A., Blaser, K. & Akdis, C. A. IL-10 directly acts on T cells by specifically altering the CD28 co-stimulation pathway. European journal of immunology 30, 1683–1690, https://doi.org/10.1002/1521-4141(200006)30:6<1683::aid-immu1683>3.0.co;2-a (2000).
Murray, P. J. The primary mechanism of the IL-10-regulated antiinflammatory response is to selectively inhibit transcription. Proceedings of the National Academy of Sciences of the United States of America 102, 8686–8691, https://doi.org/10.1073/pnas.0500419102 (2005).
Saraiva, M. & O’Garra, A. The regulation of IL-10 production by immune cells. Nature reviews. Immunology 10, 170–181, https://doi.org/10.1038/nri2711 (2010).
Acknowledgements
We thank all the participants who took part in this study and the clinicians who referred participants. The AIBL study (www.AIBL.csiro.au) is a collaboration between CSIRO, Edith Cowan University (ECU), National Ageing Research Institute (NARI), The Florey Institute of Neuroscience and Mental Health (FINMH) and Austin Health. It also received support from Hollywood Private Hospital, CogState Ltd., and Sir Charles Gairdner Hospital and funding support from Alzheimer’s Australia (AA), CSIRO, the Science and Industry Endowment Fund, McCusker Alzheimer’s Research Foundation, Brightfocus, USA and the WA Dept. of Health, as well as industry sources. The authors acknowledge the financial support of the Cooperative Research Centre (CRC) for Mental Health, which is an Australian Government Initiative. Pfizer International has provided financial support to assist with analysis of blood samples and to further the AIBL research program.
Author information
Authors and Affiliations
Consortia
Contributions
S.P., V.B.G., E.H. collected data and drafted the manuscript, I.J. provided data analysis and interpretation, J.D., S.O., A.I.B., C.C.R., V.L.V., D.A., C.L.M. and the A.I.B.L. Research Group reviewed the manuscript, R.N.M. designed the work and reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
S. O’Bryant has received honorarium for involvement in Roche Diagnostics Advisory Board and has multiple patents pending regarding blood-based biomarkers in neurodegenerative diseases. He is supported by funds from the NIH/NIA (AG051848).
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
A comprehensive list of consortium members appears at the end of the paper.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pedrini, S., Gupta, V.B., Hone, E. et al. A blood-based biomarker panel indicates IL-10 and IL-12/23p40 are jointly associated as predictors of β-amyloid load in an AD cohort. Sci Rep 7, 14057 (2017). https://doi.org/10.1038/s41598-017-14020-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-14020-9
This article is cited by
-
Evaluation of serum interleukin-12 and interleukin-4 as potential biomarkers for the diagnosis of major depressive disorder
Scientific Reports (2024)
-
Evaluation of the Neuroprotective Effect of Organic Selenium Compounds: An in Vitro Model of Alzheimer’s Disease
Biological Trace Element Research (2024)
-
Blood and CSF chemokines in Alzheimer’s disease and mild cognitive impairment: a systematic review and meta-analysis
Alzheimer's Research & Therapy (2023)
-
Plasma extracellular vesicles reveal early molecular differences in amyloid positive patients with early-onset mild cognitive impairment
Journal of Nanobiotechnology (2023)
-
Pharmacological Management of Apathy in Dementia
CNS Drugs (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
