
Abstract
Synapses are an early pathological target in many neurodegenerative diseases ranging from well-known adult onset conditions such as Alzheimer and Parkinson disease to neurodegenerative conditions of childhood such as spinal muscular atrophy (SMA) and neuronal ceroid lipofuscinosis (NCLs). However, the reasons why synapses are particularly vulnerable to such a broad range of neurodegeneration inducing stimuli remains unknown. To identify molecular modulators of synaptic stability and degeneration, we have used the Cln3 −/− mouse model of a juvenile form of NCL. We profiled and compared the molecular composition of anatomically-distinct, differentially-affected pre-synaptic populations from the Cln3 −/− mouse brain using proteomics followed by bioinformatic analyses. Identified protein candidates were then tested using a Drosophila CLN3 model to study their ability to modify the CLN3-neurodegenerative phenotype in vivo. We identified differential perturbations in a range of molecular cascades correlating with synaptic vulnerability, including valine catabolism and rho signalling pathways. Genetic and pharmacological targeting of key ‘hub’ proteins in such pathways was sufficient to modulate phenotypic presentation in a Drosophila CLN3 model. We propose that such a workflow provides a target rich method for the identification of novel disease regulators which could be applicable to the study of other conditions where appropriate models exist.
Similar content being viewed by others


Introduction
Synapses are an early pathological target in a range of diseases1,2 including conditions associated with advancing age (e.g. Alzheimer (AD)3,4 and Parkinson disease5,6), neurodevelopmental conditions (e.g. spinal muscular atrophy (SMA)7,8), protein misfolding/accumulation diseases (e.g. Huntington disease (HD)9), prion diseases10, spinocerebellar ataxias (SCA)11 and lysosomal storage disorders (Neuronal ceroid lipofuscinosis (NCLs or Batten disease)12,13,14,15,16. However, our understanding of the reasons why specific synaptic populations are so vulnerable to such a broad range of neurodegenerative stimuli, and the mechanisms that govern their stability, remains in its infancy2,17,18.
In this study, we sought to define the molecular regulators of synaptic stability, using animal models of CLN3 disease (a.k.a. juvenile NCL or JNCL, OMIM # 204200). The NCLs, are the most frequent autosomal-recessive neurodegenerative disease and form of dementia in childhood19. Incidence in the USA is estimated at 1.6–2.4/100,000 whereas in Scandinavian countries it is 2–7/100,00020,21. The term “NCL” currently encompasses up to 14 disease subtypes, which are grouped together due the lysosomal accumulation of autofluorescent storage material, distinct ultrastructural properties, broadly similar pathology and clinical features and a severe neurodegenerative phenotype16,22. Crucially for the purposes of this study, pre-synaptic disruption is a key early event in NCL, accurately predicting the distribution of subsequent neuronal loss12,13,14,15,16.
The knowledge of the underlying genetic cause and/or storage material composition have provided a base for the basic understandying of the pathogenesis and their correlation to the clinical progression of the disease, the design of gene replacement therapies and the development of animal models23,24,25. However, the pathways and molecular cascades leading to neurodegeneration in NCL, as in many other neruodegenerative diseases, are still unknown.
Mutations in CLN3 underlie a juvenile form of NCL, the most prevalent form worldwide26. The CLN3 gene encodes a putative transmembrane protein whose function is not completely understood. However, experiments in yeast and in vitro studies have revealed possible functions relating to vacuolar pH regulation27 and endocytic membrane trafficking28. Although CLN3 is ubiquitously expressed throughout the body, the most obviously affected tissues are neurologic based. This feature is shared by other monogenetic neurodegenerative conditions such as SMA29. Yet, the reasons why neurons appear to be particularly vulnerable to defects in such broadly expressed proteins is not understood.
In humans, the onset of CLN3 disease occurs typically between 4–7 years of age, when loss of visual acuity is identified. Learning deficits are followed by speech and motor difficulties and seizures. As no treatment is currently available, premature death results at a mean age of 24 years old30. Unlike other more complex neurodegenerative diseases, the growing knowledge of the NCL-causative genes has facilitated the development of powerful animal models in recent years. These models have significatively improved our understanding of the progressive nature of the different forms of NCL. Cln3 null mice (Cln3 −/−) reproduce various aspects of the human disorder31,32. Although neuronal loss is widespread in terminal disease31,33, differential vulnerability can be detected across brain regions and their respective resident cell types/subcellular compartments at specific time-points. This differential degenerative progression follows the same pattern in the vast majority of NCL murine models: during pre-symptomatic stages there is an early selectivity for relay neurons within the thalamic nuclei followed by the corresponding cortical areas14,31,32,33,34,35, as well as GABAergic hippocampal interneurons and Purkinje neurons31,32. These reports in murine models regarding the vulnerability of the thalamus correlates with MRI studies in human patients showing alterations in thalamic areas at “pre-clinical” stages30,36,37. Furthermore, differential regional/neuronal vulnerability is also a shared event in a great number of neurodegenerative diseases in which some brain regions seem affected earlier than others, such as in Alzheimer38,39,40, Parkinson41, Huntington diseases42,43.
In this study, we aimed to define the molecular regulators of both synaptic stability and vulnerability using animal models of NCL. We initially characterized differential patterns of pre-synaptic pathology in Cln3 −/− mice31 across two different time-points. This enabled the subsequent application of high-throughput proteomics in order to map the molecular fingerprint of differentially vulnerable biochemically-isolated pre-synaptic populations. We identify multiple cascades correlating with synaptic vulnerability and describe valine degradation and rho signalling pathways as two major regulators of synaptic vulnerability. The direct contribution of these pathways to neurodegeneration was confirmed in vivo using a Drosophila CLN3 model. Finally, we are confident that regulators of synaptic vulnerability and degeneration identified in the context of NCL are also conserved in their expression across other neurodegenerative conditions, in which pre-synaptic alterations are an early event2. This research therefore opens a window for further investigation into common molecular therapeutic targets and strategies for novel interventions across a range of neurodegenerative conditions during early disease onset.
Results
Quantitative immunohistochemistry identifies differential rates of pre-synaptic pathology between brain regions in Cln3 −/− mice
Although the spatio-temporal pattern of neuron loss in Cln3 −/− mice has already been studied32,33, little is known about the progression of synaptic pathology across differentially vulnerable brain regions. Studies in other NCLs using Ppt1 −/−, Cln6 −/− or CathD −/− mice have revealed progressive synaptic loss starting in the thalamus and followed later in corresponding cortical areas12,14. Hippocampal structures seem to be also affected early, although to a lesser extent than seen in the thalamus33. To investigate whether a similar pattern of synaptic pathology might be present in Cln3 −/− mice we studied the expression of the presynaptic marker synaptophysin (Syp; as previously described in12) in three brain regions that exhibit different degrees of neuronal vulnerability using quantitative immunohistochemistry (at 6.5 and 13 months; see methods – Fig. 1 and Supplementary Fig. S1). The three brain regions studied were: 1. Thalamus – ventral posteromedial/ventral posterolateral nuclei (VPM/VPL) and lateral geniculate nucleus (LGNd); 2. The corresponding cortical target regions in the primary somatosensory barrel field (S1BF) and primary visual areas (V1) respectively, and 3; Hippocampus- stratum radiatum and stratum oriens (Fig. 1A). At 6.5 months no significant changes in Syp expression were detected between Cln3 −/− and control mice in any of the brain regions studied (Supplementary Fig. S1). However, by 13 months, some differences between genotypes were apparent (Fig. 1B and C). Similar to mouse models of other NCLs, thalamic nuclei demonstrated greater synaptic pathology at 13 months as indicated by the lower Syp immunoreactivity to synaptophysin in Cln3 −/− vs. controls. Hippocampal structures were also affected, although as predicted, the difference in Syp immunoreactivity between genotypes in both hippocampal subfields was less than in thalamic nuclei. In contrast, cortical regions such as S1BF and V1 (corresponding to the thalamic nuclei VPM/VPL and LGNd) showed no statistical difference between genotypes in Syp immunoreactivity at 13 months. Immunohistochemistry using antibodies against VAMP2 and SNAP-25 (two more pre-synaptic markers) also hint at pre-synaptic alterations in the thalamus (Fig. S2–S4).

Spatio-temporal synaptic loss study in Cln3 −/− detected differentially vulnerable synaptic populations across brain regions. (A) Brain region schematic showing the brain areas measured in grey. Thalamic regions includes the ventral posterior medial/ventral posterior lateral thalamic nucleus (VPM/VPL) (top left) and the dorsal lateral geniculate nucleus (LGNd) (bottom left); their respective cortical projections in the primary somatosensory cortex (S1BF) (top right) and primary visual cortex (V1) respectively (bottom right); hippocampal regions measured within the CA1-3 were the stratum radiatum and stratum oriens (bottom). (B and C) Representative photomicrographs of coronal sections of the same brain regions immunostained with synaptophysin (Syp) and bar chart showing its corresponding quantification based on the area of immunoreactivity in 13 month old control and Cln3 −/− mice. Syp immunoreactivity was lower in thalamic regions (VPM/VPL and LGNd) in the Cln3 −/− mice when compared to controls indicating more pathology, detectable earlier in the thalamus. Hippocampal stratum oriens and stratum radiatum also showed reduced Syp immunostaining, although the difference between genotypes was smaller. Cortical regions did not show difference in immunoreactivity for synaptophysin indicating that no synaptic loss is happening in these cortical areas at 13 months. (Mean ± SEM; *P < 0.05; **P < 0.01; ***P < 0.001; ns P > 0.05, Student T test, 5 mice per each genotype and time-point were used, Scale bar = 200 um (left) and 20 um (right).
Thus, the pre-synaptic alterations seen here (as determined with the markers detailed above) appears to follow a similar pattern to that previously reported for neuronal loss in mouse models of other forms of NCL12,14, with the thalamus being the most affected region, followed by the hippocampus, whereas cortical measurements were still unaffected at this stage of disease. We would categorize these alterations in pre-synaptic markers as being moderate at 13 months, as the pathology as quantified here is less than 10% (9.57% relative to controls).
Comparative molecular profiling reveals proteomic perturbations that correlate with the extent of pre-synaptic alterations
Despite advances in the characterization of the mammalian synaptic proteome, little is known about how these proteins interact and the molecular mechanisms that govern synaptic vulnerability. After confirming the synaptic vulnerability pattern across three distinct brain regions, we wanted to examine if the onset of pre-synaptic pathology correlated with distinct molecular alterations in the proteome. To address this, we examined biochemically isolated pre-synaptically enriched fractions (through the generation of “crude” synaptosomes – see methods) produced from microdissected brain regions from Cln3 −/− mice and controls at 13 months of age where we characterise the pre-synaptic pathology as thalamus > hippocampus > cortex.
After synaptosome production and protein extraction, iTRAQ proteomics was carried out as detailed in Fig. 2. 1536 total proteins were identified in hippocampus and thalamic samples whereas 2068 were detected in cortex. We then pre-filtered the proteins by those which were identified by at least 2 peptides and therefore are more likely to be reliable identifications. 914 proteins passed that filter in hippocampus and thalamus and 1295 in cortex. Almost 40% of our pre-filtered thalamic proteome was altered when compared to control littermates. In contrast, only 20% and 17.6% of the synaptic proteome was altered in the cortex and hippocampus relative to controls respectively. The number of protein alterations also correlates with pre-synaptic pathology being more apparent in thalamic areas at this “moderate” stage of disease progression.

Experimental design workflow. Experiments are divided into in vivo (animal models), MS processing and in-silico where the different bioinformatics tools are detailed. *This schematic was reproduced for the three brain regions of interest: cortex, hippocampus, and thalamus at 13 months of age.

Differentially vulnerable synaptic population molecular profiling. (a) Venn diagram and heat map show the 709 common proteins identified and overlapped in cortical, hippocampus and thalamic synaptic proteomic datasets. (B) Heat map representing the significance (−log P value) of the canonical pathways identified in IPA across cortex, hippocampus and thalamus datasets. It is observed a preogressive increase in significance correlating to the synaptic vulnerability pattern described previously. (C) Heat map showing the 709 common proteins identified and overlapping in cortical, hippocampus and thalamic synaptic proteomic datasets. (D) BioLayout clustering 3D representation of proteomic expression data across differentially vulnerable synaptic populations orientated at 3 different angles. Each sphere represents a single protein and the edge represents how similar their expression trend is towards the other proteins in the dataset. The closer the spheres are the more similar expression trend they have. The colours represent the different clusters of co-expressed proteins. (E) Expression profile means in log scale (Cln3 −/− /WT) of co-expressed proteins in clusters 3, 4, 6 and 7 (Supplementary Tables S1) and its main biological function/subcellular compartment identified by DAVID enrichment analysis (See Supplementary Table S2). Clusters highlighted show steady up or downregulation across cortex (C), hippocampus (H) and thalamic (T) regions correlating with the vulnerability status of synapses.
Next, these pre-filtered proteins from the three pre-synaptic populations were aligned to look for overlapping proteins. 709 proteins were identified in all three brain regions and these were taken forward for comparative analysis (Fig. 3A).
In order to gain a broad and unbiased overview for the types of processes that may be represented by the protein alterations detected in all three pre-synaptic populations, we used Ingenuity pathway analysis (IPA) (see Methods – Fig. 3B). Interestingly, we identified multiple canonical pathways which IPA designated as perturbed in a strikingly progressive manner, consistent with the degree of pre-synaptic pathology outlined above i.e. −log P value = higher in thalamus > hippocampus > cortex (Fig. 3B). An example cascade can be found in more detail in Supplemetary Fig. S5. This analysis therefore indicates that in general, alterations in specific molecular cascades were increasing with pre-synaptic pathology, further confirming the differential vulnerability pattern previously identified by IHC. Thus, the synaptic vulnerability pattern described here appears consistent at both the morphological and that molecular level.
Whilst canonical cascade analysis is an interesting tool to use at the outset, multiple members of the same cascade do not necessarily need to change in the same manner, and whilst the trend of the majority of canonical cascades seen in Fig. 3B are consistent with the regional pathology reported, the individual protein alterations are far more complex (ass seen in the heatmap in Fig. 3C). Thus, to better understand and visualize the different molecular expression trends occurring across the three datasets, individual proteins were tracked using BioLayout Express 3D (see Methods). Simplistically, BioLayout is a complex pattern recognition software which generates a visual representation of the data based on protein abundance44. In this case the software interrogates a regional expression profile generated by arranging the individual proteomic sets in order of increasing pre-synaptic pathology/vulnerability (cortex < hippocampus < thalamus). The outcome of this is that proteins were found to cluster into 14 different groups according to similarities in their relative expression across these three regions (Fig. 3D)45. Of interest to us were Clusters 3 (70 proteins), 4 (68 proteins), 6 (51 proteins) and 7 (50 proteins) comprise proteins whose expression can be grouped (or clustered) together as having either a gradual upregulation or downregulation, therefore correlating (directly or inversely with) with pre-synaptic vulnerability (Fig. 3D and E, Supplementary Table S1). Next, we applied an enrichment analysis to these distinct clusters in order to determine if each expression trend could be associated with a specific biological or functional category (using the DAVID software tool – see methods and Fig. 3E). Unexpectedly, clusters with proteins that were increasingly upregulated in vulnerable regions were enriched for structural associated candidates like cytoskeletal-related proteins, and specific biological functions such as endocytosis (Cluster 4 and 7). Perhaps more surprisingly, clusters whose protein expression decreased with increasing vulnerability were related to specific mitochondrial functions (Clusters 3 and 6) (Fig. 3D and E and Supplementary Table S2). Thus, comparative proteomic profiling of differentially vulnerable pre-synaptic populations revealed that significant alterations to the synaptic proteome correlated with the degree of pathology seen at these relatively moderate disease stages.
In silico analysis highlights alterations in valine catabolic and ROCK2 signalling cascades in vulnerable pre-synaptic compartments
After confirming that the pre-synaptic proteome changes in a manner consistent with the differential vulnerability/regional disease progression in Cln3 −/− mice, we next sought to tease out the potential molecular regulators of neuronal stability from our complex proteomic datasets. To do this, we focused our analyses on the thalamus, because pre-synaptic compartments here were perturbed to a greater extent than those in the hippocampus or cortex (according to the pre-synaptic markers we employed). In the thalamic dataset, 1536 total proteins were identified following iTRAQ processing. Further DAVID enrichment analysis was applied to the unfiltered thalamic data to confirm that the starting material was suitably enriched for synaptic proteins (Table 1). As described above, we applied filtering criteria based on the number of peptides (more than 2 unique peptides), those changed > 20% (1.2 fold-change) and those which were mapped by Ingenuity Pathway Analysis (IPA) software. A total of 374 proteins met these criteria (Fig. 4A and Supplementary Tables S3–S4). Next, we confirmed the veracity of the filtered data by validating a range of proteins using quantitative fluorescent Western blotting (QWB) (Fig. 4B–G).

Synaptic thalamic proteome filtering and validation. (A) Dot plot demonstrating the process of proteomic data filtering. Each data point represents an individual protein identified using iTRAQ proteomic technique. LHS 1536 proteins were identified across all thalamic samples. Following filtering (see Methods) a molecular fingerprint for thalamic synaptic alterations comprising 374 candidate proteins was produced (RHS; see also Supplementary Tables S3 and S4). (B) QWB representative bands for two upregulated (β-catenin and ROCK2), two downregulated (HIBCH and α-synuclein) and one unchanged protein candidate (calretinin) verifying the proteomic data. (C and G). Quantification and statistical QWB analysis showing the magnitude of alteration in Cln3 −/− thalamic synaptic fractions. All the selected candidate proteins were altered as indicated by this iTRAQ analysis. Mean ± SEM; *P < 0.05; ***P < 0.001 (Student T test, n = 6 mice per each genotype).
Higher order functional clustering highlights similarities with other neurodegenerative conditions
Here, we used ingenuity pathway analysis (IPA) to elucidate which molecular pathways and biological networks were disrupted in “affected” thalamic pre-synaptic compartments. Here we input the total pre-filtered thalamic data into IPA and applied the 1.2 fold-change cut-off in the software. Of the three-hundred and seventy-four proteins recognised by IPA, 68.3% have previously been reported in the literature as being associated with neurological disease (Fig. 5A). The majority of the diseases and disorders which fall under the category of “neurological disease” are known to demonstrate synaptic pathology as an early event1. Interestingly, the main molecular and cellular functions identified in IPA were “molecular transport” and “cellular assembly and organization”. These functional categories are consistent with the comparative analysis of differentially vulnerable synaptic populations carried out above (see Fig. 3E), where clusters showing increases in protein expression from the levels detected in the comparatively spared cortical synapses through to greater levels again in the more vulnerable thalamic populations highlighted cytoskeletal protein alterations.

IPA analysis of the thalamic synaptic proteome. (A) Table of top 5 “diseases and disorders” and “molecular and cellular functions”. (B and C) Canonical pathways bar chart of representative pathways showing, (B) The percentage of upregulated and downregulated proteins within each pathway. Numbers on the top indicate the total number of proteins within the canonical pathway. (C) Significance of the association between the dataset and the canonical pathway (−log(p-value) and ratio) and z-score prediction of activation/inhibition (see Methods). Pathways highlighted in green and blue are associated to ROCK2 and HIBCH, respectively. (D) HIBCH and ROCK2 protein expression ratio across cortex (“less affected’) and thalamic (“degenerating”) synapses in these moderately affected Cln3 −/− mice (Cln3 −/−/WT). (E) HIBCH and ROCK2 protein expression is conserved in models of injury, following similar pattern of expression 24 hours post-injury when synapses start to degenerate2.
Molecular cascade tracking identified potentially conserved regulators of vulnerability
The “Canonical pathways” function in IPA revealed that, mitochondrial dysfunction was the most significant canonical pathway identified (see Methods). The contribution of synaptic mitochondria in neurodegeneration has been highlighted in recent years46, and mitochondrial abnormalities have also been related to CLN3 disease and other NCLs47,48,49,50, and our data is therefore consistent with these previous findings. However, in this study we wanted to consider pathways that have not previously been related to the NCLs before. Within the top canonical pathways identified with IPA, we also detect perturbations in ROCK signalling and valine degradation pathways (Fig. 5B and C). Examining these pathways, proteins such as ROCK2 and HIBCH were differentially expressed key “hub” components. Their expression was identified as tracking in BioLayout clusters 4 and 6 respectively correlating to the degree of pre-synaptic pathology in Cln3 −/− mice (see above, Figs 3D and 5). Moreover, we have previously identified ROCK2 and HIBCH expression alterations in pre-synaptic striatal isolates following cortico-striatal lesion injury (Fig. 5E), and demonstrated their potential to alter neurodegeneration in an injury specific context in vivo 2. Collectively these findings hinted that there may be a common molecular cascade underlying pre-synaptic pathology following a range of neurodegeneration inducing insults (from injury through to genetic). ROCK2 signaling and valine degradation pathways and specifically proteins such as ROCK2 and HIBCH, could therefore represent potentially novel regulators of synaptic stability in an NCL disease context, which have not been investigated yet. So we next sought to determine if such candidates could have the ability to moderate NCL disease progression in vivo.
Targeting the valine catabolic cascade is sufficient to modulate CLN3-induced neurodegeneration in vivo
HIBCH is a 3-hydroxyisobutyryl-CoA hydrolase protein that fulfils a core function within the valine degradation cascade51. Mutations in HIBCH cause a progressive infantile neurodegeneration in humans, characterized by hypotonia, motor delay and neurological regression51,52,53. Moreover, manipulation of HIBCH has been suggested to modulate injury-induced axonal degeneration2. However, despite this apparently restricted function, very little is known about its role (if any) in synaptic compartments or its binding/interaction partners (see Fig. 6A).

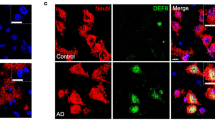
CLN3 induced degeneration is enhanced by the genetic downregulation of HIBCH and HADHA in vivo impacting in valine catabolism pathway. (A) HIBCH proteome interactome represented by IPA indicating only 6 gene/proteins are known to interact with HIBCH. (B) QWB bands and quantification of HIBCH in synaptic and “non synaptic” fractions of Cln3 −/− and control mice in thalamic protein extracts showing a downregulation in synapses and upregulation in non-synaptic parts **P < 0.01, ns P > 0.05 (Student T test). (C) QWB bands and quantification of cortical “crude” synaptosomes isolated from post mortem human JNCL patients (P) and controls (C) showing its downregulation at end stages in human disease *P<0.05 (Student T test). (D) HIBCH and HADHA are co-expressed and their expression trend correlates to the vulnerability status of synapses: unchanged in cortex (“spared”) and downregulated in thalamus (“degenerating”). (E) Valine degradation pathway represented with IPA. The first protein which expression is perturbed in the degradation process of valine is HADHA; upstream of HIBCH. (F–G) Representative light microscope images of Drosophila eyes of disease model (DM), DM + HIBCH RNAi and DM + HADHA RNAi and corresponding quantification of the eye surface area. Scale bar = 100um. ***P < 0.001 (One-way ANOVA and Tukey’s multiple comparison test as a post-hoc). (H) Schematic representation of the fourth and fifth steps of the valine catabolism cascade showing the enzymes HADHA and HIBCH catalyzing each step respectively. (A,E & H) Colored nodes illustrate proteins present in the thalamic dataset in Cln3 −/− with respect WT. Nodes in grey represent proteins changed <20%, down-/up-regulated proteins by >20% are represented in green and red respectively. Orange box highlights the steps that are disrupted in the valine degradation cascade. Dotted lines on representative westerns indicate cropping of bands from the same membrane.
Using QWB techniques we confirmed that HIBCH protein was less abundant in “crude” synaptosome extracts from thalamus in Cln3 −/− mice relative to wild type controls. Interestingly, HIBCH protein levels trended towards a slight up-regulation of HIBCH in “non-synaptic” compartments. Whilst not statistically significant, this non-synaptic increase may indicate a redistribution or altered trafficking of this protein in response to Cln3 deficiency (Fig. 6B). Importantly, examination of human post mortem brain samples showed that HIBCH protein levels were significantly reduced in JNCL patients (Fig. 6C).
From comparing synaptic vulnerability (Fig. 3), HIBCH belonged to Cluster 6 (gradual downregulation profile) and was co-expressed with other mitochondria-related proteins (Fig. 3E) including Hydroxyacyl-CoA Dehydrogenase/3-Ketoacyl-CoA Thiolase/Enoyl-CoA Hydratase (Trifunctional Protein), Alpha Subunit (HADHA), which also fulfils an enzymatic role within the valine degradation pathway (Fig. 6D). HADHA sits upstream of HIBCH in the valine catabolism cascade (Fig. 6E) and catalyzes the final three steps of mitochondrial long chain fatty acid β-oxidation54. HADHA codes for the α subunit of the mitochondrial trifunctional protein (αMTP), and its deficiency causes a metabolic disease that presents with Reye-like syndrome, with cardiomyopathy, neuromyopathy and sudden death in infancy55. More recent reports have indicated that HADHA deficiency results in a pigmentary retinopathy leading to vision loss56.
Given the important role of these two proteins in the valine catabolism cascade, we wanted to explore the consequences of experimental suppression of HIBCH and HADHA levels in vivo in order to assess their influence on synaptic and neuronal stability in an NCL-disease specific context. To do so, we used a CLN3 gain of function fly model previously characterized by Tuxworth et al. in which the Drosophila eye undergoes degeneration (becoming smaller and rougher)57. Although this is an accepted model for investigations into the cascades underpinning CLN3, the human disease is normally caused by loss of function. As with the human disease where dose is crucial to phenotype (i.e. heterozygous humans are not obviously affected) we can also demonstrate that degeneration ocuring in the Drosophila eye is CLN3 dose dependent (Fig. S6) as is the case for other neurodegeneration associated proteins (i.e. TDP43)58.
Thus, using this Drosphila model as a rapid screen for candidate ability to effect CLN3 driven phenotype (whilst bearing in mind its potential limitations) we first established a recombinant fly containing the GMR-GAL4 eye driver and the UAS-CLN3 gene in the second chromosome (GMR-GAL4;UAS-CLN3/CyOGFP, see Table 2), and crossed this with fly lines containing RNA interference (RNAi) of HIBCH and HADHA mouse orthologs. CG5044 and CG4389/MTPα orthologs were selected using the DRSC Integrative Ortholog Prediction Tool (DIOPT) (see Methods and Table 2). Progeny of the relevant genotype was counted (GMR-GAL4;UAS-CLN3/HibchRNAi and GMR-GAL4;UAS-CLN3/MTPαRNAi), eyes photographed and total surface area measured with Image J (see Methods). As expected, experimental suppression of either HIBCH or HADHA increased the neurodegenerative phenotypes observed (i.e. acting as enhancers of the CLN3-dependent phenotype) (Fig. 6F and G). Interestingly, the further upstream the valine degradation cascade is disrupted, the greater the effect on CLN3 phenotype (Fig. 6H). Interestingly, in this context neither HIBCH or HADHA were able to cause degeneration in the eye in the absence of the CLN3 disease background (Supplementary Fig. S7). This suggests that their manipulation alone is not sufficient to affect general neuronal stability, and that the effect seen when crossing HIBCH and HADHA with the CLN3 fly is likely mediated by the CLN3 allele. As such HIBCH and HADHA appear to be in vivo modifiers of neuronal stability in a CLN3-disease context, and this provides further evidence that valine catabolism may play a role in CLN3-dependent neurodegeneration.
Increased pre-synaptic ROCK2 expression appears conserved across a range of diseases and its downregulation can reduce CLN3-induced neurodegeneration in vivo
As discussed above, ROCK cascades were also identified as being perturbed in degenerating thalamic pre-synaptic compartments and were predicted (by in silico analysis) to be activated when compared to wildtype controls (Fig. 5B and C). ROCK2 is a Rho-kinase belonging to a family of serine/threonine kinases, of which isoform 2 is the most predominant in the brain59. ROCK2 directly interacts with, and is activated by, Rho GTPases. These are the central mediators of actin reorganization59, which are reported to have a key role in synaptic plasticity and long term potentiation60. Activation of ROCK2 has been implicated in several adult-onset neurodegenerative conditions where synaptic pathology is present, such as HD, AD, ataxia, and in Purkinje cell degeneration2,61,62,63,64. Moreover, ROCK2 has been shown to influence childhood neurodegenerative conditions such as SMA, where pharmacological inhibition partially recued symptoms and increases lifespan in an SMA mouse model65,66. Moreover, we have previously reported that ROCK2 is capable of influencing axonal degeneration in vivo following injury2. Taken together with our current data, this is consistent with a model whereby upregulation/activation of ROCK2 signalling in pre-synaptic compartments may correspond with increased neuronal vulnerability or degeneration in multiple infantile- and late-onset neurodegenerative conditions (Fig. 7D).

CLN3 induced degeneration is suppressed by the downregulation of ROCK2 genetically and pharmacologically (A) ROCK2 protein/gene interactome represented with IPA showing that some of the interactors of ROCK2 are also altered in “crude” thalamic synaptosomes. (B) QWB bands and quantification of control and Cln3 −/− synaptic and “non synaptic” fraction. Upregulation of ROCK2 is higher in synapses than in the non-synaptic fraction indicating a potential synaptic specific response. *P < 0.05 (T-student test). (C) “Signaling by Rho family GTPases” canonical pathway represented by IPA. Upregulation/activation of ROCK2 impacts in actin nucleation and polymerization that may disrupt actin dynamics in the synapse. (A&C) Coloured nodes illustrate proteins present in the thalamic dataset in Cln3 −/− with respect WT. Nodes in grey represent proteins changed <20%, down-/up-regulated proteins by >20% are represented in green and red respectively, orange indicates predicted activation and blue; predicted inhibition. (D) ROCK2 upregulation at early stage of disease is a converved event across animal models of injury, two adult-onset neurodegenerative diseases (Huntington and Spinocerebelar ataxia)2 and two childhood neurodegenerative conditions (SMA and the lysosomal storage disorders-NCLs). (E and H) Representative light microscope images of Drosophila eyes and corresponding 200x zoom of eye structure of (A) Disease Model (DM) (B) DM + 1.5 mM Fasudil. (C) DM + Rok RNAi TRiP and (D) Control fly (Canton S). (I) Bar chart representing the % of normalized suppression calculated from the average eye surface areas of three independent experiments as “(x- average eye surface area of the disease model)/(average of eye surface area of the control - average eye surface area of disease model) × 100” for each of the three independent experiments. Scale bar = 100um. ***P < 0.001, *P < 0.05 (One-way ANOVA and Tukey’s multiple comparison test as a post-hoc).
The ROCK2 interactome (displayed in Fig. 7A) illustrates its reported interactions with other gene/proteins, many of which were also altered in Cln3 −/− thalamic synaptic extracts. In the Cln3 −/− thalamus, QWB confirmed that ROCK2 was upregulated by 53.7% in the synaptic fractions, and by 27.7% in the corresponding “non synaptic” Cln3 −/− isolates, relative to controls (Fig. 7B), suggesting a more synaptically-focussed response to altered CLN3 expression. Our in silico analysis added support for this model by highlighting specific sub-cascades under multiple pathways related to ROCK2 such as “Signalling by Rho Family GTPases” and “Rho A signalling” (Fig. 5B and C). These cascades regulate actin dynamics and cytoskeletal organization (as shown in Fig. 7C) and can impact synaptic structure and affect synaptic transmission. Figure 7D summarises previously published data generated by our laboratory where ROCK2 was identified to be upregulated in a range of conditions at early stages of disease progression2 and new synaptic data regarding its role in CLN3 disease. Thus, the conserved increased expression of ROCK2 in Cln3 −/− mice, and in other related conditions and its ability to change degeneration, is highly suggestive of a potentially conserved contribution to synaptic vulnerability.
Thus, in order to investigate whether ROCK2 is capable of directly modulating the process of neurodegeneration in a CLN3-specific disease context in vivo, we again turned to the CLN3 Drosophila model. RNAi GD and RNAi TRiP fly lines for a ROCK2 ortholog were crossed to our established CLN3 recombinant Drosophila (see Table 2). Genetic downregulation of ROCK2 with two independently generated RNAi lines were tested and both were found to drastically improve their CLN3-induced small eye phenotype, although was more obvious with the TRiP library line (Fig. 7G–I). The total surface area of the eyes of the offspring (GMR-GAL4;UAS-CLN3/RokRNAi) were non-significant with respect the control eyes and therefore the phenotype was fully-recued. Qualitatively, there was also an obvious reduction/absence of black patches (indicative of cell death), and clear recovery of the gross structure of the Drosophila compound eye. With the RNAi GD line the eye size was also improved (25.22% of recue) (Fig. 7I). Thus, experimental manipulation of ROCK2 was sufficient to modulate neurodegeneration in a CLN3 model in vivo. To exclude the possibility of the suppression effect being due to the dilution of GAL4 among two UAS transgenes, a control experiment was carried out, where the CLN3 recombinant fly was crossed to a UAS-GFP line. No significant changes were detected in the eye of the offspring, indicating that the results showed in our RNAi experiments are likely a consequence of the downregulation of ROCK2.
The ROCK inhibitor Fasudil ameliorates CLN3-dependent neurodegeneration in vivo
Following the rescue of the CLN3-dependent eye degeneration by means of genetic tools we therefore wanted to test the possibility of targeting ROCK2 pharmacologically in Drosophila.
Several ROCK inhibitors have been characterized, but their specificity for individual ROCK kinases is still unclear. Fasudil is a small molecule that unspecifically inhibits ROCK2, originally discovered as a blocker of cerebral vasospasm in animal models67. Importantly, fasudil has been demonstrated to be safe and effective in clinical trials of cerebral vasospasm, pulmonary hypertension, Raynaud phenomenon and cancer68,69,70,71,72,73. Moreover, fasudil has previously been applied to murine models of SMA showing an improvement of the phenotype and lifespan of these mice65,66. However, fasudil also targets other kinases such as MAP4K4, PKC, PRKAA1 or PRKAA274, which are predicted to be also activated by IPA (Supplementary Fig. S8A). The activation of these kinases seem to have downstream consequences on biological functions relevant to NCL such as autophagy (Supplementary Fig. S8A) or and “Protein kinase A signalling” (Supplementary Fig. S8B and C). Given the success of the compound in mammalian systems65,66,73, including humans69,70, to date and the apparently conserved ROCK2 alterations across multiple-neurodegenerative paradigms, we decided to test Fasudil in CLN3 Drosophila (GMR-GAL4/UAS-CLN3). The drug was added into the fly food in order to obtain 1.5 mM of fasudil in the food and the Drosophila were raised as normal (see Methods). Pharmacological inhibition with fasudil caused a significant, albeit modest suppression of the CLN3-derived eye phenotype. Quantification of eye surface areas confirmed a 17% rescue of the CLN3 eye phenotype in the treated animals (Fig. 7E,F and I). Thus, targeting of ROCK cascades with fasudil was sufficient to ameliorate neurodegeneration in a CLN3 model in vivo.
Discussion
Here we show what may be the first proteomic profiling carried out across multiple differentially vulnerable synaptic populations at an early stage of disease progression. We have demonstrated that such unbiased proteomic mapping of distinct pre-synaptic populations coupled with in silico analysis, and in vivo rapid phenotypic screening in Drosophila, is an effective target-rich workflow for the identification of novel molecular alterations that regulate synaptic/neuronal stability.
Here we have confirmed that Cln3 −/− murine pre-synaptic pathology appears to follow the same neuronal differential vulnerability pattern as other NCL subtypes12,14. We show that, at 13 months of age in Cln3 −/− mice the synaptic pathology is more pronounced in the thalamus (although still <10%), followed by hippocampal pre-synaptic compartments, while cortical synaptic populations remain relatively unaffected (Fig. 1). iTRAQ proteomic and bioinformatics showed that differential synaptic pathology was also reflected at the molecular level, showing a progressive increase in the disruption of multiple canonical pathways correlating with our observations at the immunohistological level (Fig. 3B). Further analyses by means of complex pattern recognition software identified alterations in the pre-synaptic proteome (identified by iTRAQ analysis of “crude” synaptosomes) that correlated with the assigned vulnerability status of pre-synaptic populations in the Cln3 −/− mice. Pathway analysis highlighted perturbations in valine catabolism and rho signaling pathways, with proteins including HIBCH, HADHA and ROCK2 as key players, correlating with synaptic vulnerability. Furthermore, HIBCH and ROCK2 displayed a conserved direction of change in multiple neurodegenerative conditions (Figs 5E and 7D), and we have previously demonstrated that they can alter axonal degeneration in an injury dependent context2. Importantly, this suggests that not only do conserved pathways regulating degenerative processes exist, but they may be targetable across multiple conditions (Fig. 7D).
In contrast, no lysosomal specific pathways were identified in our in silico analysis in synapses, indicating that loss of CLN3 may be impacting in other pathways beyond lysosomal function. This is supported by studies in which CLN3 was found to be expressed in other locations such as in synaptosomes, lipid rafts, Golgi, mitochondria, glia and endothelial cells75,76,77,78,79, where it may play other roles such as regulation of oxidative stress78,80 and glial function32,34 or blood-brain barrier homeostasis79. However, individual NCL and lysosomal-storage disorder-related proteins were identified, such as cathepsin D81, prosaposin82,83 and acid ceramidase84 confirming the veractity of the data generated.
By influencing the valine catabolic cascade through genetic disruption of HIBCH and HADHA it was possible to alter the CLN3 degenerative phenotype in Drosophila in vivo. Moreover, the further upstream in the catabolic cascade we intervened, the greater the resulting effect on disease phenotype. In humans, both HIBCH and HADHA deficiency cause infantile onset diseases with neurologic clinical scheme53,85,86. Crucially, this could indicate that perturbations in valine catabolic pathways might be a shared event across multiple neurodegenerative conditions and therefore targets identified here may be transferable to other diseases.
Interestingly, the upregulation of ROCK2 has been reported in a range of conditions by our laboratory and others2,61,62,64 (Fig. 7D). ROCK2 upregulation seemed to be a more synaptic-specific perturbation (at least at early stages) in the Cln3 −/− mice and its magnitude of change tracked across differentially vulnerable brain regions (i.e. Thalamus > Hippocampus > Cortex). Its genetic and/or pharmacological downregulation has also been shown to ameliorate the phenotype of several conditions including SMA, another relatively early onset neurodegenerative disease65,66. In this study we demonstrate for the first time that downregulation of ROCK2 also recues the phenotype in a CLN3 in vivo model when manipulated genetically, and to a lesser degree with available pharmacological agents (Fig. 7E–I).
Cortical synaptic extracts from human patients have shown that ROCK2 was downregulated in JNCL patients with respect to controls (Supplementary Fig. S9). Nevertheless, the interpretation of the results from post mortem tissue is challenging as at end stage of the disease brain atrophy is much more pronounced, making it difficult to distinguish which molecular alterations are more likely to be causative, and which are a consequence of the ongoing degeneration taking place. ROCK2 upregulation could be a disease stage-specific change that contributes to the initiation of degenerative stimuli, and it is not necessarily globally upregulated throughout disease progression. It would be advantageous to study the molecular alterations and pathways taking place throughout the time-course of disease progression in NCL as this would further our understanding of the mechanisms underlying the initiation and progression of neurodegeneration. In general, ROCK2 appears to be an attractive target for the development of new therapeutic strategies, not only for NCL, but also in a wide range of more common neurodegenerative diseases to potentially halt or delay disease progression (Fig. 7D). There are currently multiple ROCK inhibitors with different degrees of specificity available68,74. However, in order to avoid off-target effects in future studies, it will be necessary to develop a compound that targets ROCK2 more specifically.
It would be of interest for future investigations to study the molecular overlaps between valine degradation and ROCK2 signaling pathways contributing to synaptic vulnerability. In silico analysis based on published literature, suggests potential overlaps, although the level of complexity in the published data allowing us to draw links between these candidates would require carefully planned experiments in order to unravel such interactions (See Fig. S10).
We suggest that common mechanisms regulating the process of neurodegeneration and/or vulnerability of synapses may occur across a range of neurodegenerative conditions triggered by different insults. Unlike more common adult-onset complex polygenetic diseases such as Alzheimer or Parkinson disease for which the genetic insult triggering the neurodegeneration is not well understood, the NCLs are monogenetic diseases for which there are available well-characterized mice models that more accurately replicate the human disease14,31,32,35,87,88. Here we demonstrate that we can use the NCLs as a model to study a specific neurodegenerative process (i.e. synaptic loss) which is likely goverened by mechanisms which may indeed be conserved across multiple conditions ranging from injury through to chronic neurodegenerative conditions (i.e. Huntington disease and spinocerebellar ataxia) and diseases of childhood (i.e. SMA and the NCLs).
The workflows highlighted here should not be viewed as a mass screening “stamp collecting” exercise. Instead it should be clear that combining mammalian model “–omic” screening of differentially vulnerable tissues with in silico candidate identification and in vivo phenotypic assessment in Drosophila is an efficient pipeline for elucidating the mechanistic cascades governing neurodegenerative processes, and a “target-rich” way of identifying factors which are capable of modulating neurodegeneration, and therefore may be therapeutically targetable.
Materials and Methods
Tissue harvesting and processing
Ethics statement
All animal experiments were approved by a University of Edinburgh internal ethics committee and were performed under license by the UK Home Office (project license numbers 70/6567 and 70/7364).
Mice
Cln3 −/− mice31 inbred on a C57BL/6 background for at least 10 generations and control (+/+) littermates resulting from heterozygous crosses were used in this study. Mice were genotyped as described in31. Five Cln3 −/− and five wild-type control mice (WT) at 6.5 months and 12–13 months of age were used for immunohistochemistry experiment. Four Cln3 −/− and four control mice (WT) at 12–13 months were used for the proteomics and biochemical experiments.
Human post mortem samples
Human brain samples were obtained from the Human Brain and Spinal Fluid Resource Center, Los Angeles and from The MRC London Neurodegenerative Disease Brain Bank, Institute of Psychiatry, Psychology & Neuroscience, King’s College London. Samples were obtained at routine autopsy with fully informed written consent by the families. Study protocols for the use of this human material were approved by the Ethical Research Committees of the Institute of Psychiatry under the approval numbers 223/00 and 181/02. All experiments using human tissue samples and/or derivatives of, were performed in accordance with the relevant guidelines and regulations governing their storage, handling and disposal. Details about the samples are described in Supplementary Table S5.
Immunohistochemistry (IHC)
Five Cln3 −/− and five wild-type control mice (WT) at 6.5 months and 12-13 months of age were terminated and brains rapidly removed and immersed for fixation in 4% paraformaldhyde (pH 7.4). The IHC protocol used allows quantitative and qualitative comparisons between animal tissues as previously described in89,90. Please refer to Supplemental Experimental Procedures for more details.
Biochemical separation of neuronal compartments
Brains were harvested from six Cln3 −/− and wild-type (WT) controls mice at 13 months old and briefly chilled in ice cold ACSF (125 mM NaCl, 26 mM NaHCO3, 25 mM glucose, 2.5 mM KCl, 1.25 mM NaH2PO4, 1 mM CaCl2, 4 mM MgCl2) before regional microdissection. Thalamus, cortex (predominantly pre-/frontal) and hippocampus and were microdissected bilaterally12,17,91 pooled by mouse and immediately processed for “crude” synaptosome production as previously described2,17,92. Please refer to Supplemental Experimental Procedures for more details.
For the post mortem human samples, a portion of the total sample was obtained from frozen tissue (stored at −80 °C) and transferred in an eppendorf tube containing cold isotonic sucrose solution. Please refer to Supplemental Experimental Procedures for more details.
Protein extraction
Mice
For iTRAQ proteomics, protein extraction from “crude” synaptosomes was carried out as previously described in Fuller et al. 93. Please refer to Supplemental Experimental Procedures for more details.
Human
“crude” synaptosomes were resuspended and homogenized in a solution containing 100 mM Tris-HCl, 4% (w/v) SDS and 1% protease inhibitor cocktail (Roche). Samples were spun at 20,000 g and 4 °C for 20 minutes. Supernatant was aspirated and collected as extracted sample. Pellets and extracted samples were stored at −80 °C for QWB. Extracted samples concentration was determined using a BCA assay (Thermo).
Proteomic processing
Sample preparation and protein identification and quantification analysis by mass spectrometry was carried out as previously described in Fuller et al. 93. Please refer to Supplemental Experimental Procedures for more details.
Quantitative fluorescent western blotting (QWB)
QWB was performed as previously described in refs2,91,92,94,95. Briefly, samples were denatured in NuPage® LDS Sample buffer 4X (Invitrogen, UK) at 98 °C and 15ug of protein loaded and run on 4–12% Bis-Tris gel (Invitrogen). Accuracy of loading and protein estimation was confirmed by total protein analysis of Instant Blue (Expedeon) stained gels as previously described94. Protein transfer to a polyvinylidene fluoride (PDVF) membrane was carried out using the I-Blot® transfer system (Invitrogen, UK). Membranes were incubated with Odyssey blocking buffer (Li-Cor) for 30 minutes. Next, membranes were incubated in primary antibodies overnight at 4 °C and secondary antibodies for 1 h at room temperature. Please refer to Supplemental Experimental Procedures for more details.
In silico proteomic analysis
BioLayout Express 3D
BioLayout software incorporates a complex pattern recognition algorithm which groups protein data based only on expression profile. It allows the visualization and graphing of expression trends of co-expressed proteins to allow better characterization and understanding of complex large datasets44. Please refer to Supplemental Experimental Procedures for more details.
Enrichment analysis
To obtain an indication of the level of sample enrichment afforded through the process of synaptosome production, un-filtered mass spectrometry data was processed using The Database for Annotation, Visualization and Integrated Discovery (DAVID) software (available at http://david.abcc.ncifcrf.gov). DAVID provides a relatively comprehensive set of functional annotation tools for large data set interpretation96,97. DAVID was also used to characterize the functions associated to the protein clusters correlating to vulnerability mapped using BioLayout. For analysis to confirm synaptic protein enrichment we have applied a cut off of four fold as previously described in98 (see Table 1).
Ingenuity Pathway analysis
To obtain further insight into potential cellular pathways that may be perturbed in the Cln3 −/− thalamic pre-synaptic compartments compared to control mice, the Ingenuity Pathways Analysis (IPA) application (Ingenuity Systems) was used, as previously described2,91,92,99. Please refer to Supplemental Experimental Procedures for more details.
Phenotypic assessment methods and Drosophila husbandry
Drosophila mouse orthologs for candidate proteins were identified using DRSC Integrative Ortholog Prediction Tool (DIOPT)100 and RNAi lines were chosen from Flybase and obtained from Vienna Drosophila stock centre (VDSC) and from Bloomington Drosophila Stock Centre (BDSC) (see Table 2). UAS-CLN3 mutant fly was donated by Richard Tuxworth57. A stock carrying GMR-GAL4 and UAS-CLN3 on the second chromosome was established by conventional recombination methods and used as a tester line for the screen (GMR-GAL4;UAS-CLN3/CyO GFP). RNAi candidate lines were crossed to this line and the F1 progeny was assessed for suppression or enhancement of the CLN3-derived small and rough eye phenotype. To have a quantitative read out of the severity of the phenotypes obtained in our experiments we measured the eye surface area, as previously described101. Stocks were maintained on standard cornmeal food at room temperature. For all the crosses, flies were raised at 25 °C in a circadiarian light incubator. Details of the lines used can be found in Table 2.
Fasudil drug assay
The rock inhibitor used was Fasudil, Monohydrochloride Salt (LC laboratories, F-4660), which was previously used in mice models of SMA improving survival and skeletal muscle development65. GMR-GAL4 x UAS-CLN3 crosses were raised in Nutri-Fly instant food (Genesee Scientific, 66-118) containing 1.5 mM of Fasudil Monohydrochloride Salt and blue food colouring diluted in water, as described in102. Eyes of the offspring were photographed and total surface area was measured with Image J. Suppression of the CLN3-degenerative eye were calculated from 3 independent experiments as “(x- average eye surface area of the disease model)/(average of eye surface area of the control − average eye surface area of disease model) × 100” for each experiment.
Imaging
Immunohistochemistry
Micrographs were taken with Leica DMRB × 5/0.12 objective and AxioCam HRC Zeiss from the following brain regions: 1. The thalamic nuclei ventral posterior medial/ventral posterior lateral thalamic nucleus (VPM/VPL) and the dorsal lateral geniculate nucleus (LGNd); 2. Their respective cortical projections in the primary somatosensory cortex (S1BF) and primary visual cortex (V1) respectively, and 3; the hippocampal stratum radiatum and stratum oriens (See Fig. 1A). Immunoreactivity measurements were carried out using a thresholding image analysis in Image J (National Institute of Health, Bethesda, MD), USA), as previously described32,34,103. Thirty non-overlapping images (x40) were taken, on three consecutive sections, through the brain areas described above. All parameters including lamp intensity, video camera setup and calibration were kept constant throughout image capturing.
Drosophila eye images
For the HIBCH and HADHA experiments fruit fly eye images were photographed with a Nikon D5100 camera attached to a SZX9 Nikon stereomicroscope. For the ROCK2 experiments eye images were taken with an AxioCam ERc 5 s Rev.2 attached to a Zeiss Stemi 305 trino stereomicroscope. Images were analyzed with Image J software (National Institute of Health, Bethesda, MD, USA) as previously described in101.
Data analysis and figure production
QWB data was analysed using Odyssey software as per manufactures guidelines and as previously described2,17,104. Data was graphed and statistical comparisons for IHC thresholding, QWB protein intensity and Drosophila eye surface area quantification were carried out using GraphPad Prizm as previously described91,101. For QWB and IHC Student’s T test was applied. For Drosophila eye quantifications one-way ANOVA was carried out followed by Tukey’s multiple comparison test as a post-hoc when a significant difference was found in the ANOVA. P values < 0.05 were considered to be statistically significant for all analyses (*P < 0.05; **P < 0.01; ***P < 0.001). Statistical analysis of proteomic data was automatically carried out by MASCOT. P & Benjamini values to determine enrichment were automatically calculated by DAVID software (see above and http://david.abcc.ncifcrf.gov). P values/Fishers exact tests for pathway and networking analysis were automatically determined by IPA (see above and http://www.ingenuity.com/). Heatmap representing the differentially expressed proteins and pathways across brain regions was produced using “R” (http://www.R-project.org)105.
Availability of data and material
The raw datasets used and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Wishart, T. M., Parson, S. H. & Gillingwater, T. H. Synaptic vulnerability in neurodegenerative disease. J Neuropathol Exp Neurol. 65, 733–739 (2006).
Wishart, T. M. et al. Combining comparative proteomics and molecular genetics uncovers regulators of synaptic and axonal stability and degeneration in vivo. PLoS genetics 8, e1002936, https://doi.org/10.1371/journal.pgen.1002936 (2012).
DeKosky, S. T. & Scheff, S. W. Synapse loss in frontal cortex biopsies in Alzheimer’s disease: correlation with cognitive severity. Ann Neurol. 27, 457–464 (1990).
Selkoe, D. J. Alzheimer’s disease is a synaptic failure. Science 298, 789–791 (2002).
Meek P. D, M. K., Schumock GT. Economic considerations in Alzheimer’s disease. Pharmacotherapy. (2 Pt 2): 68–73; discussion 79-82 (1998).
Forno, L. S. Neuropathology of Parkinson disease. J. Neuropathol. Exp. Neurol 55, 259–272 (1996).
Fischer, L. R., Culver, D. G., Tennant, P., Davis, A. A. & Wang, M. Amyotrophic lateral sclerosis is a distal axonopathy: evidence in mice and man. Exp Neurol 185, 232–240 (2004).
Murray, L. M., Comley, L. H., Thomson, D., Parkinson, N. & Talbot, K. Selective vulnerability of motor neurons and dissociation of pre- and post- synaptic pathology at the neuromuscular junction in mouse models of spinal muscular atrophy. Human molecular genetics 17, 949–962 (2008).
Murphy, K. P., Carter, R. J., Lione, L. A., Mangiarini, L. & Mahal, A. Abnormal synaptic plasticity and impaired spatial cognition in mice transgenic or exon 1 of the human Huntington’s disease mutation. J Neurosci 20, 5115–5123 (2000).
Cunningham, C., Deacon, R., Wells, H., Boche, D. & Waters, S. Synaptic changes characterize early behavioural signs in the ME7 model of murine prion disease. Eur J Neurosci 17, 2147–2155 (2003).
Perkins, E. M., Clarkson, Y. L., Sabatier, N., Longhurst, D. M. & Millward, C. P. Loss of beta-III spectrin leads to Purkinje cell dysfunction recapitulating the behavior and neuropathology of spinocerebellar ataxia type 5 in humans. J Neurosci 30, 4857–4867 (2010).
Kielar, C. et al. Molecular correlates of axonal and synaptic pathology in mouse models of Batten disease. Human molecular genetics 18, 4066–4080, https://doi.org/10.1093/hmg/ddp355 (2009).
Virmani, T., Gupta, P., Liu, X., Kavalali, E. T. & Hofmann, S. L. Progressively reduced synaptic vesicle pool size in cultured neurons derived from neuronal ceroid lipofuscinosis-1 knockout mice. Neurobiol. Dis 20, 314–323 (2005).
Partanen, S. et al. Synaptic changes in the thalamocortical system of cathepsin D-deficient mice: a model of human congenital neuronal ceroid-lipofuscinosis. J. Neuropathol. Exp. Neurol. 67, 16–29 (2008).
Koch, S. et al. Morphologic and functional correlates of synaptic pathology in the cathepsin D knockout mouse model of congenital neuronal ceroid lipofuscinosis. Neuropathol Exp Neurol 70, 1089–1096 (2011).
Cooper, J. D. The neuronal ceroid lipofuscinoses: the same, but different? Biochemical Society transactions 38, 1448–1452, https://doi.org/10.1042/BST0381448 (2010).
Wishart, T. M. et al. Dysregulation of ubiquitin homeostasis and beta-catenin signaling promote spinal muscular atrophy. The Journal of clinical investigation 124, 1821–1834, https://doi.org/10.1172/JCI71318 (2014).
Powis, R. A. et al. Systemic restoration of UBA1 ameliorates disease in spinal muscular atrophy. JCI insight 1, e87908, https://doi.org/10.1172/jci.insight.87908 (2016).
Goebel, H. H. The neuronal ceroid-lipofuscinoses. J. Child. Neurol 10, 424–437 (1995).
Uvebrant, P. & Hagberg, B. Neuronal ceroid lipofuscinoses in Scandinavia. Epidemiology and clinical pictures. Neuropediatrics 28, 6–8 (1997).
Mole, S., Williams, R. & Goebel, H. H. The neuronal ceroid lipofuscinoses (Batten disease). 2nd ed. Oxford: Oxford University Press (2011).
Cooper, J. D. Progress towards understanding the neurobiology of Batten disease or neuronal ceroid lipofuscinosis. Current opinion in neurology 16, 121–128, https://doi.org/10.1097/01.wco.0000063762.15877.9b (2003).
Mole, S. E., Williams, R. E. & Goebel, H. H. Correlations between genotype, ultrastructural morphology and clinical phenotype in the neuronal ceroid lipofuscinoses. Neurogenetics 6, 107–126 (2005).
Cooper, J. D., Russell, C. & Mitchison, H. M. Progress towards understanding disease mechanisms in small vertebrate models of neuronal ceroid lipofuscinosis. Biochimica et biophysica acta 1762, 873–889, https://doi.org/10.1016/j.bbadis.2006.08.002 (2006).
Bond, M., Holthaus, S. M., Tammen, I., Tear, G. & Russell, C. Use of model organisms for the study of neuronal ceroid lipofuscinosis. Biochimica et biophysica acta 1832, 1842–1865, https://doi.org/10.1016/j.bbadis.2013.01.009 (2013).
The International Batten Disease Consortium. Isolation of a novel gene underlying Batten disease, CLN3. Cell 82, 949–957 (1995).
Pearce, D. A., Nosel, S. A. & Sherman, F. Studies of pH regulation by Btn1p, the yeast homolog of human Cln3p. Mol Genet Metab. 66, 320–323 (1999).
Luiro, K. et al. Interconnections of CLN3, Hook1 and Rab proteins link Batten disease to defects in the endocytic pathway. Human molecular genetics 13, 3017–3027, https://doi.org/10.1093/hmg/ddh321 (2004).
Hamilton, G. & Gillingwater, T. H. Spinal muscular atrophy: going beyond the motor neuron. Trends Mol Med. 19, 40–50 (2013).
Järvelä, I. et al. Clinical and magnetic resonance imaging findings in Batten disease: analysis of the major mutation (1.02-kb deletion). Ann Neurol. 42, 799–802 (1997).
Mitchison, H. M., Bernard, D. J. & Greene, N. Targeted disruption of the Cln3 gene provides a mouse model for Batten disease. The Batten Mouse Model Consortium. Neurobiol Dis. 6, 321–334 (1999).
Pontikis, C. C. et al. Late onset neurodegeneration in the Cln3−/− mouse model of juvenile neuronal ceroid lipofuscinosis is preceded by low level glial activation. Brain research 1023, 231–242, https://doi.org/10.1016/j.brainres.2004.07.030 (2004).
Kuhl, T. G., Dihanich, S., Wong, A. M. & Cooper, J. D. Regional brain atrophy in mouse models of neuronal ceroid lipofuscinosis: a new rostrocaudal perspective. Journal of child neurology 28, 1117–1122, https://doi.org/10.1177/0883073813494479 (2013).
Pontikis, C. C., Cotman, S. L., MacDonald, M. E. & Cooper, J. D. Thalamocortical neuron loss and localized astrocytosis in the Cln3Deltaex7/8 knock-in mouse model of Batten disease. Neurobiol Dis. 20, 823–836 (2005).
Kielar, C. et al. Successive neuron loss in the thalamus and cortex in a mouse model of infantile neuronal ceroid lipofuscinosis. Neurobiology of disease 25, 150–162, https://doi.org/10.1016/j.nbd.2006.09.001 (2007).
Santavuori, P., Vanhanen, S. L. & Autti, T. Clinical and neuroradiological diagnostic aspects of neuronal ceroid lipofuscinoses disorders. Eur J Paediatr Neurol. 5, 157–161 (2001).
Autti, T. et al. Thalami and corona radiata in juvenile NCL (CLN3): a voxel-based morphometric study. Eur J Neurol. 14, 447–450 (2007).
Wang, X., Michaelis, M. & Michaelis, E. Functional genomics of brain aging and Alzheimers disease: focus on selective neuronal vulnerability. Curr Genomics 11, 618–633 (2010).
Khan, U. et al. Molecular drivers and cortical spread of lateral entorhinal cortex dysfunction in preclinical Alzheimer’s disease. Nat Neurosci. 17, 304–311 (2014).
Braak, H. & Braak, E. Staging of Alzheimer’s disease-related neurofibrillary changes. Neurobiol Aging 16, 271–278 (1995).
Surmeier, D. J., Obeso, J. A. & Halliday, G. M. Selective neuronal vulnerability in Parkinson disease. Nature reviews. Neuroscience 18, 101–113, https://doi.org/10.1038/nrn.2016.178 (2017).
Lewandowski, N. M. et al. Regional vulnerability in Huntington’s disease: fMRI-guided molecular analysis in patients and a mouse model of disease. Neurobiology of disease 52, 84–93, https://doi.org/10.1016/j.nbd.2012.11.014 (2013).
Sieradzan, K. A. & Mann, D. M. A. The selective vulnerability of nerve cells in Huntington’s disease. Neuropathology and applied neurobiology 27, 1–21 (2001).
Theocharidis, A., van Dongen, S., Enright, A. J. & Freeman, T. C. Network visualization and analysis of gene expression data using BioLayout Express(3D). Nat. Protoc. 4, 1535–1550 (2009).
Enright, A. J., Van Dongen, S. & Ouzounis, C. A. An efficient algorithm for large-scale detection of protein families. Nucleic Acids Res. 30, 1575–1584 (2002).
Gillingwater, T. H. & Wishart, T. M. Mechanisms underlying synaptic vulnerability and degeneration in neurodegenerative disease. Neuropathology and applied neurobiology 39, 320–334, https://doi.org/10.1111/nan.12014 (2013).
Luiro, K. et al. Batten disease (JNCL) is linked to disturbances in mitochondrial, cytoskeletal, and synaptic compartments. Journal of neuroscience research 84, 1124–1138, https://doi.org/10.1002/jnr.21015 (2006).
Kang, S., Seo, J. H., Heo, T. H. & Kim, S. J. Batten disease is linked to altered expression of mitochondria-related metabolic molecules. Neurochem Int 62, 931–935 (2013).
Kolikova, J., Afzalov, R., Surin, A., Lehesjoki, A. E. & Khiroug, L. Deficient mitochondrial Ca(2+) buffering in the Cln8(mnd) mouse model of neuronal ceroid lipofuscinosis. Cell Calcium 50, 491–501 (2011).
Jolly, R. D., Brown, S., Das, A. M. & Walkley, S. U. Mitochondrial dysfunction in the neuronal ceroid-lipofuscinoses (Batten disease). Neurochemistry International 40, 565–557 (2002).
Loupatty, F. J. et al. Mutations in the gene encoding 3-hydroxyisobutyryl-CoA hydrolase results in progressive infantile neurodegeneration. American journal of human genetics 80, 195–199, https://doi.org/10.1086/510725 (2007).
Brown, G. K. et al. b- hydroxyisobutyryl coenzyme A deacylase deficiency: a defect in valine metabolism associated with physical malformations. Pediatrics 70, 532–538 (1982).
Ferdinandusse, S. et al. HIBCH mutations can cause Leigh-like disease with combined deficiency of multiple mitochondrial respiratory chain enzymes and pyruvate dehydrogenase. Orphanet J Rare Dis. 8, 188 (2013).
Kamijo, T., Aoyama, T., Komiyama, A. & Hashimoto, T. Structural analysis of cDNAs for subunits of human mitochondrial fatty acid β-oxidation trifunctional protein. Biochem. Biophys. Res. Commun. 199, 818–825 (1994).
Ibdah, J. A. et al. Lack of mitochondrial trifunctional protein in mice causes neonatal hypoglycemia and sudden death. J Clin Invest. 107, 1403–1409 (2001).
Fletcher, A. L., Pennesi, M. E., Harding, C. O., Weleber, R. G. & Gillingham, M. B. Observations regarding retinopathy in mitochondrial trifunctional protein deficiencies. Mol Genet Metab. 106, 18–24 (2012).
Tuxworth, R. I., Vivancos, V., O’Hare, M. B. & Tear, G. Interactions between the juvenile Batten disease gene, CLN3, and the Notch and JNK signalling pathways. Human molecular genetics 18, 667–678, https://doi.org/10.1093/hmg/ddn396 (2009).
Xu, Z.-S. Does a loss of TDP-43 function cause neurodegeneration? Molecular Neurodegeneration 7, 27, https://doi.org/10.1186/1750-1326-7-27 (2012).
Riento, K. & Ridley, A. J. Rocks: multifunctional kinases in cell behaviour. Nat Rev Mol Cell Biol 4, 446–456 (2003).
Zhou, Z., Meng, Y., Asrar, S., Todorovski, Z. & Jia, Z. A critical role of Rho-kinase ROCK2 in the regulation of spine and synaptic function. Neuropharmacology 56, 81–89, https://doi.org/10.1016/j.neuropharm.2008.07.031 (2009).
Deyts, C. et al. Dopamine D2 receptor stimulation potentiates PolyQ-Huntingtin-induced mouse striatal neuron dysfunctions via Rho/ROCK-II activation. PLoS One 4, e8287, https://doi.org/10.1371/journal.pone.0008287 (2009).
Herskowitz, J. H. et al. Rho kinase II phosphorylation of the lipoprotein receptor LR11/SORLA alters amyloid-beta production. The Journal of biological chemistry 286, 6117–6127, https://doi.org/10.1074/jbc.M110.167239 (2011).
Zhou, Y. et al. Nonsteroidal Anti-Inflammatory Drugs Can Lower Amyloidogenic AB42 by Inhibiting Rho. Science 302, 1215–1217 (2003).
Blaise, S. et al. In vivo evidence that TRAF4 is required for central nervous system myelin homeostasis. PLoS One 7, e30917, https://doi.org/10.1371/journal.pone.0030917 (2012).
Bowerman, M., Murray, L. M., Boyer, J. G., Anderson, C. L. & Kothary, R. Fasudil improves survival and promotes skeletal muscle development in a mouse model of spinal muscular atrophy. BMC medicine 10, 24, https://doi.org/10.1186/1741-7015-10-24 (2012).
Coque, E., Raoul, C. & Bowerman, M. ROCK inhibition as a therapy for spinal muscular atrophy: understanding the repercussions on multiple cellular targets. Frontiers in neuroscience 8, 271, https://doi.org/10.3389/fnins.2014.00271 (2014).
Takayasu, M. et al. The effects of HA compound calcium antagonists on delayed cerebral vasospasm in dogs. Journal of Neurosurgery 65, 80–85 (1986).
Olson, M. F. Applications for ROCK kinase inhibition. Curr Opin Cell Biol 20, 242–248 (2008).
Suzuki, Y. et al. Safety and efficacy of fasudil monotherapy and fasudil-ozagrel combination therapy in patients with subarachnoid hemorrhage: sub-analysis of the post-marketing surveillance study. Neurol Med Chir (Tokyo) 48, 241–248 (2008).
Masumoto, A. et al. Suppression of coronary artery spasm by the Rho-kinase inhibitor fasudil in patients with vasospastic angina. Circulation 105, 1545–1547 (2002).
Velat, G. J., Kimball, M. M., Mocco, J. D. & Hoh, B. L. Vasospasm after aneurysmal subarachnoid hemorrhage: review of randomized controlled trials and meta-analyses in the literature. World Neurosurg 6, 446–454 (2011).
Bharadwaj, S., Thanawala, R., Bon, G., Falcioni, R. & Prasad, G. L. Resensitization of breast cancer cells to anoikis by tropomyosin-1: role of Rho kinase-dependent cytoskeleton and adhesion. Oncogene 24, 8291–8303, https://doi.org/10.1038/sj.onc.1208993 (2005).
Hinsenkamp, I. et al. Inhibition of rho-associated kinase 1/2 attenuates tumor growth in murine gastric cancer. Neoplasia 18, 500–511, https://doi.org/10.1016/j.neo.2016.07.002 (2016).
Anastassiadis, T., Deacon, S. W., Devarajan, K., Ma, H. & Peterson, J. R. Comprehensive assay of kinase catalytic activity reveals features of kinase inhibitor selectivity. Nat. Biotechnol. 29, 1039–1045 (2011).
Luiro, K., K., O., Lehtovirta, M. & Jalanko, A. CLN3 protein is targeted to neuronal synapses but excluded from synaptic vesicles: new clues to Batten disease. Hum Mol Genet. 10, 2123–2131 (2001).
Phillips, S. N., Benedict, J. W., Weimer, J. M. & Pearce, D. A. CLN3, the protein associated with batten disease: structure, function and localization. Journal of neuroscience research 79, 573–583, https://doi.org/10.1002/jnr.20367 (2005).
Katz, M. L. et al. Immunochemical localization of the Batten disease (CLN3) protein in retina. Investigative Ophthalmology & Visual Science 38, 2375–2386 (1997).
Mohammed, A., O’Hare, M. B., Warley, A., Tear, G. & Tuxworth, R. I. in vivo localization of the neuronal ceroid lipofuscinosis proteins, CLN3 and CLN7, at endogenous expression levels. Neurobiology of disease 103, 123–132, https://doi.org/10.1016/j.nbd.2017.03.015 (2017).
Tecedor, L. et al. CLN3 Loss Disturbs Membrane Microdomain Properties and Protein Transport in Brain Endothelial Cells. The Journal of Neuroscience 33, 18065–18079, https://doi.org/10.1523/JNEUROSCI.0498-13.2013 (2013).
Tuxworth, R. I. et al. The Batten disease gene CLN3 is required for the response to oxidative stress. Human molecular genetics 20, 2037–2047, https://doi.org/10.1093/hmg/ddr088 (2011).
Steinfeld, R. et al. Cathepsin D Deficiency Is Associated with a Human Neurodegenerative Disorder. The American Journal of Human Genetics 78, 988–998, https://doi.org/10.1086/504159 (2006).
O’Brien, J. S. & Kishimoto, Y. Saposin proteins: structure, function, and role in human lysosomal storage disorders. FASEB J. 5, 301–308 (1991).
Tyynelä, J., Palmer, D. N., Baumann, M. & Haltia, M. Storage of saposins A and D in infantile neuronal ceroid-lipofuscinosis. FEBS Letters 330, 8–12, https://doi.org/10.1016/0014-5793(93)80908-D (1993).
Sugita, M., Dulaney, J. T. & Moser, H. W. Ceramidase Deficiency in Farber’s Disease (Lipogranulomatosis). Science 178, 1100–1102, https://doi.org/10.1126/science.178.4065.1100 (1972).
Peters, H. et al. Metabolite studies in HIBCH and ECHS1 defects: Implications for screening. Molecular genetics and metabolism 115, 168–173, https://doi.org/10.1016/j.ymgme.2015.06.008 (2015).
Stiles, A. R. et al. Successful diagnosis of HIBCH deficiency from exome sequencing and positive retrospective analysis of newborn screening cards in two siblings presenting with Leigh’s disease. Molecular genetics and metabolism 115, 161–167, https://doi.org/10.1016/j.ymgme.2015.05.008 (2015).
Cotman, S. L. et al. Cln3(Deltaex7/8) knock-in mice with the common JNCL mutation exhibit progressive neurologic disease that begins before birth. Hum Mol Genet. 11, 2709–2721 (2002).
Gupta, P. et al. Disruption of PPT1 or PPT2 causes neuronal ceroid lipofuscinosis in knockout mice. Proc. Natl Acad. Sci. USA 98, 13566–13571 (2001).
Bible, M., Lehesjoki, A. E., Jalanko, A., Cooper, J. D. & Kopra, O. Selective spatiotemporal patterns of glial activation and neuron loss in the sensory thalamocortical pathways of neuronal ceroid lipofuscinosis 8 mice. Neurobiol. Dis. 47, 444–457 (2012).
Tegelberg, S., Kopra, O., Joensuu, T., Cooper, J. D. & Lehesjoki, A. E. Early microglial activation precedes neuronal loss in the brain of the cstb−/− mouse model of progressive myoclonus epilepsy, EPM1. J. Neuropathol. Exp. Neurol 71, 40–53 (2012).
Wishart, T. M. et al. SMN deficiency disrupts brain development in a mouse model of severe spinal muscular atrophy. Human molecular genetics 19, 4216–4228, https://doi.org/10.1093/hmg/ddq340 (2010).
Wishart, T. M. et al. Differential proteomics analysis of synaptic proteins identifies potential cellular targets and protein mediators of synaptic neuroprotection conferred by the slow Wallerian degeneration (Wlds) gene. Mol Cell Proteomics 6, 1318–1330 (2007).
Fuller, H. R. et al. Valproate and bone loss: iTRAQ proteomics show that valproate reduces collagens and osteonectin in SMA cells. J Proteome Res. 9, 4228–4233 (2010).
Eaton, S. L. et al. Total protein analysis as a reliable loading control for quantitative fluorescent Western blotting. PLoS One 8, e72457 (2013).
McGorum, B. C. et al. Proteomic Profiling of Cranial (Superior) Cervical Ganglia Reveals Beta-Amyloid and Ubiquitin Proteasome System Perturbations in an Equine Multiple System Neuropathy. Mol Cell Proteomics 14, 3072–3086, https://doi.org/10.1074/mcp.M115.054635 (2015).
Huang, D. W., Sherman, B. T. & Lempicki, R. A. Systematic and integrative analysis of large gene lists using DAVID Bioinformatics Resources. Nature Protoc. 4, 44–57 (2009).
Huang, D. W., Sherman, B. T. & Lempicki, R. A. Bioinformatics enrichment tools: paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 37, 1–13 (2009).
Amorim, I. S. et al. Sideroflexin 3 is an α-synuclein-dependent mitochondrial protein that regulates synaptic morphology. Journal of Cell Science 130, 325–331, https://doi.org/10.1242/jcs.194241 (2017).
Savli, H., Szendröi, A., Romics, I. & Nagy, B. Gene network and canonical pathway analysis in prostate cancer: a microarray study. Exp Mol Med. 40, 176–185 (2008).
Hu, Y. et al. An integrative approach to ortholog prediction for disease-focused and other functional studies. BMC Bioinformatics 12, 357 (2011).
Sanhueza, M. et al. Network analyses reveal novel aspects of ALS pathogenesis. PLoS genetics 11, e1005107, https://doi.org/10.1371/journal.pgen.1005107 (2015).
Verspoor, R. L., Heys, C. & Price, T. A. Dyeing Insects for Behavioral Assays: the Mating Behavior of Anesthetized. J Vis Exp. 98 (2015).
Bible, E., Gupta, P., Hofmann, S. L. & Cooper, J. D. Regional and cellular neuropathology in the palmitoyl protein thioesterase-1 null mutant mouse model of infantile neuronal ceroid lipofuscinosis. Neurobiol. Dis. 16, 346–359 (2004).
Mutsaers, C. A., Lamont, D. J., Hunter, G., Wishart, T. M. & Gillingwater, T. H. Label-free proteomics identifies Calreticulin and GRP75/Mortalin as peripherally accessible protein biomarkers for spinal muscular atrophy. Genome Med. 5, 95 (2013).
R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria (2008).
Acknowledgements
The authors would like to thank Richard Tuxworth for the donation of the UAS-CLN3 as well as Mario Sanhueza and Daniel Sakovics-Matutes for their support carrying out the Drosophila experiments. Work in the authors’ laboratories has been supported by Institute Strategic Programme Grant Funding from the BBSRC (TMW), the Euan MacDonald Centre (TMW, MLH), The Darwin Trust of Edinburgh (MLH), the Batten Disease Support and Research Association (JDC) and the RJAH Institute of Orthopaedics (HRF). MLH is now supported by NCL-Stiftung.
Author information
Authors and Affiliations
Contributions
T.M.W. conceived and designed the study. M.L.H., H.R.F., A.M.S.W., S.L.E. and T.M.W. carried out experiments. T.M.W., and J.D.C. contributed reagents. M.L.H., J.D.C., T.H.G. and T.M.W. wrote the manuscript. G.P. contributed to the interpretation of the fly data. All authors contributed to the data analysis, interpretation and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Llavero Hurtado, M., Fuller, H.R., Wong, A.M.S. et al. Proteomic mapping of differentially vulnerable pre-synaptic populations identifies regulators of neuronal stability in vivo . Sci Rep 7, 12412 (2017). https://doi.org/10.1038/s41598-017-12603-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-12603-0
This article is cited by
-
Comparative proteomic profiling reveals mechanisms for early spinal cord vulnerability in CLN1 disease
Scientific Reports (2020)
-
Collateral Sprouting of Peripheral Sensory Neurons Exhibits a Unique Transcriptomic Profile
Molecular Neurobiology (2020)
-
Synapse alterations precede neuronal damage and storage pathology in a human cerebral organoid model of CLN3-juvenile neuronal ceroid lipofuscinosis
Acta Neuropathologica Communications (2019)
-
Comparative profiling of the synaptic proteome from Alzheimer’s disease patients with focus on the APOE genotype
Acta Neuropathologica Communications (2019)
-
Deep proteome profiling of the hippocampus in the 5XFAD mouse model reveals biological process alterations and a novel biomarker of Alzheimer’s disease
Experimental & Molecular Medicine (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
