
Abstract
Queensland fruit fly, Bactrocera tryoni (Froggatt), is the most significant pest of Australia’s $9 billion horticulture industry. The sterile insect technique (SIT) and cue-lure (a synthetic analogue of raspberry ketone (RK))-based male annihilation technique (MAT) are two of the most effective management tools against this pest. However, combining these two approaches is considered incompatible as MAT kills sterile and ‘wild’ males indiscriminately. In the present study we tested the effect of pre-release feeding of B. tryoni on RK on their post-release survival and response to MAT in field cages and in a commercial orchard. In both settings, survival was higher for RK supplemented adults compared to control (i.e. RK denied) adults. A lower number of RK supplemented sterile males were recaptured in MAT baited traps in both the field cages and orchard trials compared to RK denied sterile males. The advantage of this novel “male replacement” approach (relatively selective mortality of wild males at lure-baited traps while simultaneously releasing sterile males) is increasing the ratio of sterile to wild males in the field population, with potential for reducing the number of sterile males to be released.
Similar content being viewed by others


Introduction
Fruit flies of economic importance belong to the family Tephritidae, which is among the largest families of Diptera, comprising approximately 4000 species1. A total of 180 fruit fly species are established as economically significant pests across 118 countries2. The most important fruit fly pest species in Australia is the Queensland fruit fly, Bactrocera tryoni (Froggatt), which attacks almost all commercial fruits and many vegetables3. Bactrocera tryoni causes economic losses through direct crop damage and associated control costs and is a significant barrier to national and international horticultural market access4. Among management options for this pest, the sterile insect technique (SIT) and male annihilation technique (MAT) are two important Area Wide Integrated Pest Management tools currently in use in Australia at national, regional and smaller scales4.
In SIT programs, sterile males are released to mate with wild females; the mated females are then unable to produce viable offspring5,6. Consequently, pest populations are suppressed or may even be eradicated5. In SIT programs, tephritids, including B. tryoni, are typically released as immature adults, one to three days after eclosion7,8,9. Therefore, upon release, sterile males must first survive to maturation before successfully locating and copulating with wild females10.
SIT programs are more effective and less costly if the density of wild individuals is reduced prior to the release of sterile insects, because this reduces the numbers of sterile insects that need to be reared and released. For tephritids, MAT is a potential means of reducing populations of wild males11,12. MAT uses large numbers of ‘bait stations’, containing a male attractant and an insecticide, which can significantly reduce local male densities11,13,14,15. Bait stations may take multiple forms, with some recent methods employing a waxy matrix that contains an attractant and insecticide11,15,16,17,18.
For managing B. tryoni, MAT and SIT are typically used consecutively rather than concurrently, because MAT kills sterile as well as wild males. Accordingly, MAT is used as a first step to reduce local densities of wild males. Thereafter, MAT is typically terminated, and sterile males released. Modelling studies19of tephritids, other than B. tryoni, suggest that MAT and SIT are synergistic and are more effective in combination than when used alone or consecutively provided that traps preferentially capture wild rather than sterile males.
Studies of several Bactrocera species show that pre-release exposure of males to plant-derived semiochemicals and synthetic lures dramatically reduces their subsequent response to attractant lures used in MAT20,21. Several commercially available lures and other male attractants are known for B. tryoni, including cue-lure, melolure, raspberry ketone (RK), RK formate, RK trifluoroacetate, and zingerone, which are all structurally similar11,15,22,23,24,25. While RK and zingerone are naturally occurring, cue-lure, RK formate, and RK trifluoroacetate are synthetic analogues of RK26. However, RK and cue-lure are more attractive than zingerone to B. tryoni 27. Usually, the majority of B. tryoni are attracted to cue-lure when mature, i.e. 8–12 days age28. Recent laboratory studies have shown that feeding by fertile B. tryoni males on cue-lure and RK triggers physiological changes that lead to accelerated sexual maturation and enhanced mating success29,30, but the consequent effect on male response to MAT devices has not been studied. Further, studies to date have used fertile males and have not considered sterile males as used in SIT.
Here, we consider the hypothesis that pre-release feeding on RK by immature sterile adult male B. tryoni may change their subsequent response to cue-lure based MAT. This is important because a reduction in attraction of sterile males would open the practical possibility of using MAT concurrently with SIT. Accordingly, this study examined the response of sterile B. tryoni to cue-lure in field cages and in a commercial orchard after feeding on RK prior to release. Subsequent survival of sterile male B. tryoni was also assessed to examine any adverse effects of pre-release RK exposure.
Results
RK supplementation improves B. tryoni survival
Field cage assay
Mortality of sterile male B. tryoni was reduced when they were fed on a diet of 1% or 2% RK compared with the control (RK denied). The shape of the survival curves differed between the four RK treatments (F4,545 = 8.03; p < 0.001) (Fig. 1a). No significant difference was found in the daily mortality rate between control and 0.5% RK fed B. tryoni (F2,247 = 0.24; p > 0.05; Fig. 1a). The daily mortality rate for each RK diet treatment was 2.57% (0%), 2.32% (0.5%), 1.36% (1%) and 1.33% (2%). The mean survival of sterile male B. tryoni after 5 weeks was also higher for 1% and 2% RK diet treatments than 0.5% or the control (p < 0.05; Fig. 1b).

Survival of sterile male B. tryoni when fed a raspberry ketone (RK) supplemented diet when immature. (a) Survivorship curves show the daily survival (b) mean survival after 5 weeks. Means followed by the same letter are not significantly different (p > 0.05).
Commercial orchard assay
The survival of RK supplemented B. tryoni was higher compared to the control. The estimated daily survivorship of RK supplemented B. tryoni was 0.9407 (dependent variable: ln[1 + mean recaptures] independent variable: time since last release; intercept = 1.7943; slope = −0.06113). This is equivalent to survival of 11.7% at 5 weeks. For control B. tryoni, daily survivorship was estimated to be 0.9337 using the same linear regression model (intercept = 2.1903; slope = −0.0687), equating to 9.04% survival at 5 weeks. Full regression details are given in Table 1.
RK supplementation reduces the attraction of sterile male B. tryoni to cue-lure
Field cage assay
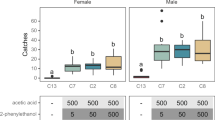
The number of RK supplemented and control sterile males recaptured in cue-lure baited traps differed significantly among treatments (F3,10 = 8.96; p = 0.003; Fig. 2). Reduced numbers of 0.5% and 1% RK supplemented sterile male B. tryoni were captured compared to the RK denied control males (F1,10 = 21.92; p < 0.001). Sterile male recaptures did not differ between 2% RK and control males (F1,10 = 2.71; p = 0.131; Fig. 2).

Effect of feeding raspberry ketone (RK) to immature sterile male B. tryoni on their recapture in cue-lure baited Lynfield traps in walk-in field cages. Means followed by the same letter are not significantly different (p > 0.05).
Commercial orchard assay
Pre-release RK supplementation of the immature fly diet changed the response of sterile male B. tryoni to cue-lure in field trials. A significantly lower number of RK fed individuals were found in cue-lure baited traps in the orchard compared to RK denied control flies (p = 0.01; Table 2).
Discussion
Here we show that simultaneous use of SIT and MAT is made viable by pre-release feeding of sterile B. tryoni. RK treatment increased the survival of sterile B. tryoni males under semi-field experimental conditions, with validation under commercial field conditions. In a SIT program, millions of sterile flies are typically released in the field, so even a slight improvement in the survival rate will result in increased abundance of sterile B. tryoni and a higher ratio of sterile to wild males, leading to enhanced suppression of B. tryoni pest populations4.
While this is the first study to test immature sterile male B. tryoni under field conditions, two prior studies have investigated effects on fertile males in the laboratory. With continuous access to liquid cue-lure (>95% purity) over the first 8 weeks of adult life, B. tryoni males displayed greater mortality than control males denied access to cue-lure29. In contrast, a powdered form of RK (1.25–5%) mixed with the adult diet and provided to fertile flies aged 0–24 h, which were permitted to feed for 48 h, did not affect the longevity of B. tryoni 30. The inconsistency between those two studies is likely due to the markedly different lengths of feeding permitted, with only the 8-week long regime having an effect in the form of reduced survival. Reasons for the improved survival of B. tryoni in the present study are not clear but maybe associated with the RK supplementation technique.
There is now solid evidence that when sterile31 or fertile32,33,34 fruit fly males ingest a semiochemical lure, or its analogue, subsequent attraction to that same lure significantly decreases. The present study is, however, the first to show a reduced response to cue-lure of RK supplemented immature B. tryoni. Previous studies have focused on methyl eugenol-responding species, and to a lesser extent RK/cue-lure responding species21,35. Bactrocera cucurbitae was found to be 3–10 times less responsive to traps baited with cue-lure after 4-5 days exposure to cue-lure32. Other Bactrocera species, such as B. dorsalis, were less likely to be captured in methyl eugenol (ME) baited traps after being fed ME supplemented diets33,34,36. While two previous studies have demonstrated an effect of lure feeding with cue-lure and RK on B. tryoni mating performance29,30, the field cage study and the open field experiment results presented in this study support an additional advantage of reduced responsiveness to cue-lure by RK-supplemented immature B. tryoni.
In this study trap-capture of sterile B. tryoni was reduced with RK supplementation under field-cage and field conditions, although the response was variable. The higher RK treatment (2%) in the field cage assay resulted in higher captures relative to the 1% dose, and survival at the lowest tested dose (0.5%) was comparable to the control. However, the field study demonstrated increased survival and reduced trap capture of RK supplemented flies, when provided a low dose. Future studies are required to determine the optimal RK dose that provides both a reduced response to MAT and a positive (or neutral) influence on other physiological, ecological and behavioural traits of sterile males. As current B. tryoni SIT involves the release of sterile females in addition to males (i.e. bisex strain), the effects of RK on female longevity also requires investigation. Future field studies also need to use a survival monitoring method that does not employ a semiochemical lure to tease apart the effects on survival and attraction to lure-baited traps.
The concept of the concurrent use of SIT and MAT to improve management of tephritids dates back to the 1970s, but its use has been hampered by the attraction of sterile as well as fertile male flies to MAT traps. Reducing captures of sterile flies, as the present study shows to be possible, is likely to allow increased SIT efficiency and reduced cost. Area wide management programs incorporating concurrent use of SIT and MAT would benefit from increased survival and reduced vulnerability to MAT by RK fed immature B. tryoni (Fig. 3) and may reduce the number of sterile flies required for release.

The comparative advantages of pre-release raspberry ketone (RK) supplementation of the immature sterile adult Bactrocera tryoni diet in sterile insect technique (SIT) programs which incorporate the use of the male annihilation technique (MAT), resulting in increased sterile male survival and a reduced response to MAT, and therefore increased suppression of wild B. tryoni populations.
Conclusion
Area wide management SIT programs are costly, and reducing the cost is a continual focus of research efforts. This study shows the first evidence that a diet containing RK fed to immature sterile B. tryoni prior to release improves subsequent survival of sterile males and reduces their response to MAT. Semiochemical-mediated enhancement of sterile flies, utilising RK, has the potential to enable simultaneous use of SIT and MAT in operational SIT programs, reducing costs and increasing effectiveness of such programs.
Methods
Study Insects
Bactrocera tryoni (bisex strain) were obtained as pupae from the Fruit Fly Production Facility (FFPF) at the Elizabeth Macarthur Agricultural Institute, Menangle, New South Wales (NSW), Australia. Insects were reared on a standard lucerne chaff diet37,38 in a controlled environment room under standard conditions (i.e., 26 ± 2 °C, 65 ± 10% relative humidity [RH], and a photoperiod of 14:10 [L:D] h, with a simulated dawn and dusk as the lights ramp up and down at the beginning and end of the light phase). Late-stage B. tryoni pupae were irradiated under hypoxia at 60–65 Gy of gamma irradiation from a cobalt-60 source at the Australian Nuclear Science and Technology Organisation (ANSTO) facility at Lucas Heights, NSW, which is the standard protocol to produce insects for SIT in Australia.
Effect of RK supplementation on the survival of B. tryoni
Field cage assays
Each week, approximately 10 g of sterile pupae (approximately 1000 flies) were placed in each of four cages (30 × 30 × 30 cm, Bugdorm cage, Megaview Science Co Ltd, Taiwan) and maintained during the experimental period under standard laboratory conditions. Soon after adult eclosion (0–24 h), remaining unemerged pupae were discarded, and the newly emerged adults (age 0–24 h) were provided with 100 g of a single RK diet treatment (see below) on a Petri dish and water and permitted to feed for 48 h. Prior to sterile male release in field cages (see below), females were separated and discarded.
The agar based diet for adult B. tryoni was prepared with the following ingredients: agar, sugar and yeast hydrolysate enzymatic (MP Biomedical, LLC, OH, USA; 60% protein) (3:1 by weight), and water. To prepare the diet, sugar and agar were mixed with water and boiled for 5–10 min. The mixture was allowed to cool to 40 °C, then yeast hydrolysate enzymatic was added to this mixture and stirred gently until combined.
The RK powder (99%, Sigma Aldrich, USA) was first dissolved in absolute ethanol and left for 24 h prior to incorporation with the agar-based diet. Immediately after diet preparation, the RK solution was added to the diets separately at a concentration of 0.0% (control), 0.5%, 1.0%, and 2.0% RK and then gently stirred with an electric stirrer to achieve uniform mixing. The diet was then allowed to cool and solidify for 30 min at room temperature before use.
To evaluate the survival of sterile male flies in field cages, PVC pipe-framed field cages (1.5 × 1 × 1 m) with zippers to allow access into the cage were placed over potted citrus trees (Citrus aurantium, Seville Orange) that had leaves but no fruit or flowers. Fifty sterile male flies, aged 48–72 h from each RK diet treatment were released in a field cage, and a total of three cages were maintained for each treatment. Each field cage contained yeast hydrolysate and sugar as food and water provided ad libitum. The number of dead flies were recorded daily and removed from the field cages. The trial continued until over 75% fly mortality was reached in the control treatment at five weeks after fly release. Mean climatic conditions during the survival assay periods were 21.95 ± 0.10 °C (min), 28.0 ± 0.40 °C (max) and 19.1 ± 1.18% RH (min), 58.0 ± 5.75% RH (max).
Commercial orchard Study site
Warroo (27°38′00″S, 148°45′00″E) near Stanthorpe, Queensland is a 60 ha orchard (approximately 75,000 trees) of plum variety ‘Queen Garnet’ and pollinator trees. Twenty sterile fly release sites on a 400 m grid were established. Release sites were located central to four trapping sites, located at least 150 m from any trap.
Sterile adult release protocol
Irradiated pupae obtained from the FFPF as described above were transported by air and road to Warroo. Upon receipt, pupae were divided into 200 g lots and placed inside translucent lidded plastic adult rearing containers (PARCs) (Silverlock MH 0110, colour ‘natural’, size: 645 × 413 × 275 mm; or Ezy Storage 31619, colour ‘clear’, size: 570 × 354 × 380 mm), with a 430 × 200 mm, mesh on the lid and a 150 × 100 mm mesh on two sides of the container for ventilation. Additional resting space was provided by wedging cardboard dividers (approximately 160 mm in height) inside each container. To distinguish between treatments and release periods, the pupae were marked with 0.8 g dye (Fiesta FEX 1 fluorescent pigments, Swada, 30–32 Kilkenny Court, Dandenong South, Victoria, Australia) per 100 g pupae, with dye colour (arc chrome, strong magenta, pink and stellar green) alternated between treatment types for each period of release. Dye pigments used in this experiment have no significant effect on emergence, flight and trap recapture rates for B. tryoni 39.
Two release periods, each comprising weekly releases over five consecutive weeks, were conducted, and average pupal weights ranged between 9.0–9.9 mg for release period 1 and between 9.2–9.7 mg for release period 2, which was within the expected range produced by the FFPF38.
During each release, PARCs containing sterile flies were taken to the release sites, and one PARC containing RK-fed flies and one PARC containing RK-deprived flies was opened at each site and the flies allowed to disperse freely. The sterile B. tryoni were aged 3 days when released.
Diet treatments
For flies fed 0.2% RK, an agar block (6 × 4 × 2 cm) containing a mixture of white sugar, yeast hydrolysate (3:1 by weight) and water40, mixed with RK was placed inside the PARC (20 PARCs) on a plastic board on top of cardboard dividers before fly emergence. For ‘RK-deprived’ flies, an agar block with yeast hydrolysate and sugar was provided without the added RK (a further 20 PARCs). The pupae in the PARCs were acclimated under field conditions in a roofed shed until release. Mean climatic conditions were for release period 1: 14.4 ± 0.51 °C (min), 26.0 ± 0.42 °C (max) and 54.0 ± 3.14% RH (min), 74.4 ± 1.53% RH (max) and release period 2: 10.2 ± 0.58 °C (min), 22.8 ± 0.36 °C (max) and 49.1 ± 2.21% RH (min), 72.7 ± 1.8% RH (max). Temperature and RH data for Warroo, Queensland, were obtained from the Bureau of Meteorology, Australia.
Recapture of sterile male B. tryoni
Recaptures were made using a 400 m spaced trapping grid comprising 21 Lynfield traps baited with cue-lure (International Pheromones, London) and malathion41 positioned at 1.5–2 m height in trees. Traps were located throughout the orchard, in adjacent native vegetation, and two single residential properties, each located within several hundred metres from the orchard. Traps were cleared weekly after each release until six weeks after the final release. Vials containing trapped flies were then sent to EMAI, where they were assessed and classified by colour as RK-fed or RK-denied39. The total number of B. tryoni (for each RK diet treatment) caught in the trapping grid was used to estimate survival and response to cue-lure baited traps.
Effect of RK supplementation on the attraction of sterile male B. tryoni to cue-lure
Field cage assay
After 48 h, sterile male B. tryoni, from each RK diet treatment were transferred and maintained in individual BugDorm cages (120 flies/cage). Food and water were provided as described above until release in field cages.
To evaluate the attraction of mature male flies to MAT lures, a single potted citrus tree (as described above) was placed in each of four walk-in circular field cages (4.0 m diameter, 2.5 m height). In each cage, a single Lynfield trap, baited with a MAT comprising cue-lure and malathion (Bio Trap Australia Pty Ltd), was placed in the tree canopy at 1.5 m height. One hundred sterile male flies aged 10 days from a single diet treatment were released in a field cage. The number of flies recaptured in traps was recorded every 24 h for 4 consecutive days. The experiment was repeated four times using sterile B. tryoni from four different generations, with a total of 1600 flies tested for their responsiveness to cue-lure. Mean environmental conditions during the periods of MAT lure response evaluation were 16.81 ± 0.89 °C (min), 35.73 ± 1.28 °C (max) and 37.73 ± 3.08% RH (min), 85.99 ± 2.69% RH (max).
Commercial orchard
The protocol was as described above for survival.
Statistical analyses
Effect of RK supplementation on survival of sterile male B. tryoni in field cages
Prior to survival analysis of data from field cages, observed time of an individual insect death was randomly allocated to time between the last observation time and the recorded time. This individual mortality time is also called accelerated failure time (AFT). The AFT were analysed using a survival analysis. After examination of the observed versus fitted survival function on iterated log (time), Weibull distribution was the best fit to the AFT.
The survival (S) function considered to fit the data (t) was as follows
where \(\lambda =\exp (\sum {b}_{i}{X}_{i})\);
Xi is the ith treatment; b i is the shape parameter of the ith treatment, α is the scale parameter of Weibull distribution. The parameter λ measures the instantaneous mortality rate (i.e. daily mortality rate)42. All the parameters were estimated using a maximum likelihood estimation analysis, which was run via GenStat (Eighteenth Edition) for Windows using RSURVIVAL procedure. All significant differences are based on comparing the shape parameter (b i) estimates and their standard errors.
Effect of RK supplementation on the response of sterile male B. tryoni to cue-lure in field cages
To measure the responsiveness of B. tryoni to cue-lure in walk-in field cages, data on the number of males captured in cue-lure baited traps were analysed by fitting a generalized linear model with error assumed to follow a binomial distribution43. A logit link was used to relate the recapture data to the treatment effects and a restricted maximum likelihood (REML) analysis was used to estimate the effects. Least significant difference (LSD) at 5% level was used to determine significance between treatment effects.
Effect of RK supplementation on the survival and response of sterile male B. tryoni to cue-lure in a commercial orchard
The following model was used to relate recaptures of marked sterile males in traps baited with cue-lure to daily survival44,45:
where A is the number recaptured, N is the total number marked and released, a is the recapture probability, p is the daily survival probability and n is the number of days since release44. A least-squares linear regression of time since release against the natural logarithm of (1 + A) usually provides a good fit, and the antilogarithm of the slope can be interpreted to be p, the daily probability of survival. Survival at day x was calculated as p x 46. Overlapping releases occurred weekly for 5 weeks in Warroo. For analysis of the data, average per trap recaptures following the last release was considered. This was necessary due to overlapping sexual maturation and mortality of each released cohort, which could cause the number of responsive (“capturable”) flies to increase after release. As analysed, the recaptures are of a pool of flies released over those weeks, with average weekly per trap recaptures for the first 35 days since the last overlapping release as the response variable and time since last release in days as the independent variable. Only release period 1 was analysed as the model fit to the second period was not significant.
To analyse the responsiveness of the released flies to cue-lure in the field, a generalized linear model with errors assumed to follow a Poisson distribution with variance adjusted for heterogeneity was employed. A log-link function was used to relate insect counts to RK effects. Group means were compared using the least significant difference (LSD) test at 5% significant level. Observations within the first 3 weeks trapping dates were discarded from analysis due to very low trap catches.
References
White, I. M., Elson-Harris, M. M. Fruit Flies of Economic Significance: Their Identification and Bionomics. (CABI Publishing (1992).
Qin, Y., Paini, D. R., Wang, C., Fang, Y. & Li, Z. Global establishment risk of economically important fruit fly species (Tephritidae). PLoS ONE 10, e0116424, https://doi.org/10.1371/journal.pone.0116424 (2015).
Clarke, A. R., Powell, K. S., Weldon, C. W. & Taylor, P. W. The ecology of Bactrocera tryoni (Diptera: Tephritidae): what do we know to assist pest management? Ann. Appl. Biol. 158, 26–54, https://doi.org/10.1111/j.1744-7348.2010.00448.x (2011).
Jessup, A. J. et al. in Area-Wide Control of Insect Pests: From Research to Field Implementation (eds M. J. B. Vreysen, A. S. Robinson, & J. Hendrichs) 685–697 (Springer Netherlands, 2007).
Knipling, E. F. Possibilities of insect control or eradication through the sse of sexually sterile males. J. Econ. Entomol. 48, 459–462, https://doi.org/10.1093/jee/48.4.459 (1955).
Fanson, B. G., Sundaralingam, S., Jiang, L., Dominiak, B. C. & D’Arcy, G. A review of 16 years of quality control parameters at a mass-rearing facility producing Queensland fruit fly. Bactrocera tryoni. Entomol. Exp. Appl. 151, 152–159, https://doi.org/10.1111/eea.12180 (2014).
Dominiak, B. C., Westcott, A. E. & Barchia, I. M. Release of sterile Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae), at Sydney, Australia. Aust. J. Ex. Agric. 43, 519–528 (2003).
Meats, A., Smallridge, C. J. & Dominiak, B. C. Dispersion theory and the sterile insect technique: application to two species of fruit fly. Entomol. Exp. Appl. 119, 247–254, https://doi.org/10.1111/j.1570-7458.2006.00415.x (2006).
Reynolds, O. L., Orchard, B. A., Collins, S. R. & Taylor, P. W. Yeast hydrolysate supplementation increases field abundance and persistence of sexually mature sterile Queensland fruit fly, Bactrocera tryoni (Froggatt). Bull. Entomol. Res. 104, 251–261, https://doi.org/10.1017/S0007485313000758 (2014).
Gurr, G. M. & Kvedaras, O. L. Synergizing biological control: Scope for sterile insect technique, induced plant defences and cultural techniques to enhance natural enemy impact. Biol. Control 52, 198–207, https://doi.org/10.1016/j.biocontrol.2009.02.013 (2010).
Dominiak, B. C., Ekman, J. & Broughton, S. Mass trapping and other management option for mediterranean fruit fly and Queensland fruit fly in Australia. Gen. App. Ento. 44, 1–8 (2016).
Lloyd, A. C. et al. Area-wide management of fruit flies (Diptera: Tephritidae) in the Central Burnett district of Queensland, Australia. Crop Protect. 29, 462–469, https://doi.org/10.1016/j.cropro.2009.11.003 (2010).
Steiner, L. F. Methyl eugenol as an attractant for oriental fruit fly. J. Econ. Entomol. 45, 241–248, https://doi.org/10.1093/jee/45.2.241 (1952).
Vargas, R. I., Leblanc, L., Piñero, J. C. & Hoffman, K. M. in Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies (eds T. E. Shelly et al.) 493–511 (Springer Netherlands (2014).
Dominiak, B. C., Clifford, C. S. & Nielsen, S. G. Queensland fruit fly (Bactrocera tryoni Froggatt) attraction to and chemical analysis of male annihilation blocks using three concentrations of cuelure at Dubbo, New South Wales, Australia. PPQ 24, 157–160 (2009).
Vargas, R. I. et al. Attraction and mortality of oriental fruit flies to SPLAT-MAT-methyl eugenol with spinosad. Entomol. Exp. Appl. 131, 286–293, https://doi.org/10.1111/j.1570-7458.2009.00853.x (2009).
Reynolds, O. L., Osborne, T., Crisp, P. & Barchia, I. M. Specialized pheromone and lure application technology as an alternative male annihilation technique to manage Bactrocera tryoni (Diptera: Tephritidae). J. Econ. Entomol. 109, 1254–1260, https://doi.org/10.1093/jee/tow023 (2016).
Dominiak, B. C. & Nicol, H. I. Chemical analysis of male annihilation blocks used in the control of Queensland fruit fly Bactrocera tryoni (Froggatt) in New South Wales. PPQ 27, 31–35 (2012).
Barclay, H. J., McInnis, D. & Hendrichs, J. Modeling the area-wide integration of male annihilation and the simultaneous release of methyl eugenol-exposed Bactrocera spp. sterile males. Ann. Entomol. Soc. Am. 107, 97–112, https://doi.org/10.1603/AN13010 (2014).
Vargas, R. I., Shelly, T. E., Leblanc, L. & Piñero, J. C. Recent advances in methyl eugenol and cue lure technologies for fruit fly detection, monitoring and control in Hawaii. In Vitamins & Hormones Vol. 83 (ed Gerald Litwack) Ch. 23, 575–595 (Academic Press, 2010).
Benelli, G. et al. Sexual communication and related behaviours in Tephritidae: current knowledge and potential applications for integrated pest management. J Pest Sci 87, 385–405, https://doi.org/10.1007/s10340-014-0577-3 (2014).
Siderhurst, M. S. et al. Raspberry ketone trifluoroacetate, a new attractant for the Queensland fruit fly, Bactrocera tryoni (Froggatt). J. Chem. Ecol. 42, 156–162, https://doi.org/10.1007/s10886-016-0673-3 (2016).
Royer, J. E. Responses of fruit flies (Tephritidae: Dacinae) to novel male attractants in north Queensland, Australia, and improved lures for some pest species. Austral Entomology 54, 411–426, https://doi.org/10.1111/aen.12141 (2015).
Gillespie, P. Observations on fruit flies (Diptera: Tephritidae) in New South Wales. Gen. App. Ento. 32, 41–47 (2003).
Dominiak, B. C., Campbell, A. J., Jang, E. B., Ramsey, A. & Fanson, B. G. Field evaluation of melolure, a formate analogue of Cuelure, and reassessment of fruit fly species trapped in Sydney, New South Wales, Australia. J. Econ. Entomol. 108, 1176–1181, https://doi.org/10.1093/jee/tov048 (2015).
Tan, K. H., Nishida, R., Jang, E. B. & Shelly, T. E. in Trapping and the Detection, Control, and Regulation of Tephritid Fruit Flies: Lures, Area-Wide Programs, and Trade Implications (eds Todd Shelly et al.) 15–74 (Springer Netherlands (2014).
Fay, H. A. C. A highly effective and selective male lure for Bactrocera jarvisi (Tryon) (Diptera: Tephritidae). Aust. J. Entomol. 51, 189–197, https://doi.org/10.1111/j.1440-6055.2011.00847.x (2012).
Weldon, C. W., Perez-Staples, D. & Taylor, P. W. Feeding on yeast hydrolysate enhances attraction to cue-lure in Queensland fruit flies. Bactrocera tryoni. Entomol. Exp. Appl. 129, 200–209, https://doi.org/10.1111/j.1570-7458.2008.00768.x (2008).
Kumaran, N., Balagawi, S., Schutze, M. K. & Clarke, A. R. Evolution of lure response in tephritid fruit flies: phytochemicals as drivers of sexual selection. Anim Behav 85, 781–789, https://doi.org/10.1016/j.anbehav.2013.01.024 (2013).
Akter, H., Mendez, V., Morelli, R., Pérez, J. & Taylor, P. W. Raspberry ketone supplement promotes early sexual maturation in male Queensland fruit fly, Bactrocera tryoni (Diptera: Tephritidae). Pest Manag. Sci., https://doi.org/10.1002/ps.4538 (2017).
Shelly, T. E. & Villalobos, E. M. Cue lure and the mating behavior of male melon flies (Diptera: Tephritidae). Fla. Entomol. 78, 473–482 (1995).
Chambers, D. L., Ohinata, K., Fujimoto, M. & Kashiwai, S. Treating Tephritids with attractants to enhance their effectiveness in sterile-release programs. J. Econ. Entomol. 65, 279–282, https://doi.org/10.1093/jee/65.1.279 (1972).
Shelly, T. E. Consumption of methyl eugenol by male Bactrocera dorsalis (Diptera: Tephritidae): low incidence of repeat feeding. Fla. Entomol. 77, 201–208 (1994).
Shelly, T. E. Trapping male oriental fruit flies (Diptera: Tephritidae): does feeding on a natural source of methyl eugenol reduce capture probability? Fla. Entomol. 83, 109–111 (2000).
Shelly, T. Effects of methyl eugenol and raspberry ketone/cue lure on the sexual behavior of Bactrocera species (Diptera: Tephritidae). Appl. Entomol. Zool. 45, 349–361, https://doi.org/10.1303/aez.2010.349 (2010).
Tan, K. H., Nishida, R. & Toong, Y. C. Floral synomone of a wild orchid, Bulbophyllum cheiri, lures Bactrocera fruit flies for pollination. J. Chem. Ecol. 28, 1161–1172 (2002).
Fruit Fly Production Facility. Standard Operating Procedure for Sterile Insect Production (Larval Production). Vol. WI 4.8 3–4 (New South Wales Department of Primary Industries (2014).
Dominiak, B. C., Sundaralingam, S., Jiang, L., Jessup, A. J. & Barchia, I. M. Production levels and life history traits of mass reared Queensland Fruit Fly Bactrocera tryoni (Froggatt) (Diptera: Tephritidae) during 1999/2002 in Australia. PPQ 23, 131–135 (2008).
Reynolds, O. L., Smallridge, C. J., Cockington, V. G. & Penrose, L. D. The effect of release method and trial site on recapture rates of adult sterile Queensland fruit fly, Bactrocera tryoni (Froggatt) (Diptera: Tephritidae). Aust. J. Entomol. 51, 116–126 (2012).
Reynolds, O. L. & van der Rijt, V. Queensland Fruit Fly –Adult Sterile Insect Technique. (NSW Department of Primary Industries, Australia (2011).
Meats, A., Clift, A. D. & Perepelicia, N. Performance of permanent and supplementary traps for Mediterranean and Queensland fruit flies in South Australia 1975–2001: comparison of male lure and food lure traps. Gen. App. Ento. 31, 53–58 (2002).
Aitkin, M., Anderson, A., Francis, B. & Hinde, J. Statistical Modelling in GLIM. (Oxford University Press (1989).
McCullagh, P. & Nelder, J. A. Generalized Linear Models. Second edition edn, (Chapman and Hall 1989).
Gillies, M. T. Studies on the dispersion and survival of Anopheles gambiae Giles in East Africa, by means of marking and release experiments. Bull. Entomol. Res. 52, 99–127, https://doi.org/10.1017/S0007485300055309 (2009).
Manoukis, N. C., Jang, E. B. & Dowell, R. V. Survivorship of male and female Bactrocera dorsalis in the field and the effect of male annihilation technique. Entomol. Exp. Appl. 162, 243–250, https://doi.org/10.1111/eea.12536 (2017).
MacDonald, G. The epidemiology and control of malaria. (Oxford University Press (1957).
Acknowledgements
We would like to thank the Department of Education and Training, Australian Government, for providing an Endeavour Research Fellowship (5203/2016) to Prof. Mohammed Abul Monjur Khan (PhD). This project has been funded by the Graham Centre for Agricultural Innovation (an alliance between NSW Department of Primary Industries and Charles Sturt University). We are grateful to Bim Goodrich and Rowan Berecry for allowing the use of Warroo orchard for the field trial and for assisting with sterile releases. We thank Phil Taylor and Soo Jean Park for advice on RK. Anne Johnson, Mohammad Shadmany and William Cuddy are thanked for assistance drafting the manuscript. Opinions, findings, conclusions, or recommendations expressed in this publication are those of the authors and do not necessarily reflect the views of the USDA. USDA is an equal opportunity provider and employer.
Author information
Authors and Affiliations
Contributions
M.A.M.K., O.L.R. & G.M.G.: conceived and designed the experiments. M.A.M.K., O.L.R. & T.O.: performed the experiments. I.B. & N.C.M.: analyzed the data. M.A.M.K., N.C.M., G.M.G. & O.L.R.: wrote the manuscript; other authors provided editorial input.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Khan, M.A.M., Manoukis, N.C., Osborne, T. et al. Semiochemical mediated enhancement of males to complement sterile insect technique in management of the tephritid pest Bactrocera tryoni (Froggatt). Sci Rep 7, 13366 (2017). https://doi.org/10.1038/s41598-017-13843-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-13843-w
This article is cited by
-
A review on natural phenylbutanoid attractants: Occurrence, distribution, and role in nature, especially in relation to Dacini fruit fly behavior and pollination
Journal of Chemical Ecology (2024)
-
Raspberry ketone diet supplement reduces attraction of sterile male Queensland fruit fly to cuelure by altering expression of chemoreceptor genes
Scientific Reports (2021)
-
Different methods of methyl eugenol application enhance the mating success of male Oriental fruit fly (Dipera: Tephritidae)
Scientific Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
