
Abstract
The blind subterranean mole rat Spalax shows a remarkable tolerance to hypoxia, cancer-resistance and longevity. Unravelling the genomic basis of these adaptations will be important for biomedical applications. RNA-Seq gene expression data were obtained from normoxic and hypoxic Spalax and rat liver tissue. Hypoxic Spalax broadly downregulates genes from major liver function pathways. This energy-saving response is likely a crucial adaptation to low oxygen levels. In contrast, the hypoxia-sensitive rat shows massive upregulation of energy metabolism genes. Candidate genes with plausible connections to the mole rat’s phenotype, such as important key genes related to hypoxia-tolerance, DNA damage repair, tumourigenesis and ageing, are substantially higher expressed in Spalax than in rat. Comparative liver transcriptomics highlights the importance of molecular adaptations at the gene regulatory level in Spalax and pinpoints a variety of starting points for subsequent functional studies.
Similar content being viewed by others



Introduction
Much of our knowledge on biological processes and principles, especially in biomedical research, is based on studies in a rather small number of model organisms such as mouse and rat. However, the applicability of standard animal models e.g. for cancer research is limited1,2 and there is a need for a more diverse set of “non-model” organisms showing unusual phenotypes. Two rodents that have attracted attention as emerging models for biomedical research are the blind mole rat Spalax and the naked mole rat Heterocephalus glaber 3; each of which shares a set of unique biological traits highly relevant for human health. Both taxa are tolerant to extreme environmental conditions such as strong hypoxia and reoxygenation. Studying their adaptive mechanisms to prevent hypoxic ischemia or respiratory diseases will be highly interesting. Furthermore, both taxa have extraordinarily long life spans and show strong cancer resistance, making them attractive for ageing and cancer research4,5,6,7,8,9. The less known Spalax, however, has an additional advantage: Spalax belongs to the same rodent superfamily (Muroidea) as mouse and rat, while Heterocephalus has been phylogenetically separated from this clade for more than 70 million years7,10. Thus Spalax allows for a better comparison of biological traits to established rodent models.
The blind mole rats from the Spalax superspecies complex11,12 inhabit self-dug burrows in plains from northern Africa over the Middle East to southwestern Asia. The hypoxia tolerance, which the species complex has evolved during millions of years of subterranean life13, enables them to tolerate oxygen levels as low as 3% for up to 14 hours without noticeable detrimental effects, while rats survive such hypoxic stress for only 2–3 hours14. In natural habitats, oxygen levels of 7.2% were measured in Spalax burrows after heavy rainfalls15, confirming the ecological relevance of effective adaptation to strong environmental hypoxia. While the life span of rats is approximately four years, Spalax can reach ages of 20 years and more16. At present Spalax is the only species where no spontaneous tumour formation has been detected in thousands of individuals studied in the past 40 years. With only a few exceptions, carcinogenic substances were not able to induce malignancies, while all similarly treated mice and rats developed tumours6. Moreover, fibroblasts of Spalax secrete substance/s, yet to be identified, which lead to the death of cancer cells from different species, including a wide range of human cancer cells6. Interferon-β secretion has been speculated to be involved in Spalax cancer resistance17, but these results have been criticized6. While in Heterocephalus the longevity and linked cancer resistance have been ascribed at least partially to the species’ eusociality18, these traits in the solitary living Spalax have probably evolved by convergence using different molecular mechanisms. Genome sequencing of Spalax recently unravelled interesting genomic features, which potentially form the basis of some of the observed adaptive traits7. Observations included high rates of DNA editing, reduced chromosomal rearrangements, adaptation to darkness by pseudogenisation of visual system genes, adaptation to hypoxic/hypercapnic conditions by positive selection in respiratory proteins and reduced sensitivity to hypercapnia-induced acid pain7. A hypoxia-induced upregulation of SINE B1 and LINE-1 retrotransposon expression was noticed, which could possibly play a role in the cancer resistance of Spalax: In the presence of a naturally occurring substitution in the TP53 tumour suppressor gene19, the TRAIN (‘transcription of repeats activates interferon’) system is activated, resulting in diminished apoptosis but increased necrosis in Spalax, as well as an increase in cell cycle arrest/DNA repair7,19. Further insight into the molecular basis of Spalax’ adaptations has been gained on the transcriptome level by microarray studies of brain16 and muscle tissues16,20, inferring differentially expressed genes at hypoxia and normoxia. In these experiments, biological processes and pathways altered by hypoxia in the gene expression of Spalax included angiogenesis, apoptosis, cancer, embryonic/sexual development, epidermal growth factor receptor binding, coordinated suppression and activation of distinct groups of transcription factors, membrane receptors and oxidative stress management. In a recent RNA-Seq study on brain tissue, normoxic mRNA levels of genes associated with DNA damage repair were expressed significantly higher in Spalax when compared to rat, while genes associated with bioenergetics were expressed at lower levels21.
To achieve a more comprehensive understanding of Spalax’ stress adaptation phenotype, it is mandatory to study regulated genes and pathways active in additional organs of the body. The liver is indispensable to life in all vertebrates, and its response to hypoxia is crucial for the organism, as the organ exerts highly oxygen-dependent functions such as glycolysis/gluconeogenesis and the detoxification of harmful endogenous or xenobiotic substances22. Accordingly, hypoxia can cause severe pathological phenotypes in the liver of non-adapted organisms, including inflammation and carcinogenesis23. Comparing Heterocephalus liver transcriptomes to those of mouse, an increased expression of DNA repair signalling genes has recently been observed4. In Spalax however, our knowledge of differential gene expression in liver tissue is very limited so far. Respiratory proteins and proteins of the anti-oxidant defence indeed suggested the existence of species-specific, possibly adaptive gene regulatory expression patterns in Spalax liver24,25, confirming the need for a transcriptome-wide analysis. The gene expression profiles of normoxic and hypoxic livers from Spalax and rat specimens were therefore investigated by Illumina-based mRNA-Seq. The aim of the present study was to reveal Spalax-specific gene-regulatory strategies (as opposed to rat) to cope with strong, acute hypoxia in liver tissue. In addition we aimed to identify genes which are differentially expressed constitutively between Spalax and rat liver tissue under normoxic, non-stress conditions. These data may help unravel the genetic basis of traits associated with cancer-resistance and longevity in Spalax.
Results
Sequencing and raw data processing
In order to infer mRNA gene expression, we performed Illumina-based deep-sequencing of transcriptomes, producing around 257 million sequence reads for Spalax and 327 million reads for rat. In both species, on average 93% of the reads passed quality processing, resulting in a total of 240 million cleaned Spalax reads and 303 million cleaned reads from rat. On average, 97% of the 121 million Heterocephalus reads passed the same quality processing pipeline, resulting in 117 million cleaned reads for Heterocephalus and 660 million cleaned reads overall (Additional Table 1).
Read-mapping and differential gene expression analysis
Expression levels of individual genes were obtained by bioinformatical mapping of the cDNA-derived sequence reads to annotated reference genomes, followed by read-counting. Normalized gene expression was expressed as FPKM values ( = fragments per kilobase of exon sequence per million of mapped fragments). Thereby, we could detect expression in at least one of the replicates for 14,380 genes in Spalax and 15,033 genes in rat (Supplementary dataset 1). Correlation of the FPKM values between the two species under normoxia was moderate with a coefficient of determination of 0.69. Under normoxia, 9,173 genes were expressed in all three rodent species (Additional Fig. 1). Among the top 20 most highly expressed genes under normoxic conditions, twelve were found in Spalax and rat, with the serum transporter transthyretin (TTR) being the highest expressed gene in both species (Supplementary dataset 2). Noteworthy, n = 1227 genes showed at least a 10-fold higher expression in Spalax than in rat, while only 461 genes were >10-fold higher expressed in rat compared to Spalax. This quantitative relation did not change under hypoxia.

Differential gene expression in normoxia versus hypoxia measured in Spalax (A) and rat (B) The scatter-plot shows FPKM values for each measured gene (represented by a dot). Blue dots indicate genes not differentially regulated under hypoxic stress. Red dots represent genes with a significant gene expression change under hypoxia. A Cuffdiff q-value of 0.05 was set as the threshold. Note the logarithmic scales.
The gene FPKM values for normoxic and hypoxic conditions correlated highly within the species (coefficient of determination of 0.89 in Spalax and 0.88 in rat across all genes) ((Supplementary dataset 1). We found 1,722 (12%) genes to be differentially expressed in Spalax between normoxic and hypoxic conditions, and 4,046 (26.9%) in rat (Fig. 1). In both species roughly three quarters (72.3% in Spalax, 74.8% in rat) of differentially expressed genes were up- and one quarter downregulated in response to hypoxia. In the Heterocephalus liver transcriptome data, 17,586 genes showed expression in at least one sample. Due to missing replicates in this taxon, we only could detect significance of hypoxia-induced differential expression for 39 genes. General pattern of hypoxia-induced gene regulation for all three rodents were highly species-specific on the transcriptome-wide scale and suggested only a slightly higher similarity in expression response between Spalax and rat, possibly reflecting their closer phylogenetic relationship (Fig. 2A).

Gene expression heatmaps on a global scale (A) and for genes specifically involved in carcinogenesis, ageing and DNA repair (B) (A): Differential gene expression (log2-transformed fold changes) under hypoxia across the whole transcriptome. Red colour indicates up-, blue colour downregulation after hypoxic exposure. Species from the left to the right are: Heterocephalus, rat, Spalax. Note that the hypoxia response is globally different between the three species, with slightly more similarity between the phylogenetically closer related rat and Spalax. B: Level of transcriptional gene expression in genes involved in cancer, ageing and DNA repair processes (see text) at normoxia. The hotter the colour, the higher the gene expression value. The bar left to the species columns shows membership to the respective functional classes of genes. Species from the left to the right: rat, Heterocephalus, Spalax. The heat map pattern suggests a closer similarity of the two hypoxia-tolerant rodents. Note that the Heterocephalus data must be interpreted with caution since they originated from a single published dataset using a different hypoxia regime, thus prohibiting statistical verification.
Pathway analysis of the hypoxia response
The genes differentially regulated by induction of hypoxic stress were functionally annotated by assignment of gene ontology (GO) designations. Amongst those differentially regulated genes, we could identify several GO terms that were significantly enriched. Spalax and rat showed similar numbers of enriched GO terms for their genes upregulated under hypoxia (Table 1). In rat, the overall pattern of linked GO terms was represented by six clusters (Fig. 3). These clusters comprised GO terms associated with ‘positive regulation of metabolism’, ‘energy metabolism’, ‘response to hypoxia’, ‘structure/organ development’, ‘positive & negative regulation of apoptosis’ and ‘response to hormone stimulus’ (Fig. 3). The clusters ‘structure/organ development’, ‘response to hormone stimulus’ and ‘response to hypoxia´ also emerged in the graph of GO terms for Spalax (Fig. 4), the latter however not reaching statistical significance. In addition, Spalax showed GO term clusters linked to ‘intracellular signalling’, ‘negative regulation of apoptosis’, ‘cellular processes’ and ‘gene expression/transcription’, with the latter being the most prominent in terms of quantity. Of note, both species showed hardly any GO terms which were significantly enriched amongst the downregulated genes (Table 1). Some GO terms linked to ‘chemical sensory perception’ were underrepresented in the downregulated genes of Spalax and rat, and a few GO terms linked to ‘lipid biosynthetic processes’ were overrepresented in the genes only downregulated in rat.

GO term cluster analysis for genes upregulated under hypoxia in rat Shown are GO terms from the category ‘Biological Process’ which show signs of significant enrichment amongst genes that are upregulated in rat under hypoxia, and their closest neighbours. Circles indicate clusters of GO terms linked to certain broader complexes. A darker colour represents a higher level of significance of enrichment.

GO term cluster analysis for genes upregulated under hypoxia in Spalax Shown are GO terms from the category ‘Biological Process’ which show signs of significant enrichment amongst genes that are upregulated in Spalax under hypoxia, and their closest neighbours. Circles indicate clusters of GO terms linked to certain broader complexes. A darker colour represents a higher level of significance of enrichment.
KEGG pathway enrichment analysis of the hypoxia response revealed striking interspecific differences, mostly being associated with metabolism and signalling processes (Table 2). In the subset of rat hypoxia-upregulated genes, the term ‘metabolic pathway’ showed highest significance, followed by the related pathways ‘glycolysis/gluconeogenesis’ and ‘pentose phosphate pathway’. None of these was listed amongst the significantly enriched pathways in Spalax. In hypoxia-downregulated genes of Spalax, two of the most significant pathways were associated with cytochrome P450 activity. KEGG pathways enriched with upregulated genes in Spalax included ‘MAPK signaling’, ‘pathways in cancer’ and ‘circadian rhythm’, all of which are linked to cell cycle regulation (Table 2).
Species differences in constitutive gene expression associated with cancer, ageing, DNA repair and hypoxia
The GO analysis of the many genes more abundantly expressed in Spalax vs. rat under normoxia (i.e. constitutively without hypoxic stress exposure) yielded highly significant terms, which were all broadly associated with general cell metabolism (Supplementary dataset 3), thus making it difficult to infer more specific functional hypotheses. Genes higher expressed in rat were also mostly associated with the GO term ‘metabolism’, but at lower p-value significances. Interestingly, however, we detected a number of specific GO terms in the list of genes more abundantly expressed in Spalax (i.e. DNA damage, cell cycle, apoptosis) that were not enriched amongst genes expressed to a higher extent in rat. With the adaptive phenotype of Spalax in mind, this led us to specifically compare normoxic expression values of genes potentially involved in DNA repair, carcinogenesis and ageing between the rodent species. Broadly visualizing the strength of constitutive, normoxic expression of these functional gene categories, a high similarity between the two phylogenetically distant mole rats emerged from the heat-map picture (Fig. 2B). This similarity reflects the general finding that the majority of genes were expressed at higher transcript levels in both, Spalax and Heterocephalus, as opposed to rat (see below). Again have in mind that the Heterocephalus transcriptome data lacked replicates, so that interpretations below must be viewed with a certain caution.
Investigating functional gene categories, the majority of candidate oncogenes (15 of 24 genes; all having FPKM >0.1 in either species) were transcribed at ≥2-fold higher levels in Spalax than in rat at normoxia. Only two genes were expressed 2-fold higher in rat than in Spalax (Supplementary dataset 4). Only for one of the 15 genes (MSH2, a p53-targeted DNA repair gene), the augmented normoxic expression in Spalax was compensated for in rat by hypoxia-induced upregulation. A tendency towards increased normoxic expression in Heterocephalus was noted for 8 of the 15 Spalax cases. Out of 107 expressed candidate tumour-suppressors genes, 49 showed significant 2-fold higher mRNA levels in Spalax liver, while only 12 were higher expressed in rat (Supplementary dataset 4). In 4 genes (GADD45B, BHMT, SDS, OTC), rat compensated the lower normoxic expression by its hypoxia response. Of the 49 genes higher expressed in Spalax, 34 showed the same tendency in Heterocephalus.
Amongst genes linked to the ageing process, 28 of 48 were expressed at >2-fold higher levels in normoxic Spalax liver compared to rat (Supplementary dataset 5). Of those 28, 17 also showed the tendency for higher normoxic expression in Heterocephalus. Ten of these genes, namely BUB3, ATR, A2M, HELLS, WRN, CISD2, CETP, SLC13A1, AGTR1 and GH1, were expressed more than 10-fold higher in normoxic Spalax than in rat. Not a single gene was overexpressed to such high extent in rat, where only five genes (MSRA, ARNTL, PLAU, ADCY5, FGF21) had FPKM values at least 2-fold higher than their Spalax counterparts. Interestingly, expression of the longevity-associated fibroblast growth factor 21 gene (FGF21) was approximately 10x lower in Spalax than rat; however, FGF21 was 123-fold upregulated in Spalax by hypoxia, thereby out-numbering rat mRNA levels by 10-fold.
In our list of 167 genes associated with DNA damage and repair, 103 showed >2-fold higher expression in normoxic Spalax liver than in rat, and 19 genes were higher expressed in rat than Spalax (Supplementary dataset 6). Of those 103, 67 also showed elevated FPKM values in Heterocephalus. The remarkable number of 29 genes had >10-fold higher FPKM values in Spalax versus rat. Of special note, 18 genes from the Fanconi anemia (FA) DNA repair pathway were at least 2-fold and 12 genes even >10-fold upregulated in normoxic Spalax liver versus rat (labelled in Supplementary dataset 6). Involvement of this crucial pathway clearly points at differences amongst the two rodents in dealing with DNA damage and its repair. Additionally, Rae et al.4 have published a list of 34 DNA damage and repair genes, which showed significantly elevated expression in Heterocephalus liver compared to mouse. For 31 of those, we have obtained FPKM expression values from both Spalax and rat, and detected that 21 genes were >2-fold higher expressed in Spalax than rat.
Of n = 6 genes from the FOXO3 cell survival pathway, which were discussed with regard to their hypoxia-regulated differential expression in Spalax muscle (comp. Fig. 10 in Ref.16), 5 also showed hypoxia-regulation in Spalax liver (Supplementary dataset 7). From our extended list of hypoxia-related genes (n = 48; Supplementary dataset 7), 24 genes showed >2-fold and eight >10-fold higher FPKM values in normoxic Spalax liver than in rat, while 5 genes showed the opposite. Inspecting individual genes of known importance in hypoxia response, HIF1A showed a mild (2.4-fold) hypoxic transcriptional induction in Spalax liver and about equivalent normoxic FPKM levels in Spalax and rat. Spalax liver HIF2A mRNA levels exceeded those of Spalax HIF1A (9-fold) and those of rat HIF2A (12-fold). Vascular endothelial growth factor (VEGFA), a master factor of angiogenesis, was only slightly transcriptionally activated in Spalax and rat liver (1.5 and 1.8-fold). However, the anti-angiogenic factor TSP1 showed 7-fold hypoxic upregulation only in Spalax.
An RNA-Seq study comparing liver tissue of mice and Heterocephalus reported the strongly increased expression of a set of mitochondrial and oxidation reduction genes in Heterocephalus 26. Amongst the set of genes tested in this study, alpha2-macroglobulin (A2M), a potential biomarker for ageing, showed a high increase of expression in Heterocephalus. Our data were strikingly in line, with an FPKM = 303 for Spalax A2M, but virtually no A2M transcription in rat (FPKM <0.01) (Supplementary dataset 8). Under hypoxia, A2M transcription was detectable in rat (FPKM = 15), but Spalax still overcompensated this (FPKM = 398).
Validation of the RNA-Seq analysis
We complemented the comprehensive RNA-Seq analysis presented above by a second bioinformatic approach, seeking to eliminate potential biases resulting from the choice of methods. This approach differed in two aspects: (1) To particularly strengthen the interspecies comparison, an alternative bioinformatic pipeline producing very stringently defined Spalax-rat gene orthologs was applied, (2) different read-mapping and statistical validation algorithms, as well as other functional gene/pathway annotation tools were applied. Although these steps reduced the number of genes directly comparable between Spalax and rat to ca. 7,000, the major results of interspecies differential gene expression were confirmed and validated (Additional Note 1, Supplementary datasets 9–10).
In addition, we studied the hypoxia inducibility and interspecies expression differences of a selected set of important candidate genes of different functional categories (A2M, ATR, CISD2, CITED2, FGF21, GNMT, PTEN, TOP2B, VEGFA, WRN, XPA) by quantitative realtime RT-PCR (Additional Note 2). Thereby we could confirm the direction of all relevant gene expressional changes.
In order to account for potential biases in liver gene expression (sex, species, strain, age), we inferred transcriptome-wide FPKM values using public rat, mouse and human RNA-Seq data and compared them to our own data (Supplementary dataset 11). The results indicated that expression ratios, in particular those of females and males, did not exceed values of 2-fold for the vast majority of 200 candidate genes in rat, mouse and humans. Hence, such biases are improbable to substantially affect the interspecific and hypoxia-induced gene expression changes reported in our study.
Signs of positive selection in gene coding sequences
In addition to gene expression changes, Darwinian positive selection for functional sequence changes may cause species-specific adaptations in Spalax genes. The coding sequences of 125 genes (Supplementary dataset 12), which were either classified as universal hypoxia genes showing signs of adaptation in other species27 or as functionally related to DNA repair and ageing, were therefore analysed for sequence signatures of positive selection. Using standard bioinformatical procedures to infer the ratio of non-synonymous to synonymous nucleotide substitutions (ω), none of the genes showed prominent signs of positive selection on the Spalax branch, typically defined as ω ≥ 1 (Supplementary dataset12). However, manual inspection of amino acid sequence alignments revealed several Spalax-specific substitutions, which nevertheless appear interesting and might require further attention. A conspicuous example is given by the protein kinase mTOR, which displays 17 amino acid replacements at positions which are fixed in all other mammals included in our study (Additional Fig. 2). Twelve of these replacement positions are clustered within the FAT (FRAP-ATM-TTRAP) domain, while the rest of the protein is highly conserved.
Discussion
The rationale of the present study was to pinpoint the most important features of molecular adaptation in the hypoxia-tolerant, long-lived and cancer-resistant rodent Spalax, by performing a transcriptome-wide analysis of gene expression in comparison to the phylogenetically related, hypoxia-sensitive biomedical model species rat. We focused on analysing liver gene expression since the liver represents a highly active metabolic organ, crucial for survival, which should readily unveil interspecies differences. Measured RNA levels as a proxy of gene expression can explain two thirds of the protein abundance in mammalian cells28, thus forming an excellent guiding basis for subsequent molecular biological experiments. The main results of our comparative RNA-Seq analysis should be largely independent of technical biases, since they were validated using different ortholog gene annotation pipelines, read-mapping algorithms and tools to identify and functionally interpret differential gene expression. Potential biological biases (e.g. male/female) were also relatively small, compared to the gene expression differences observed between species or treatments.
Analysing differential gene expression in liver after hypoxic exposure of animals, we found more than twice as many genes being upregulated under hypoxia in rat (26.9%) than in Spalax (12%). The O2 level chosen for experimental hypoxia (6% O2) should not have been too mild for Spalax to provoke its full hypoxia stress response, since we applied a value below the lowest O2 content of 7.2%, which was measured experimentally in Spalax burrows in its natural habitat15. An alternative explanation is that, because Spalax is frequently confronted with hypoxia, its hypoxia stress response genes might already be expressed constitutively at levels required to avoid hypoxic damage. Indeed, Spalax showed higher constitutive, normoxic expression levels in many genes, which are functionally related to the investigated traits hypoxia, longevity, and tumorigenesis. The present comprehensive transcriptome comparison thus confirms and extends previous studies on phenotypically relevant candidate genes, which consistently displayed elevated expression in the blind mole rat (e.g. vascular endothelian growth factor14, hypoxia inducible factor and erythropoietin15, ROS defense genes24, globins25). Despite lacking replicate datasets and a different hypoxia regime, a similar trend of constitutively higher mRNA expression was also seen in naked mole rat liver for many genes involved in carcinogenesis, ageing and DNA damage repair. A noteworthy example is the increased expression of alpha-2 macroglobulin (A2M) in both mole rats, as opposed to rat and mouse (discussed below). With all necessary caution, this finding raises the interesting possibility that an augmented expression of genes has evolved convergently in the two distantly related, hypoxia-tolerant rodents. Constitutive, high expression of stress-response genes may confer a selective advantage to species encountering either extreme acute hypoxia (like Spalax at times of flooding in heavy soils) or chronic hypoxia (like Heterocephalus in its crowded eusocial colonies) by saving time and energy to switch-on the stress response. A hypoxia-sensitive species such as rat, which only occasionally encounters short-term mild hypoxia, may instead profit from a dedicated stress-induced gene regulatory response. Different strategies of gene regulation may thus have shaped the adaptive evolution of hypoxia-tolerant and -sensitive rodents. In contrast, our molecular evolutionary analysis of coding sequences revealed a lack of pronounced Darwinian evolution within candidate genes associated with ageing, longevity, and DNA repair and also in genes previously shown to be involved in hypoxia adaptation in humans and fruit flies27. Therefore, the Spalax phenotype might have evolved predominantly by gene regulatory changes, which in turn might have been orchestrated by positively selected key amino acid changes in important transcriptional master regulators like p5319 and the Nrf2-Keap1 complex29. Interestingly, here we report Spalax-specific amino acid replacements in the FAT domain of the mTOR protein (Additional Fig. 2), which is a key player in coupling cancer susceptibility, ageing and energy metabolism30. In addition, mTOR is involved in cell signalling and carcinogenesis31. Gain-of-function-mutations in the mTOR FAT domain cause hyperactivation, leading to increased proliferation of cancer cells and poor prognostic outcomes for patients32. The mTOR pathway in turn controls cell proliferation via activation of the FOXO3 transcription factor33. Indeed we found an hypoxia-induced upregulation of the FOXO3 pathway genes in the Spalax liver transcriptomes, in agreement with published data for Spalax myocytes16. We hypothesize that the increased transcription in the FOXO3 pathway might be caused by the mutated mTOR protein.
A general outcome emerging from our study is that both species, Spalax and rat, react to hypoxia by differential gene expression. However, the hypoxia-adapted Spalax regulates substantially less genes than the rat when temporarily exposed to 6% O2. In both species, the hypoxia-upregulated genes showed many more overrepresented GO terms than the downregulated genes (total gene numbers were approximately equal). Hypoxia thus prompted a strong upregulatory gene response, most probably by hypoxia-inducible transcriptional activators such as HIF134.
In the hypoxia-induced genes of both rodents, GO terms linked to ‘structure/organ development’ and ‘response to hormone stimulus’ were enriched, in line with a generally known stress reaction of many organs. In addition, the enriched term ‘structure/organ development’ with its sub-terms ‘angiogenesis’ and ‘vasculature development’ implicates an initiation of vascularisation, possibly to ensure oxygen supply during hypoxic stress. Stimulation of vascularisation seemed to be more pronounced in rat, suggesting that Spalax livers are already well vascularized, as indeed shown before in Spalax muscle tissue14. Again, in both species GO terms linked to stress were identified. However, only in rat the specific GO cluster ‘response to hypoxia’ was significantly enriched in hypoxia-inducible genes, while in Spalax only the more general term ‘response to stress’ emerged. This is in line with the more pronounced hypoxia response of the rat, as noted before (Fig. 1).
Another enriched GO term cluster among hypoxia-regulated genes of both species was ‘regulation of apoptosis’. In rat, GO terms ‘positive regulation of apoptosis’ as well as ‘negative regulation of apoptosis’ were enriched, while in Spalax only the sub-branch ‘negative regulation of apoptosis’ appeared. This finding is in agreement with the genome-wide analysis of sequence evolution and gene duplication events in the apoptosis pathway of Spalax 7, which suggested a repression of apoptosis in Spalax, possibly to avoid cell loss during physiological stress.
Besides these shared expression patterns of hypoxia-upregulated genes, Spalax and rat also showed species-specific clusters of enriched GO terms. In rat, two major clusters were related to ‘positive regulation of metabolism’ and ‘energy metabolism’. The latter included GO terms like ‘catabolic process’ and ‘saccharide metabolic processes’. At the same time, GO terms linked to ‘lipid biosynthetic processes’ emerged amongst hypoxia-downregulated genes. Together, we conclude that the rat maintains its metabolism under heavy stress by catabolic processes. The KEGG pathway analysis confirmed this hypothesis: The hypoxia-sensitive rat markedly upregulated metabolic pathways including glycolysis/gluconeogenesis, a most critical metabolic function of the liver. Since this is part of a known common reaction to hepatic stress35, the observed gene regulation pattern can be seen as an attempt of a hypoxia-sensitive species to cope with the life-threatening stress of 6% O2. In stark contrast, Spalax showed downregulation of metabolic genes under hypoxia, including those involved in the p450 detoxification system playing a major role in liver. Downregulation of many energy-consuming processes in Spalax under acute hypoxia has previously been shown for brain tissues (see Supplementary Information in Ref7), highlighting the general importance of this strategy. It has well been recognized that a coordinated downregulation of biosynthetic, proliferative and energetic pathways is a hallmark of hypoxia/anoxia-tolerant vertebrates36. By preventing a lethal depletion in ATP, hypometabolism is considered a key adaptation to survive and recover from the lack of O2.
In addition to metabolic changes, Spalax liver genes upregulated under hypoxia were associated with roles in tumourigenesis. We found these genes enriched for GO terms and KEGG pathways linked to intracellular signalling and cell cycle regulation. An example is the FOXO3 pathway, which controls tumourigenesis37 and is moreover associated with longevity38. In parallel, the candidate anti-angiogenic gene TSP1 was induced by hypoxia. Many of these gene regulatory changes in cancer- and angiogenesis-related genes were previously also observed in Spalax muscle and might contribute to a cancer-delimiting phenotype (comp.16). It will thus be important to investigate in detail if and how Spalax modulates these functions and possibly creates resilience to tumour formation as a by-product of its species-specific hypoxia response. Several additional candidate genes require special attention in this context:
-
BUB3, transcribed 28-fold higher in Spalax liver than in rat, is essential for the spindle assembly checkpoint in higher eukaryotes. While a complete knockout of BUB3 is lethal to mice already in utero, haplo-insufficiency leads to chromosome missegregation and increased susceptibility to chemical-induced tumourigenesis39. BUB3 further cooperates with the mitotic checkpoint protein RAE1. Mice double haplo-insufficient for BUB3 and RAE1, but not single haplo-insufficient animals, produce a premature senescence phenotype40. Interestingly, also RAE1 expression is augmented in Spalax versus rat (5.8-fold), and also Heterocephalus shows higher expression levels than rat for both genes (9.3-fold for BUB3 and 4.1-fold for RAE1). Together, this indicates a potential impact of the BUB3/RAE1-complex on longevity and cancer resistance in both mole rat species.
-
While expressed at low levels in normoxic Spalax liver, transcription of the liver-secreted hormone fibroblast growth factor 21 (FGF21) increased 123-fold under hypoxia. In humans, FGF21 is upregulated during fasting and induces oxidative metabolism in peripheral tissues. Transgenic mice which overexpress FGF21 showed increased lifespans41. FGF21 might thus be involved in linking the ecologically selected hypoxia response to the longevity phenotype.
-
Another age-related gene, which is 12-fold overexpressed under normoxia in Spalax liver compared to rat, is the CDGSH Iron Sulfur Domain 2 (CISD2), a protein involved in cellular calcium homeostasis. Mutations of human CISD2 cause Wolfram syndrome 2, a neurodegenerative disease associated with diabetes mellitus, optic atrophy and a decreased lifespan (OMIM 222300). CISD2-null mice showed early senescence and shortened life span compared to wildtype mice42. Interestingly, transgenic overexpression of CISD2 in mice increased lifespan, possibly by an interconnection between calcium homeostasis and autophagy43.
-
Longevity is plausibly associated with the activity of DNA damage repair pathways, which ensure genome integrity. Recent studies implied that long-lived species such as humans or Heterocephalus feature higher expression rates of DNA damage repair genes than short-lived organisms such as mice4. Brain tissue of the long-lived Spalax was also shown to display increased transcription of DNA damage and repair genes in comparison to rat21. We observed the same tendency in Spalax liver. For example, the DNA glycosylase NEIL2 (involved in base excision repair) and the Xeroderma pigmentosum-associated XPA (a damage-specific DNA binding protein involved in nucleotide excision repair) are about 56- and 59-fold higher transcribed in normoxic Spalax liver than in rat. Overexpression of XPA in human cells increased their resistance to ultraviolet radiation-induced DNA damage44,45. However, more transgenic animal models or cell cultures with overexpressed repair genes, mimicking the natural gene expression pattern of mole rats, will be necessary to prove a direct correlation between increased levels of DNA damage and repair proteins and the longevity phenotype.
-
Several genes associated with the Fanconi Anemia (FA) DNA repair pathway are expressed at much higher levels in Spalax than in rat (Fig. 5), including genes which encode the FA core complex and its main upstream activator ATR46. This essential pathway orchestrates recombination and repair processes which ensure chromosomal stability, while mutations in FA genes result in increased levels of chromosomal rearrangements and pre-dispose patients to cancer47. An intact FA pathway is required to prevent hypoxia-induced DNA damage in human cells in vitro 48. Overexpression of FA gene A (FANCA) decreased the susceptibility of human breast cancer cells to cross-linking agents49, and overexpression of a group C gene (FAC) protected mouse hematopoietic stem cells from apoptosis50. In line with increased expression of FA genes that could lead to an increased activity of DNA repair mechanisms, the Spalax genome also shows a reduced rate of chromosomal rearrangements7.
Figure 5 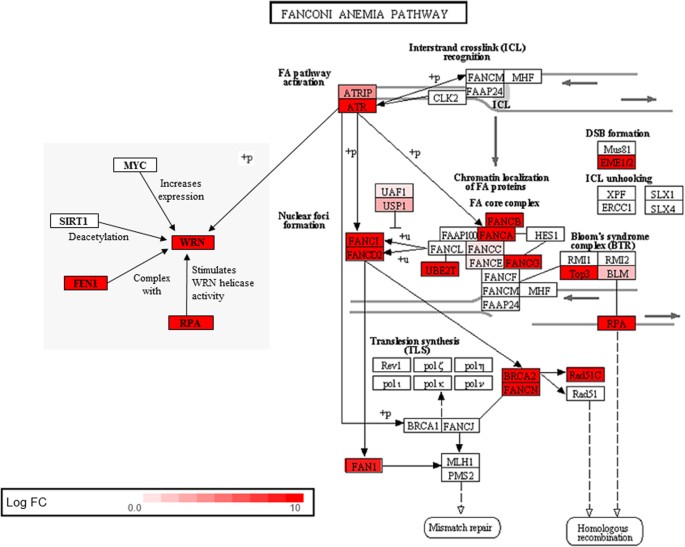
DNA damage and repair genes upregulated in Spalax liver as compared to rat. N = 18 genes from the Fanconi Anemia (FA) pathway were transcriptionally upregulated in normoxic Spalax liver, as indicated by the colour chart (prepared using Advaita Bio’s iPathwayGuide; http://www.advaitabio.com/ipathwayguide.html). Note that both, the major activator of this pathway, ATR, and its downstream interacting partner, the helicase WRN are overexpressed on the RNA level in Spalax versus rat liver. The same tendency was shown by RPA, which stimulates the helicase and exonuclease activity of WRN, and by FEN1, which complexes WRN to adjust stalled replication forks.

Interestingly, we also observed increased transcription in Spalax liver of the Werner syndrome helicase gene WRN, which encodes an interacting partner protein of the FA key activator ATR and functions as a helicase and exonuclease in many DNA repair processes51,52. Mutations in WRN cause the Werner syndrome, which is associated with premature ageing and an increased cancer susceptibility53. We assume that elevated expression levels of ATR and WRN might be linked to longevity and cancer resistance in Spalax, given that Heterocephalus also showed higher expression of these two genes. Recently, overexpression of ATR and WRN has been implicated in mediating autophagy, a cell-maintaining process known to be compromised in normal and pathological ageing54,55.
• The Alpha-2 macroglobulin (A2M) gene encodes a serum protease inhibitor produced by the liver. It has been found overexpressed in Heterocephalus compared to mouse26. Moreover, Heterocephalus plasma was described to contain higher A2M protein levels than human plasma56. Interestingly, our data also show increased A2M gene expression in Spalax compared to rat. A2M has been shown to have strong tumour-suppressor properties in astrocytoma cells, possibly by interfering with ß-catenin signalling57, and it was recently used to mediate tumour antigen presentation in cancer immunotherapy58. The convergence of high A2M expression in both mole rats highlights the protein as another promising candidate to further study its role in cancer resistance. In the presence of increased expression levels of highly potent tumour-suppressor and DNA repair genes, the observed increased expression of a subset of oncogenes in Spalax may be overcompensated for, thus leading to an overall suppression of tumourigenesis in this species.
We conclude that Spalax uses a completely different gene expression strategy to cope with hypoxic stress than rat in a vital tissue like liver. While the hypoxia-sensitive rat increases its cellular energy metabolism under hypoxia, a reaction possibly related to stress evasion, Spalax instead downregulates the majority of metabolic pathways. This latter response is indicative for the transition to a hypometabolic state and likely forms a crucial adaptation of Spalax, enabling its cells to survive strong, acute hypoxia without suffering from severe damage. Under hypoxia, Spalax differentially expresses genes involved in cell cycle regulation, hypothetically resulting in a tumourigenesis-controlling cellular state. A number of genes plausibly associated with cancer resistance, longevity and DNA damage repair are expressed at markedly higher levels in normoxic Spalax liver compared to rat, potentially ensuring constitutive protective effects. Altogether, transcriptome comparisons of hypoxia-tolerant versus -sensitive tissues of blind mole rats reveal multiple promising starting points for subsequent functional experiments in biomedical research and evolutionary genomics.
Methods
Animals, experiments and sequencing
All protocols of animal capturing, maintenance, experimental procedures and sacrificing were according to the regulations of the Israel Nature and Park Authority, Science and Conservation Unit, and have been approved by the Haifa University Committee on Animal Care. N = 6 Spalax galili (the northern subspecies of the four residing in Israel; female individuals captured from basalt, heavy soil) were used. The Spalax specimen were caught in the field and kept under routine veterinary scrutiny in the Animal House of the Institute of Evolution at Haifa University until approximately 4 years old (adults, 150–250 g). N = 6 Rattus norvegicus specimen (Sprague Dawley, 4 month old males, 350–400 g) were used. The age of the two species roughly corresponds given their different overall life expectancy. For the experiment, n = 3 specimen of Spalax and rats were kept under normoxic (21% O2) and n = 3 under hypoxic (6% O2) conditions for six hours each. This regime was chosen to ensure that a hypoxia response was elicited. No time-series of hypoxic gene expression exist for Spalax liver, but mRNA quantification in Spalax kidney indicated that HIF1a mRNA markedly increased after 4 hours (6% O2) and stayed high until 24 hours (10% O2)59. Evidence for a successful hypoxia response is indicated by upregulation of known HIF1a target genes (see Supplementary dataset 7).
For hypoxia exposure, we used 70 × 70 × 50 cm chambers divided into separate cells and a gas flow rate of 3.5 l/min. All animals were adults of similar weight (~150 g). After the six hours of exposure, animals were immediately sacrificed by injection with Ketaset CIII (Fort Dodge, USA) at 5 mg/kg of body weight and their tissues harvested and transferred to liquid nitrogen. Rat specimen were kept and processed under the same conditions. RNA samples from the liver right lobe of twelve animals was extracted with Qiagen RNeasy Mini Kit (Qiagen, Germany) including an on-column DNase I digestion and quality checked with a RNA nanochip on an Agilent 2100 Bioanalyzer (Agilent Technologies, CA, USA). Libraries for high-throughput cDNA sequencing (RNA-Seq) were constructed using the TrueSeq RNA Sample Prep Kit v.2 (Illumina, CA, USA) tagged with different multiplex indices (StarSEQ, Mainz, Germany). All six libraries were sequenced as 100 bp paired-end reads on an Illumina HiSeq 2500 instrument (Institute of Organismic and Molecular Evolution, University of Mainz, Germany).
Raw data processing
All reads from the 12 samples were processed prior to analyses using the FASTX toolkit 0.0.13 (http://hannonlab.cshl.edu/fastx_toolkit) with the following parameters: 15 bp cut from the 5′-end, removal of universal and index adapters, trimming of all positions at the reads’ ends with quality scores below 20, removal of all reads with less than 80% positions with a quality score of 20 or higher. All datasets were quality-checked with the FastQC software (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) multiple times to assure optimal results. For further interspecific comparison we downloaded RNA-Seq-derived liver transcriptome sequences of Heterocephalus glaber from NCBI’s Sequence Read Archive (SRA). One dataset was from an animal kept under normoxic conditions (SRA accession number SRR306395) and one from a hypoxia exposed individual (8% O2 for one week, SRR306404). The reads from these two datasets were processed in the same way as described above.
Mapping and differential gene expression analysis
Reference genome sequences were downloaded from public databases, using version 1.0 of Spalax’ genome from NCBI’s genome database and Ensemble release 72 for the rat genome. Spalax and rat RNA-Seq reads were mapped and analysed for differential expression (DE) using the TopHat-Cufflinks-Cuffmerge-Cuffquant-Cuffdiff-pipeline60. TopHat 2.0.1161 with Bowtie2 2.2.162 was used for mapping and Cufflinks 2.1.163 for DE analysis. A q-value of <0.05 was set as threshold for DE between normoxic and hypoxic triplicates for Spalax and rat FPKM values, respectively. Additionally, we calculated FPKM-fold-changes from normoxia to hypoxia, log2-transformed and then fed them into the heatmap.2-function from the R gplots package in RStudio 0.98.1060 (http://www.rstudio.com/).
Gene ontology and pathway analyses
We used the vocabulary of the Gene Ontology (GO) consortium64 for annotation of genes, pathways and processes regulated under hypoxic conditions. GO enrichment analyses in the category ‘Biological Process’ were performed using the BiNGO 2.44 plugin65 for Cytoscape 3.1.166, applying default parameters. A significance level of 0.01 was set as threshold for the Benjamini-Hochberg false-discovery rate-corrected hypergeometric test and R. norvegicus as the annotation background. Input lists consisted of significantly regulated genes (q-value < 0.01) for each of the two species, split into up- and downregulated genes. Each group of genes was checked for over- and under-representation of GO terms separately.
Additionally, we performed a search for KEGG pathways67,68 enriched with genes regulated under hypoxic conditions. For this task we used the webserver version of WebGestalt69,70 with R. norvegicus as the annotation background and a significance level of 0.01 for the Benjamini-Hochberg adjusted p-value.
Compilation of genes related to cancer, ageing, DNA damage and repair and hypoxia
To compare expression differences between Spalax and rat in genes known to be differentially expressed in liver cancer, we created two sets of genes using the RNA Seq Atlas resource (http://medicalgenomics.org/rna_seq_atlas 71). The first list consisted of 32 genes consistently upregulated in 15 different liver cancer cell lines, representing candidate oncogenes. The second list consisted of 162 genes consistently downregulated in these 15 different liver cancer cell lines, i.e. candidate tumour-suppressor genes. We then compared the interspecific differences in the gene expression levels under normoxia and their regulation under hypoxia. A list of 54 genes with published links to the ageing process in humans (as studied by mammalian models) was downloaded from the database GenAge72,73 and complemented with literature knowledge to identify ageing-related genes differentially expressed between Spalax and rat. Since DNA repair mechanisms are important players in both, carcinogenesis and ageing, we additionally checked a list of relevant genes (n = 202 genes) derived from www.sciencepark.mdanderson.org and extracted from literature. A list of hypoxia-related genes (n = 60) was compiled to include the hypoxia-regulated FOXO3A pathway (regulated in Spalax muscle16), a list of genes involved in hypoxia adaptation in humans and Drosophila 27 and known hypoxia-responsive genes from literature. Besides manual inspection, the log2-transformed FPKM values under normoxia from the listed genes were fed into RStudio as described above with row scaling applied.
Search for signs of positive selection
To additionally investigate the impact of positive selection within gene coding sequences as a candidate mechanism for the peculiar adaptations of Spalax, a subset of 125 ageing-, cancer- and hypoxia-associated genes (Supplementary dataset12) was screened on the sequence level in a phylogenetic context. Orthologous transcript sequences of candidate genes from Spalax, other rodents and outgroup mammals were downloaded from public databases and aligned using the MUSCLE algorithm74 implemented in MEGA 5.0175 with default settings. Alignments were trimmed to the coding sequence, manually inspected and corrected, and then used to carry out branch-specific tests of positive selection implemented in codeml as part of the program package PAML 4.9a76,77. Likelihood values from free ratio model tests were compared to likelihood values from one-ratio model tests to identify significant signs of positive selection on the Spalax branch. The underlying rodent phylogeny was obtained from Voloch et al.78.
Data availability
The datasets supporting the conclusions of this article are available in the European Nucleotide Archive ENA under the study accession number PRJEB17935.
References
Anisimov, V. N., Ukraintseva, S. V. & Yashin, A. I. Cancer in rodents: does it tell us about cancer in humans? Nat Rev Cancer 5, 807–819 (2005).
Cheon, D.-J. & Orsulic, S. Mouse models of cancer. Annu Rev Pathol Mech 6, 95–119 (2011).
Azpurua, J. & Seluanov, A. Long-lived cancer-resistant rodents as new model species for cancer research. Front Genet 3, 319 (2013).
MacRae, S. L. et al. DNA repair in species with extreme lifespan differences. Aging 7, 1171–1184 (2016).
Nasser, N. J. et al. Alternatively spliced Spalax heparanase inhibits extracellular matrix degradation, tumor growth, and metastasis. Proc Natl Acad Sci USA 106, 2253–2258 (2009).
Manov, I. et al. Pronounced cancer resistance in a subterranean rodent, the blind mole-rat, Spalax: in vivo and in vitro evidence. BMC Biol 11, 91 (2013).
Fang, X. et al. Genome-wide adaptive complexes to underground stresses in blind mole rats Spalax. Nat Commun 5, 3966 (2014).
Lewis, S. H., Salmela, H. & Obbard, D. J. Duplication and diversification of Dipteran Argonaute genes, and the evolutionary divergence of Piwi and Aubergine. Genome Biol Evol 8, 507–518 (2016).
Miyawaki, S. et al. Tumour resistance in induced pluripotent stem cells derived from naked mole-rats. Nat Commun 7, 11471 (2016).
Fang, X. et al. Adaptations to a Subterranean Environment and Longevity Revealed by the Analysis of Mole Rat Genomes. Cell Reports, (2014).
Wahrman, J., Goitein, R. & Nevo, E. Mole rat Spalax: evolutionary significance of chromosome variation. Science 164, 82–84 (1969).
Nevo, E., Ivanitskaya, E. & Beiles, A. Adaptive radiation of blind subterranean mole rats: naming and revisiting the four sibling species of the Spalax ehrenbergi superspecies in Israel: Spalax galili (2n = 52), S. golani (2n = 54), S. carmeli (2n = 58) and S. judaei (2n = 60). (Backhuys Publishers, Leiden, 2001).
Lacey, E. A., Patton, J. L. & Cameron, G. N. Life underground: the biology of subterranean rodents. (University of Chicago Press, 2000).
Avivi, A. et al. Increased blood vessel density provides the mole rat physiological tolerance to its hypoxic subterranean habitat. FASEB J 19, 1314–1316 (2005).
Shams, I., Avivi, A. & Nevo, E. Oxygen and carbon dioxide fluctuations in burrows of subterranean blind mole rats indicate tolerance to hypoxic–hypercapnic stresses. Comp Biochem Physiol, A: Mol Integr Physiol 142, 376–382 (2005).
Malik, A. et al. Transcriptome analysis of the Spalax hypoxia survival response includes suppression of apoptosis and tight control of angiogenesis. BMC Genomics 13, 615 (2012).
Gorbunova, V. et al. Cancer resistance in the blind mole rat is mediated by concerted necrotic cell death mechanism. Proc Natl Acad Sci USA 109, 19392–19396 (2012).
Williams, S. A. & Shattuck, M. R. Ecology, longevity and naked mole-rats: confounding effects of sociality? Proc R Soc London, B 282, (2015).
Avivi, A. et al. P53 in blind subterranean mole rats: loss-of-function versus gain-of-function activities on newly cloned Spalax target genes. Oncogene 26, 2507–2512 (2007).
Avivi, A., Brodsky, L., Nevo, E. & Band, M. R. Differential expression profiling of the blind subterranean mole rat Spalax ehrenbergi superspecies: bioprospecting for hypoxia tolerance. Physiol Genomics 27, 54–64 (2006).
Malik, A. et al. Genome maintenance and bioenergetics of the long-lived hypoxia-tolerant and cancer-resistant blind mole rat, Spalax: a cross-species analysis of brain transcriptome. Scientific Reports 6, 38624 (2016).
Baze, M. M., Schlauch, K. & Hayes, J. P. Gene expression of the liver in response to chronic hypoxia. Physiol Genomics 41, 275–288 (2010).
Rosmorduc, O. & Housset, C. Hypoxia: a link between fibrogenesis, angiogenesis, and carcinogenesis in liver disease. Semin Liver Dis 30, 258–270 (2010).
Schülke, S. et al. Living with stress: Regulation of antioxidant defense genes in the subterranean, hypoxia-tolerant mole rat. Spalax. Gene 500, 199–206 (2012).
Avivi, A. et al. Neuroglobin, cytoglobin, and myoglobin contribute to hypoxia adaptation of the subterranean mole rat Spalax. Proc Natl Acad Sci USA 107, 21570–21575 (2010).
Yu, C. et al. RNA Sequencing Reveals Differential Expression of Mitochondrial and Oxidation Reduction Genes in the Long-Lived Naked Mole-Rat When Compared to Mice. PLoS One 6, e26729 (2011).
Jha, A. R. et al. Shared genetic signals of hypoxia adaptation in Drosophila and in high-altitude human populations. Mol Biol Evol 33, 501–517 (2015).
Vogel, C. et al. Sequence signatures and mRNA concentration can explain two-thirds of protein abundance variation in a human cell line. Mol Syst Biol 6 (2010).
Schmidt, H., Hangmann, J., Shams, I., Avivi, A. & Hankeln, T. Molecular evolution of antioxidant and hypoxia response in long-lived, cancer-resistant blind mole rats: The Nrf2-Keap1 pathway. Gene 577, 293–298 (2016).
Zoncu, R., Efeyan, A. & Sabatini, D. M. mTOR: from growth signal integration to cancer, diabetes and ageing. Nat Rev Mol Cell Biol 12, 21–35 (2011).
Guertin, D. A. & Sabatini, D. M. Defining the role of mTOR in cancer. Cancer Cell 12, 9–22 (2007).
Ghosh, A. P. et al. Point mutations of the mTOR-RHEB pathway in renal cell carcinoma. Oncotarget 6, 17895 (2015).
Mori, S. et al. The mTOR pathway controls cell proliferation by regulating the FoxO3a transcription factor via SGK1 kinase. PloS One 9, e88891 (2014).
Semenza, G. L. Hypoxia-inducible factor 1 and cardiovascular disease. Annu Rev Physiol 76, 39–56 (2014).
Orman, M. A., Berthiaume, F., Androulakis, I. P. & Ierapetritou, M. G. Pathway analysis of liver metabolism under stressed condition. J Theor Biol 272, 131–140 (2011).
Hochachka, P., Buck, L., Doll, C. & Land, S. Unifying theory of hypoxia tolerance: molecular/metabolic defense and rescue mechanisms for surviving oxygen lack. Proc Natl Acad Sci USA 93, 9493–9498 (1996).
Zhang, Y., Gan, B., Liu, D. & Paik, J.-h FoxO family members in cancer. Cancer Biol Ther 12, 253–259 (2011).
Shimokawa, I. et al. The life-extending effect of dietary restriction requires Foxo3 in mice. Aging Cell 14, 707–709 (2015).
Babu, J. R. et al. Rae1 is an essential mitotic checkpoint regulator that cooperates with Bub3 to prevent chromosome missegregation. J Cell Biol 160, 341–353 (2003).
Baker, D. J. et al. Early aging-associated phenotypes in Bub3/Rae1 haploinsufficient mice. J Cell Biol 172, 529–540 (2006).
McCarty, M. F. Practical prospects for boosting hepatic production of the “pro-longevity” hormone FGF21. Horm Mol Biol Clin Investig 30, (2015).
Chen, Y.-F. et al. Cisd2 deficiency drives premature aging and causes mitochondria-mediated defects in mice. Genes Dev 23, 1183–1194 (2009).
Wang, C. H., Kao, C. H., Chen, Y. F., Wei, Y. H. & Tsai, T. F. Cisd2 mediates lifespan: is there an interconnection among Ca2 + homeostasis, autophagy, and lifespan? Free Radical Res 48, 1109–1114 (2014).
Tekın, S., Türker, H., Güven, T. & Yel, M. The effects of ultraviolet C radiation on the ultrastructure of the liver cells of mole rats. Ultrastruct Pathol 40, 51–56 (2015).
Sugitani, N., Sivley, R. M., Perry, K. E., Capra, J. A. & Chazin, W. J. XPA: A key scaffold for human nucleotide excision repair. DNA Repair 44, 123–145 (2016).
Shigechi, T. et al. ATR-ATRIP Kinase Complex Triggers Activation of the Fanconi Anemia DNA Repair Pathway. Cancer Res 72, 1149–1156 (2012).
Kim, T. M. et al. RECQL5 and BLM exhibit divergent functions in cells defective for the Fanconi anemia pathway. Nucleic Acids Res 43, 893–903 (2015).
Scanlon, S. E. & Glazer, P. M. Hypoxic stress facilitates acute activation and chronic downregulation of fanconi anemia proteins. Mol Cancer Res 12, 1016–1028 (2014).
Park, W. H. Overexpression of the Fanconi Anemia A Gene in Hela and MCF10A Cells. Korean J Hematol 41, 1–7 (2006).
Wang, J. et al. Overexpression of the Fanconi anemia group C gene (FAC) protects hematopoietic progenitors from death induced by Fas-mediated apoptosis. Cancer Res 58, 3538–3541 (1998).
Aggarwal, M. et al. Werner syndrome helicase has a critical role in DNA damage responses in the absence of a functional fanconi anemia pathway. Cancer Res 73, 5497–5507 (2013).
Su, F. et al. Replication stress induced site-specific phosphorylation targets WRN to the ubiquitin-proteasome pathway. Oncotarget 7, 46 (2016).
Goto, M., Miller, R. W., Ishikawa, Y. & Sugano, H. Excess of rare cancers in Werner syndrome (adult progeria). Cancer Epidemiol Biomarkers Prev 5, 239–246 (1996).
Maity, J., Bohr, V. A., Laskar, A. & Karmakar, P. Transient overexpression of Werner protein rescues starvation induced autophagy in Werner syndrome cells. BBA - Mol Basis Dis 1842, 2387–2394 (2014).
Mori, C. et al. Over-expression of ATR causes autophagic cell death. Genes Cells 18, 278–287 (2013).
Thieme, R. et al. Analysis of alpha-2 macroglobulin from the long-lived and cancer-resistant naked mole-rat and human plasma. PLoS One 10, e0130470 (2015).
Lindner, I. et al. 2-Macroglobulin inhibits the malignant properties of astrocytoma cells by impeding β-catenin signaling. Cancer Res 70, 277–287 (2010).
Pawaria, S., Kropp, L. E. & Binder, R. J. Immunotherapy of tumors with α2-macroglobulin-antigen complexes pre-formed in vivo. PLoS One 7, e50365 (2012).
Shams, I., Avivi, A. & Nevo, E. Hypoxic stress tolerance of the blind subterranean mole rat: Expression of erythropoietin and hypoxia-inducible factor 1α. Proc Natl Acad Sci USA 101, 9698–9703 (2004).
Trapnell, C. et al. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nature Protocols 7, 562–578 (2012).
Trapnell, C., Pachter, L. & Salzberg, S. L. TopHat: discovering splice junctions with RNA-Seq. Bioinformatics 25, 1105–1111 (2009).
Langmead, B. & Salzberg, S. L. Fast gapped-read alignment with Bowtie 2. Nat Methods 9, 357–359 (2012).
Trapnell, C. et al. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol 28, 511–515 (2010).
Ashburner, M. et al. Gene Ontology: tool for the unification of biology. Nat Genet 25, 25 (2000).
Maere, S., Heymans, K. & Kuiper, M. BiNGO: a Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 21, 3448–3449 (2005).
Shannon, P. et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res 13, 2498–2504 (2003).
Kanehisa, M. & Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res 28, 27–30 (2000).
Kanehisa, M., Goto, S., Sato, Y., Furumichi, M. & Tanabe, M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res 40, D109–D114 (2012).
Wang, J., Duncan, D., Shi, Z. & Zhang, B. WEB-based GEne SeT AnaLysis Toolkit (WebGestalt): update 2013. Nucleic Acids Res 41, W77–W83 (2013).
Zhang, B., Kirov, S. & Snoddy, J. WebGestalt: an integrated system for exploring gene sets in various biological contexts. Nucleic Acids Res 33, 741–748 (2005).
Krupp, M. et al. RNA-Seq Atlas - a reference database for gene expression profiling in normal tissue by next-generation sequencing. Bioinformatics 28, 1184–1185 (2012).
Tacutu, R. et al. Human ageing genomic resources: integrated databases and tools for the biology and genetics of ageing. Nucleic Acids Res, gks1155, (2012).
De Magalhães, J. P. et al. The Human Ageing Genomic Resources: online databases and tools for biogerontologists. Aging Cell 8, 65–72 (2009).
Edgar, R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 32, 1792–1797 (2004).
Tamura, K. et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol 28, 2731–2739 (2011).
Yang, Z. PAML: a program package for phylogenetic analysis by maximum likelihood. CABIOS 13, 555 (1997).
Yang, Z. PAML 4: phylogenetic analysis by maximum likelihood. Mol Biol Evol 24, 1586–1591 (2007).
Voloch, C. M., Vilela, J. F., Loss-Oliveira, L. & Schrago, C. G. Phylogeny and chronology of the major lineages of New World hystricognath rodents: insights on the biogeography of the Eocene/Oligocene arrival of mammals in South America. BMC Res Notes 6, 160 (2013).
Acknowledgements
H.S. and T.H. acknowledge financial support by the University of Mainz Center for Computational Sciences (SRFN) and the Stiftung Innovation Rhineland-Palatinate (1049). A.A., I.S. and A.M. acknowledge financial support from the John Templeton Foundation, USA (grant #53057) and the donation of Mr. Peter Kadas, UK. We thank Bettina Weich for performing RNA preparation, Dr. Steffen Rapp for operating the Illumina HiSeq machine and Dr. Dennis Strand (University Medical School Mainz) for critical reading of the manuscript.
Author information
Authors and Affiliations
Contributions
T.H., I.S. and A.A. conceived the study, designed experiments and coordinated research. A.A. and I.S. performed hypoxia experiments and tissue preparation. H.S., A.M. and T.H. performed sequencing and differential expression analysis. H.S., A.M., A.B., G.P., I.S. and T.H. analysed data. A.B. and G.P. performed q-RT-PCR experiments. H.S. and T.H. drafted the initial version of the manuscript. All authors participated in complementing the manuscript and read and approved the final version.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schmidt, H., Malik, A., Bicker, A. et al. Hypoxia tolerance, longevity and cancer-resistance in the mole rat Spalax – a liver transcriptomics approach. Sci Rep 7, 14348 (2017). https://doi.org/10.1038/s41598-017-13905-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-13905-z
This article is cited by
-
Hypoxic and hypercapnic burrow conditions lead to downregulation of free triiodothyronine and hematocrit in Ansell’s mole-rats (Fukomys anselli)
Journal of Comparative Physiology B (2024)
-
Distinct organization of adaptive immunity in the long-lived rodent Spalax galili
Nature Aging (2021)
-
Local adaptations of Mediterranean sheep and goats through an integrative approach
Scientific Reports (2021)
-
Nontraditional systems in aging research: an update
Cellular and Molecular Life Sciences (2021)
-
Comparative morphometric analysis of lungs of the semifossorial giant pouched rat (Cricetomys gambianus) and the subterranean Nigerian mole rat (Cryptomys foxi)
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
