
Abstract
Poor CD4+ T-cell recovery after cART has been associated with skewed T-cell maturation, inflammation and immunosenescence; however, T-cell functionality in those individuals has not been fully characterized. In the present study, we assessed T-cell function by assessing cytokine production after polyclonal, CMV and HIV stimulations of T-cells from ART-suppressed HIV-infected individuals with CD4+ T-cell counts >350 cells/μL (immunoconcordants) or <350 cells/μL (immunodiscordants). A group of HIV-uninfected individuals were also included as controls. Since CMV co-infection significantly affected T-cell maturation and polyfunctionality, only CMV+ individuals were analyzed. Despite their reduced and skewed CD4+ T-cell compartment, immunodiscordant individuals showed preserved polyclonal and HIV-specific responses. However, CMV response in immunodiscordant participants was significantly different from immunoconcordant or HIV-seronegative individuals. In immunodiscordant subjects, the magnitude of IFN-γ+ CD8+ and IL-2+ CD4+ T-cells in response to CMV was higher and differently associated with the CD4+ T-cell maturation profile., showing an increased frequency of naïve, central memory and EMRA CMV-specific CD4+ T-cells. In conclusion, CD4+ and CD8+ T-cell polyfunctionality was not reduced in immunodiscordant individuals, although heightened CMV-specific immune responses, likely related to subclinical CMV reactivations, may be contributing to the skewed T-cell maturation and the higher risk of clinical progression observed in those individuals.
Similar content being viewed by others

Introduction
Combination antiretroviral therapy (cART) with effective control of viral replication and subsequent immunologic reconstitution has dramatically improved the health of HIV-infected individuals, resulting in a reduction in HIV-related morbidity and mortality1. However, despite persistent virus suppression, about 15–30% of treated HIV-infected individuals fail to achieve optimal CD4+ T-cell reconstitution, referred as immunological non-responders or immunodiscordant individuals2,3. Several factors have been associated with a poor CD4+ T-cell immune recovery (reviewed in ref.4), among others altered thymic production5,6, low nadir CD4 counts7, older age8, high levels of immune activation5,7,9 and increased cell death5,7. Additionally, immunodiscordant individuals show a skewed T-cell maturation profile10,11,12,13, increased expression of markers of replicative senescence (CD28+CD57+)6,13,14 and high frequencies of programmed cell death protein-1 (PD-1)-expressing CD4+ T-cells5,15, a phenotype associated with immune exhaustion, and defined by loss of effector functions and proliferative capacity. However, it is unclear how these changes affect the functional diversity (i.e. polyfunctionality) of CD4+ and CD8+ T-cells in immunodiscordant individuals.
Cytomegalovirus (CMV) infection in healthy individuals is usually asymptomatic and results in latent infection. CMV co-infection is highly prevalent in the HIV-infected population (between 75 and 100%)16 and episodes of CMV-reactivation are increased, affecting morbidity and mortality17. CMV infection is also associated with significant changes in the composition of the T-cell repertoire, accelerated T-cell immunosenescence and immune exhaustion18,19. In particular, CMV has been described as a major contributor to the increased immune activation and senescence in HIV+ individuals with poor CD4+ T-cell recovery20,21,22. Furthermore, increased CMV-specific antibodies and/or T-cells have been associated with atherosclerosis and impaired CD4+ T-cell reconstitution and progression in HIV-infected individuals on treatment23,24,25,26,27. However, CMV-specific T-cell responses in individuals with poor CD4+ T-cell recovery have not been completely characterized.
We hypothesized that skewed CD4+ T-cell maturation and increased exhaustion could be factors contributing to an impaired T-cell polyfunctionality in immunodiscordant individuals. Therefore, in the present study we analyzed cellular immune response of CMV-seropositive HIV-infected individuals with different CD4+ T-cell recovery upon virologically suppressive cART. The frequency, functional capacity and differentiation profile of CD4+ and CD8+ T-cells after PMA and ionomycin, CMV and HIV stimulation was evaluated.
Results
Participant characteristics
A total of 43 HIV-infected individuals were included: 25 participants were classified as immunoconcordants and 18 as immunodiscordants (Table 1). Both HIV-infected groups were similar in age, gender, prevalence of HCV, time since diagnosis and treatment conditions (Table 1). As per inclusion criteria, significantly lower absolute CD4+ T-cell counts were observed in the immunodiscordant group than in the immunoconcordant group. In addition, also lower nadir CD4+ T-cell counts and CD8+ T-cell counts were observed in the immunodiscordant group. Although not significant, a higher proportion of CMV-seropositive (CMV+) individuals were found in the HIV-infected group than in the HIV-uninfected control group. None of the participants had detectable CMV viral load in urine samples as assessed using quantitative CMV-PCR.
Impact of CMV infection on T-cell maturation and polyfunctionality
We observed a significant impact of CMV serostatus in the phenotypic distribution of CD4+ and CD8+ T-cells (p = 0.04 and p = 0.01, respectively, Fig. 1A and gating strategy in Figure S1). CMV-seropositive HIV-uninfected individuals showed decreased naïve cells and increased CD27− subsets (TEM and TEMRA cells) than CMV-seronegative. Although a trend was seen, there was no significant difference in the age between CMV− and CMV+ populations (median (IQR): 38 (36–45) and 46 (41–54), respectively p = 0.07). Moreover, significant differences in the polyfunctional profile after PMA/Ionomycin stimulation of CD4+ T-cells (p = 0.049) was also observed between CMV-seropositive and CMV-seronegative HIV-uninfected individuals (Fig. 1B). Phenotypic and functional changes by CMV infection in the HIV+ ART-treated population could not be analyzed because the low number of HIV+CMV− individuals recruited. However, to avoid the bias induced by the potential immunological impact of CMV infection, only CMV-seropositive individuals were included for the rest of the analyses. Table 2 summarizes the main characteristics of CMV-seropositive individuals. Unlike the whole cohort (Table 1) no differences in age or CD4+ T-cell counts were observed between HIV-uninfected and HIV-infected CMV-seropositive groups. No other differences were observed between CMV-seropositive populations (Table 2) in comparison with the whole cohort (Table 1).

Phenotypic and functional changes induced by CMV infection. (A) CD4+ and CD8+ T-cell differentiation profile in HIV-uninfected CMV-seropositive and -seronegative individuals. The differential expression of CD45RA, CCR7 and CD27 by CD4+ and CD8+ T-cells was analyzed by boolean gating. Pie charts illustrate relative proportions of each of the different subsets in CMV-seropositive and CMV-seronegative. (B) CD4+ and CD8+ T-cell responses to PMA/ionomycin stimulation in HIV-uninfected individuals stratified by CMV serostatus. Polyfunctional profiles of CD4+ and CD8+ T-cells responding to PMA/ionomycin are showed. Detail of the 3-function combinations (IFN-γ, IL-2 and TNF-α) of CD4+ and CD8+ T-cells responding to PMA/ionomycin is depicted on the bottom of the figure. Statistical comparisons of the profiles were performed by partial permutation tests, using Spice software (*p < 0.05, **p < 0.01). HIV−: HIV-uninfected individuals; CMV−: CMV-seronegative individuals; CMV+: CMV-seropositive individuals.
Polyfunctional profile of CD4+ and CD8+ T-cell responses according to CD4+ T-cell recovery
To evaluate the T-cell functional diversity in the three analyzed groups (HIV-negative, HIV+ immunoconcordant and HIV+ immunodiscordant), frequencies of CD4+ and CD8+ T-cells that expressed IFN-γ, IL-2 and TNF-α following the strong non-specific stimulation with PMA/ionomycin were evaluated (Fig. 2). CD4+ T-cells from immunodiscordant subjects secreted slightly higher, but not significant total levels of IFN-γ and similar IL-2 and TNF-α levels than HIV-negative and immunoconcordant individuals (Fig. 2A, Panel CD4). Consistently, no significant differences between the three groups were observed when the polyfunctional index (Fig. 2B, Panel CD4) or the SPICE polyfunctional profile (Fig. 2C, Panel CD4) was compared among groups. However, in a more detailed analysis, the immunodiscordant group showed an increase in all the IFN-γ+ subsets, reaching statistical significance in the bifunctional subset displaying IFN-γ and IL-2 and the subset producing IFN-γ alone (Fig. 2D, Panel CD4). Conversely, a lower proportion of the monofunctional IL-2+ subset was found in the immunodiscordant group compared with HIV-uninfected individuals (Fig. 2D, Panel CD4).

CD4+ and CD8+ T-cell responses to PMA/ionomycin stimulation. (A) The frequency of CD4+ and CD8+ T-cells positive for each cytokine (IFN-γ, IL-2 and TNF-α) is represented (mean and SEM). (B) Polifunctionality index (PI) of HIV-uninfected individuals, immunoconcordant and immunodiscordant individuals (Mean and SEM are represented). (C) Pie charts representing the polyfunctional profile of CD4+ and CD8+ T-cells from HIV-uninfected individuals, immunoconcordant and immunodiscordant individuals. Arcs depict cytokine makeup within pie slice. (D) Dot graphs indicate percentage of responding cells based on different combinations of cytokine function. The combination of functions studied (pie slice) is indicated in the graph below (Median and IQR are represented). Nonparametric Mann-Whitney U test was used to analyze differences between groups (*p < 0.05, **p < 0.01). HIV−: CMV+ HIV-uninfected individuals; HIV+: CMV+ HIV-infected ART-treated individuals; IC: CMV+ immunoconcordant individuals; ID: CMV+ immunodiscordant individuals.
Likewise, immunodiscordant individuals showed a higher, although no significant, proportion of total CD8+ T-cells producing IFN-γ in comparison with immunoconcordant individuals (p = 0.05) with no differences in the polyfunctional index or in the SPICE polyfunctional CD8+ T-cell profile between groups (Fig. 2, Panel CD8). All these data suggest that, after a global stimulation, the CD4+ and CD8+ T-cell polyfunctionality profile is preserved in immunodiscordant individuals compared with immunoconcordant and HIV-negative individuals, although a skewed response towards high IFN-γ production could be observed in immunodiscordant individuals.
Functional T-cell responses mediated by IFN-γ and IL-2 against CMV and HIV
To further investigate the functional properties of the different groups of individuals, CMV-specific immune responses (IFN-γ and IL-2 production) were analyzed. Three different CMV antigens were used: a whole CMV viral lysate, containing a high diversity of T-cell epitopes, a pp65 peptide pool, and the pp65 recombinant protein. A similar pattern of total IFN-γ+ and IL-2+ CMV-specific CD4+ and CD8+ T-cells was observed in response to all the CMV antigens used, although the magnitude was different between antigens in the two T-cell populations (Fig. 3). As expected, the IFN-γ response was higher in CD8+ T-cells than in CD4+ T-cells after stimulation with all the stimuli and the proportion of IFN-γ + cells, in both CD4+ and CD8+ T-cells, exceeded that of IL-2+ cells. Between groups, a tendency to higher total IFN-γ-secreting cells in the immunodiscordant group was observed, although only in CD8+ T-cells stimulated with CMV viral lysate reached significant differences in comparison with the HIV-negative group (Fig. 3). The proportion of total IL-2+ pp65 peptide pool-specific CD4+ T-cells was significantly increased in both immunoconcordant and immunodiscordant individuals in comparison with the HIV-negative group. IL-2+ cells were also significantly higher in immunodiscordant than in immunoconcordant individuals after CMV viral lysate stimulation. In CD8+ T-cells, an increase of IL-2+ cells was observed in immunoconcordant individuals in comparison to both immunodiscordant and the HIV-negative group in presence of CMV pp65 peptide pool. Furthermore, an association between cellular and humoral CMV-specific immune responses was found. Higher CMV IgG levels were observed in immunodiscordant individuals with a significant association between IL-2+ CMV-specific CD4+ T-cell response and the anti-CMV IgG titers (Spearman r = 0.34; p = 0.03) (data not shown).

Functional CD4+ and CD8+ T-cell responses to CMV and HIV antigens. PBMCs were stimulated with different CMV antigenic preparation and the HIV p24 recombinant protein. The specific CD4+ and CD8+ T-cell responses were measured by flow cytometry in CMV+ HIV-uninfected individuals (blue bars, n = 12), CMV+ immunoconcordant individuals (green bars, n = 17 and n = 6 for CMV antigens and p24, respectively) and CMV+ immunodiscordant individuals (red bars, n = 12 and n = 8 for CMV antigens and p24, respectively). Response magnitudes are reported as the percentages of CD4+ and CD8+ T-cells producing IFN-γ and IL-2 after background subtraction. Bars indicate mean values ± SEM. Individual data of all subjects are represented by dots. Differences were tested using Mann-Whitney U nonparametric test (*p < 0.05).
Regarding HIV-specific responses, after stimulation with the HIV-1 p24 recombinant protein, the mean frequency of HIV-specific CD4+ T-cells was lower than the CMV-responses and no differences were observed between immunoconcordant and immunodiscordant individuals (Fig. 3). The CD8+ T-cell response to the HIV p24 recombinant protein appeared to be negligible with a mean frequency of 0.016 of the total CD8+ subset responding to the protein. Thus, the results obtained by the analysis of distinct CMV Ag-specific responses indicated that an increased CMV-specific response could be observed in HIV-positive individuals and the response was highest in immunodiscordant individuals.
Functional analysis of different memory CD4+ and CD8+ T-cell subsets
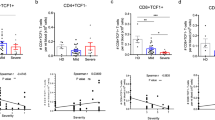
In our study, the overall profile of CD4+ and CD8+ T-cell maturation was significantly different between immunodiscordant and immunoconcordant individuals, with a significant decrease in the percentage of both naïve CD4+ and CD8+ T-cells and an increase in different memory subsets (Figure S2). To determine whether this disturbed maturational profile is also accompanied by changes in functionality, we next evaluated the capacity to produce IFN-γ and IL-2 after CMV or HIV p24 stimulation of different memory CD4+ and CD8+ T-cell subsets. Remarkably all memory CD4+ T-cell subsets of immunodiscordant individuals were functional, able to produce IFN-γ and IL-2 after CMV stimulation. Moreover, increased proportions of several memory CD4+ T-cell subsets IFN-γ + and IL-2+ were observed in immunodiscordant compared to immunoconcordant and HIV-uninfected controls, including CD4+ T-cells with a naïve phenotype, CD4+ TCM and CD4+ TEMRA early cells (Fig. 4). The same differences were found with all the CMV preparations used. No significant changes were observed in the CD8+ T-cell population (data not shown).

Functionality of CMV and HIV-specific memory CD4+ and CD8+ T-cell subsets. The differential expression of CD45RA, CCR7 and CD27 by CD4+ and CD8+ T-cells was analyzed by boolean gating. Based on the expression of these surface markers we were able to discriminate eight different subpopulations expressing each possible combination of markers: naïve (TN, CD45RA+CCR7+CD27+), central memory (TCM, CD45RA−CCR7+CD27+), early and late effector memory (TEM early, CD45RA−CCR7-CD27+ and TEM late, CD45RA−CCR7−CD27−), and early and late effector memory re-expressing CD45RA T-cells (TEMRA early, CD45RA+CCR7−CD27+ and TEMRA late, CD45RA+CCR7−CD27−). Other intermediate phenotypes (CD45RA+CCR7+CD27− and CD45RA−CCR7+CD27−), which can not as yet be ascribed to a specific subpopulation or to a functionally unique subset, observed in low percentages are not shown. The frequency of IFN-γ+ and IL-2+ CMV and HIV-specific T-cells across distinct subsets are shown. CMV+ HIV-uninfected individuals (blue bars, n = 12), CMV+ immunoconcordant individuals (green bars, n = 16 and n = 6 for pp65 peptide pool and p24, respectively) and CMV+ immunodiscordant individuals (red bars, n = 11 and n = 8 for pp65 peptide pool and p24, respectively). The median and tukey ranges are shown for each group. Differences were tested using Mann-Whitney U nonparametric test (*p < 0.05, **p < 0.01).
Several memory CD4+ T-cell subsets produce IFN-γ and IL-2 after p24 stimulation in both immunoconcordant and immunodiscordant individuals, with higher frequency of CD4+ HIV-specific T-cells producing IL-2 with a naïve phenotype observed in immunodiscordant individuals compared with immunoconcordant individuals (Fig. 4).
Association between the magnitude of CMV-specific T-cell responses and T-cell maturation profile
As shown in Fig. 1, CMV-seropositivity negatively impacts on the proportion of naïve T-cells and positively impacts on the frequency of effector memory T-cells in HIV-uninfected individuals. Accordingly, in HIV-uninfected individuals we observed a strong negative correlation between the CMV-total response (defined as the mean of the sum of the IFN-γ and IL-2 frequencies from different CMV antigens) and the proportion of CD4+ naïve T-cells (Spearman r = -0.9; p < 0.0001) and a positive correlation with the proportions of both CD4+ early and late TEM cells (Sperman r = 0.87; p < 0.0001 and Spearman r = 0.78; p = 0.002) (Fig. 5). In contrast, this clear effect seems to be perturbed in HIV+ individuals. Indeed, no correlation between the CMV-total response and the proportion of naïve CD4+ T-cells in the HIV-suppressed population was found. The correlation with the proportion of CD4+ TEM early cells was also lost in immunoconcordant individuals and was reversed in immunodiscordant individuals showing a significant negative value (Sperman r = −0.61; p = 0.03). Analysis of other memory subsets showed a significant positive correlation with the frequency of CD4+ TEM late T-cells (Spearman r = 0.61; p = 0.02) and a trend with the proportion of CD4+ TEMRA late T-cells (Spearman r = 0.55; p = 0.05) was observed (Fig. 5). No associations were observed between the CMV-response and the composition of the CD8+ T-cell compartment in any of the studied population.

Associations between total CMV-specific CD4+ T-cell response and CD4+ T-cell subsets. Correlations between total CMV-specific response (Median of total response IFN-γ and IL-2 production of all CMV antigens used) and CD4+ T-cell subsets are represented. CMV+ HIV-uninfected individuals (blue dots, n = 13), CMV+ immunoconcordant individuals (green dots, n = 18) and CMV+ immunodiscordant individuals (red dots, n = 13). Linear correlation (Spearman) r and p-values are shown.
Characterization of CMV and HIV-specific CD4+ and CD8+ T-cell differentiation profile
Finally, an analysis of the differentiation profile of CMV-specific CD4+ and CD8+ T-cells and HIV-specific CD4+ T-cells was performed. No difference between the patterns of response to the pp65 peptide pool and the pp65 protein were observed, therefore only the responses to the pp65 protein, the CMV viral lysate and the HIV-1 p24 recombinant protein are shown (Fig. 6 and Figure S3 for IFN-γ and IL-2, respectively). CMV-specific IFN-γ-producing CD4+ T-cells in HIV-negative individuals showed a similar phenotypic pattern of responses with all the stimuli used, with most of the cells displaying a TEM phenotype with an equal response of TEM early and TEM late cells (Fig. 6). Conversely, in HIV-infected individuals a different pattern of response was observed with the different CMV antigenic preparations. The differentiation pattern of the viral lysate-specific CD4+ cells among HIV+ individuals, in both immunoconcordant and immunodiscordant groups, was significantly skewed towards TEM late cells compared with the differentiation pattern of pp65 protein-specific IFN-γ+ cells. Similarly, differences were found in the profile of CMV-specific CD8+ IFN-γ + cells (Fig. 6) and in CD4+ and CD8+-specific IL-2+ T-cells responding to different CMV stimuli and among groups (Figure S3). However, it is important to note that no significant differences were found between immunodiscordant and immunoconcordant individuals with any of the CMV antigens (Fig. 6 and Figure S3). Regarding HIV-specific response, the differentiation profile of both IFN-γ+ and IL-2+ HIV-specific CD4+ T-cells was significantly different to that of CMV, but no differences were observed between immunoconcordant and immunodiscordant individuals. Therefore, immunodiscordant subjects despite displaying a skewed T-cell maturation profile do not present alterations in the differentiation pattern of either CMV- or p24-specific T-cells in comparison with immunoconcordant individuals.

Phenotypic profile of IFN-γ + virus-specific CD4+ and CD8+ T-cells. IFN-γ+ CMV- and HIV-specific CD4+ and CMV-specific CD8+ T-cells were determined after stimulation with different CMV antigens and the HIV p24 recombinant protein. The memory maturation profile of the IFN-γ+ specific cells was examined for CD45RA, CCR7 and CD27 expression by using a boolean gated strategy. The distribution of CMV- and HIV-specific IFN-γ+ response among different subsets was represented using SPICE software. The phenotypic patterns are color-coded and indicated. Significant differences between antigens are depicted in different colors: blue for CMV+ HIV− individuals, green for CMV+ immunoconcordant (IC) and red for CMV+ immunodiscordant (ID). Significant differences between groups (intra-antigen) are also represented (black lines). Statistical testing by permutation performed with SPICE software (*p < 0.05, **p < 0.01, ***p < 0.001).
Discussion
This study provides an analysis of T-cell functionality in a group of HIV-infected individuals with different CD4+ T-cell recovery upon virologically suppressive cART. Our data show that even thought immunodiscordant individuals have a skewed T-cell maturation profile10,11,12,13, T-cell polyfunctionality, assessed by cytokine production after polyclonal and HIV-specific stimulation, is preserved in those individuals. In addition, we have observed that the magnitude of CMV-specific response was differentially associated with state of CD4+ T-cell maturation in immunodiscordant and immunoconcordant or HIV-negative groups, likely related to a more frequent or intense subclinical CMV reactivation from latency in immunodiscordant individuals (Gómez-Mora et al. PLOS ONE. Accepted Manuscript).
CMV infection, in addition to shape the naïve and memory T-cell repertoire28,29,30,31, appears to be associated with accelerated T-cell immunosenescence and immune exhaustion, both phenotypes associated with reduced T-cell functionality18,19. These CMV-induced phenotypic and functional changes were corroborated in our HIV-uninfected population, with higher level of differentiated CD4+ and CD8+ T-cells and differences regarding cytokine production in CMV-seropositive individuals compared with CMV-seronegative individuals. Therefore, CMV-seronegative individuals were excluded to avoid the bias in the analysis of functionality of mixed populations (CMV-seropositive and CMV-seronegative individuals) and should be taken into account in future phenotypic and functional characterizations. Indeed, the phenotypic characterization of immunodiscordant individuals, in accordance with our previous reports, showed a decrease in the percentages of both naïve CD4+ and CD8+ T-cells and an increased frequencies of mature T-cells in comparison with the immunoconcordant group, although the differences were lower than in our previous studies7,13 likely due to the exclusion of the CMV-seronegative individuals in the present study. Differences with the HIV-uninfected group were also found, however we cannot exclude that the difference with this group, mainly the decrease in naïve T-cells, could be also associated with age. The HIV-infected population was slightly older than the HIV-uninfected population, although the difference was no statistically significant.
After the non-specific polyclonal stimulation with PMA/ionomycin, a robust production of IFN-γ, IL-2 and TNF-α was displayed in all individuals. Remarkably, CD4+ and CD8+ T-cell polyfunctionality was not reduced in immunodiscordant individuals compared with immunoconcordant and HIV-negative individuals suggesting that the homeostatic alterations and the increased replicative senescence or exhaustion observed in immunodiscordant individuals per se may not be linked to reduced polyfunctionality. A similar paradox has been reported in healthy older people, who maintain polyfunctionality despite phenotypic alterations, as compared to younger people32,33. Quantitatively, immunodiscordant individuals showed a significant enrichment in IFN-γ-producing CD4+ and CD8+ T-cells and a decrease in monofunctional IL-2-producing T-cells as compared with immunoconcordant and HIV-negative individuals. In line with this data, it has been recently reported that the decrease of mRNA level of IFN-γ after maraviroc administration was associated with the increase in CD4+ T-cells in subjects with poor CD4 T-cell recovery34. Furthermore, CD8 T cells producing C-C motif chemokine ligand 4 (CCL-4), CD107a and IFN-γ were negatively associated with immune reconstitution35. A potential cause could be the disturbed maturational profile displayed by the immunodiscordant individuals; the enrichment in highly-differentiated cells and the low proportions of naïve T-cells as IFN-γ is primarily produced by differentiated memory cells and IL-2 by naïve and central memory T-cells36. Potential consequences of the alteration in IFN-γ production could be associated with its polyfunctional effects on immune activation and pro-inflammatory responses by sustaining the chronic immune activation and the apoptosis sensitivity associated with HIV infection (reviewed in ref.37). On the other hand, IL-2 is a central regulator of T-cell proliferation, activation and differentiation38, thus, slight differences in IL-2 production, also reported by others12,39, might be contributing to the poor immune recovery and skewed T-cell maturation observed in immunodiscordant subjects.
CMV-specific T-cell responses are higher in HIV-infected individuals than in HIV-uninfected individuals, particularly those receiving cART23,40. In our study, HIV-infected subjects showed a tendency towards higher proportions of CMV-specific IFN-γ and IL-2 secreting CD4+ and CD8+ T-cells than HIV-uninfected individuals. In addition, we also found changes in the differentiation profile of CMV-specific T-cells between HIV-infected and uninfected individuals stimulated with viral lysate. All these data suggest that HIV-infected individuals could have a different pattern of CMV protein expression, maybe relate to recurrent episodes of subclinical viral reactivation41 and which could result in higher CMV-specific responses and a change of CD4+ T-cell immunodominance. In the HIV-infected population, the highest CMV-specific T-cell responses were found in immunodiscordant individuals in line with previous work reporting that strong anti-CMV responses are associated with lower CD4+ T-cell counts24. Moreover, in immunodiscordant individuals the increase in the size of the T-cell response to CMV was associated with an increased proportion of several CD4+ T-cell memory subsets responding to CMV having also an impact in the entire memory CD4+ T-cell subset distribution. Consistent with a previous report42 a correlation between the degree of T-cell differentiation and the size of the CMV-response was observed in HIV-uninfected individuals. However, in contrast with immunoconcordant individuals in whom CMV infection does not seem to have an important role in the CD4+ T-cell distribution, in immunodiscordant individuals CMV infection could be involved in the observed skewed T-cell maturation.
The heightened T-cell response against CMV observed in immunodiscordant individuals is not necessarily beneficial. In our study, an association between CMV-specific T-cell response and anti-CMV IgG antibodies was observed with higher level of anti-CMV IgG antibodies in immunodiscordant individuals (Gómez-Mora et al., PLOS ONE. Accepted Manuscript) and in line with a recent study, where an association between CMV IgG levels and disease progression has been reported27. It has been demonstrated that circulating anti-CMV IgG antibodies and CMV-specific T-cell responses are associated with cardiovascular diseases and physical function impairment23,25,26,43, suggesting that in HIV+ individuals with poor CD4+ T-cell recovery the high CMV-specific response, probably due to episodes of CMV reactivation mainly in the genital tract41,44, may have an impact in the higher mortality and morbidity observed in those individuals.
As expected, when compared to CMV, low HIV-1-specific CD4+ T-cells were detected in most ART-suppressed individuals45, with a distinct differentiation profile46,47,48. It has been previously shown that individuals with poor CD4+ T-cell recovery have reduced Gag-specific IFN-γ ELISPOT response11 and poor CD8+ lymphoproliferative responses to Gag39. However, a higher IL-2-secretion after gp120-stimulation by CD4 T-cells in individuals with a low-level CD4 T-cell repopulation has been also reported49. Using a p24 protein, we did not find any difference in the total IFN-γ+ or IL-2+ HIV-specific immune response between immunoconcordant and immunodiscordant individuals. Surprisingly we found increased frequencies of IL-2+ HIV-specific CD4+ T-cells with a naïve phenotype in immunodiscordant individuals. It has been reported that higher frequencies of HIV Gag-specific IFN-γ+IL-2+ CD4+ T-cells are strongly associated with higher cell-associated HIV DNA levels in HIV controllers50 and CMV reactivation has been associated with higher levels of HIV DNA in blood cells44. In cART-treated HIV-positive individuals a trend for association between HIV-specific CD4+ T-cells expressing IL-2 and the total HIV DNA in resting CD4+ T-cells has also been described51. Therefore, the higher levels of proviral DNA that have been found in immunodiscordant individuals52 could be associated with a higher percentage of IL-2+ HIV-specific CD4+ T-cells, which are more likely to be infected with HIV53 and related to CMV infection as previously suggested44. The use of a protein instead of a pool of peptides only allowed us to evaluate the HIV-specific CD4+ T-cell responses, so we can not rule out that there are also differences in the IFN-γ production by CD8+ T-cells between immunodiscordant and immunoconcordant individuals.
One limitation of this study is the low number of individuals analyzed, especially for the study of HIV-specific responses. In addition, polyfunctionality (measured by the expression of IFN-γ, IL-2 and TNF-α) was only determined following the non-specific stimulation. A polyfunctional profile, including cytolytic markers as perforin and granzyme B and the degranulation marker (CD107), have not been analyzed and it has been demonstrated that the highly polyfunctional antigen-specific cells are the most potent effectors and that could be correlated with clinical outcome54,55,56.
Taken together, our results indicate that HIV-infected ART-suppressed individuals with poor immune recovery displayed a maintained global T-cell polyfunctionality despite having severe homeostatic alterations and increased levels of replicative senescence and exhaustion. Furthermore, in immunodiscordant individuals, asymptomatic CMV infection, that has been previously described as a major contributor to the increased immune activation and senescence resulted in a heightened CMV-specific immune response that may be contributing to the skewed T-cell maturation and the higher risk of clinical progression observed in those individuals. Future studies are needed to determine if persistent CMV replication could be targeted as a strategy to reduce the size of specific immune response and reduce the CMV-associated immune pathology.
Methods
Subjects
A total of 43 HIV-infected individuals were recruited for the study. Inclusion criteria for all participants were confirmed diagnosis of HIV infection, continuous cART with sustained undetectable HIV-1 RNA (plasma viral load < 50 copies/ml) for at least the past 2 years (minimum of four determinations during this time period) and good antiretroviral treatment (ART) adherence. Individuals were classified as immunoconcordant (favorable virologic and immunologic response) when CD4+ T-cell counts were above 400 cells/µL and as immunodiscordant individuals (favorable virologic response but unsatisfactory immunologic response) if CD4+ T-cell counts were persistently less than 350 cells/µL. For comparative purposes a control group of 21 HIV-uninfected individuals was also analyzed. The institutional review board of the Hospital Germans Trias i Pujol approved the study (EO code: EO-07-024). The methods were carried out in accordance with the Declaration of Helsinki. Written informed consent was obtained from all participants.
Determination of anti-CMV IgG levels
The CMV-specific IgG antibodies were measured using the semi-quantitative chemiluminiscent immunoassay (Synlab Diagnostics, Barcelona, Spain). Samples determined as <12U/ml were considered negative, determinations between 12 and 14U/ml were undetermined, and >14U/ml were considered positive samples.
CMV viral load detection by RT-PCR
CMV viral load was detected by extraction of DNA from urine samples (QIAmp Viral DNA Kit, Qiagen) and analyzed by real time PCR (Realquality RQ-CMV AB Analitica) according to the manufacturer’s instructions (AB Analitica). The assay detected CMV DNA in a linear range from 600 copies/mL to 6 × 108 copies/mL.
Analysis of polyfunctionality of T-cell responses
Ex vivo expression of IFN-γ, IL-2 and TNF-α by CD4+ and CD8+ T-cells was assessed by multicolor flow cytometry analysis. In brief, freshly isolated PBMCs (2 × 106 cells per condition) were stimulated in polypropylene tubes with PMA (6.25ng/mL) plus ionomicyn (0.6 μM) and with a recombinant HIV p24 capsid protein (5.5 µg/ml, Protein Sciences Corp) to evaluate global T-cell functionality and HIV-specific response, respectively. In addition, to characterize more accurately the CD4+ T-cell functionality, the most impaired population in immunodiscordant individuals, different CMV antigenic preparations optimal for stimulating a robust response especially from CD4+ T-cells were included57,58,59: a purified CMV viral lysate (0.5 µg/ml, ZeptoMetrix, Buffalo, NY), a pool of overlapping peptides (15-mer) covering the whole HCMV pp65 protein (30 µg/ml, obtained through the NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH59,60,61 and a recombinant CMV-pp65 protein (7.5 μL per sample, Milteny Biotec, Madrid, Spain). An unstimulated sample control was also included. Co-stimulatory anti-CD49d and anti-CD28 (1 µg/ml, Becton Dickinson) were added to all tubes and after 2 hours of incubation at 37 °C, Brefeldin A and Monensin were added according to the instructions of the manufacturer (both from Becton Dickinson) and cells were incubated for an additional 6 h. Cells were then washed and labeled with the Fixable Yellow Dead Cell Stain Kit (Invitrogen) to identify dead cells, washed and stained with the following extracellular antibody combination: V450-CD3, V500-CD8, Alexa700-CD45RA, PE-Cy7-CCR7 and BV605-CD27 (all from Becton Dickinson) and eFluor650-CD4 (e-Biosciences). Subsequently, cells were fixed and permeabilized with Cytofix/Cytoperm (Becton Dickinson) and intracellular staining was carried out with FITC-IFN-γ, APC-IL-2 and PE-TNF-α (all from Becton Dickinson). After intracellular staining, cells were washed and acquired in LSR-Fortessa flow cytometer (Becton Dickinson) in the Cytometry Core Facility at Germans Trias i Pujol Research Institute.
Analysis of flow cytometry data
Flow cytometry data were analysed using FlowJo software (9.8 v; TreeStar, Portland, OR, USA). Initial gating was performed on the forward scatter height vs. forward scatter area to remove doublets, and then living T lymphocytes were gated according morphological parameters and cell viability followed by identification of CD4+ and CD8+ T-cells (Figure S1). Mitogen PMA/ionomycin stimulation leads to a rapid downregulation of membrane expression of CD4+ 62,63, therefore CD3+ CD8- T-cells were used to analyzed the CD4+ T-cell response in the non-specific analysis. Finally, CD4+ and CD8+ T-cell subpopulations were identified by the expression of CD45RA, CCR7 and CD27 (Figure S1). For the cytokine production analysis, individual gates (set on the basis of the unstimulated control) were made to identify positive responses (Figure S1). FlowJo Boolean gating was performed to create a full array of possible combinations of up to 8 response patterns from the CD4+ and CD8+ T-cell gate. After subtracting the background response (with co-stimulation but no antigen), a positive CD4+ and CD8+ T-cell cytokine response was defined as at least 40 positive events (to minimize the possibility of error due to a low number of events when further subdividing these cells based on their phenotypic profile). Polyfunctionality was represented visually using Pestle (v1.7) and SPICE (v5.35) software (provided by the National Institutes of Health, Mario Roederer, ImmunoTechnology Section, Vaccine Research Centre, National Institutes of Health, Bethesda, MD, USA)64. In addition to SPICE analysis, we used the polyfunctional index (PI) defined by Larsen et al.65 (Funky Cells Toolbox 66), which numerically evaluates the degree and variation of polyfunctionality and allows comparative statistical tests.
Differentiation profile of IFN+ and IL-2+ CD4+ and CD8+ T-cells were created by FlowJo boolean gating and subsequent SPICE analysis. In addition, functionality of each of the CD45/CCR7/CD27 differentiation subsets was assessed by the determination of IFN-γ+ and IL-2+ cells within those subsets. In the CMV- and HIV-specific response we do not report on TNF-α production to avoid false-positive results because a low signal-to-background ratio was found in the staining.
Statistical analysis
Statistical analysis and graphical presentation were performed using Prism software (5.0av; GraphPad). To compare pie charts, we used the SPICE permutation (10,000 permutations) analysis. Mann-Whitney U nonparametric test was used for comparing data among specific responses (*p < 0.05, **p < 0.01, ***p < 0.001). Spearman’s correlation coefficient was calculated to identify associations between variables.
References
Palella, F. J. et al. Declining morbidity and mortality among patients with advanced human immunodeficiency virus infection. HIV Outpatient Study Investigators. N. Engl. J. Med. 338, 853–860 (1998).
Piketty, C. et al. Discrepant responses to triple combination antiretroviral therapy in advanced HIV disease. AIDS 12, 745–750 (1998).
Kelley, C. F. et al. Incomplete peripheral CD4+ cell count restoration in HIV-infected patients receiving long-term antiretroviral treatment. Clin Infect Dis 48, 787–794 (2009).
Massanella, M., Negredo, E., Clotet, B. & Blanco, J. Immunodiscordant responses to HAART–mechanisms and consequences. Expert Rev Clin Immunol 9, 1135–1149 (2013).
Massanella, M. et al. CD4 T-cell hyperactivation and susceptibility to cell death determine poor CD4 T-cell recovery during suppressive HAART. AIDS 24, 959–968 (2010).
Molina-Pinelo, S. et al. Premature immunosenescence in HIV-infected patients on highly active antiretroviral therapy with low-level CD4 T cell repopulation. J. Antimicrob. Chemother. 64, 579–588 (2009).
Negredo, E. et al. Nadir CD4 T Cell Count as Predictor and High CD4 T Cell Intrinsic Apoptosis as Final Mechanism of Poor CD4 T Cell Recovery in Virologically Suppressed HIV‐Infected Patients: Clinical Implications. Clin Infect Dis 50, 1300–1308 (2010).
Viard, J. P. et al. Influence of age on CD4 cell recovery in human immunodeficiency virus-infected patients receiving highly active antiretroviral therapy: evidence from the EuroSIDA study. J. Infect. Dis. 183, 1290–1294 (2001).
Marchetti, G. et al. Comparative analysis of T-cell turnover and homeostatic parameters in HIV-infected patients with discordant immune-virological responses to HAART. AIDS 20, 1727–1736 (2006).
Lederman, M. M. et al. Immunologic Failure Despite Suppressive Antiretroviral Therapy Is Related to Activation and Turnover of Memory CD4 Cells. J. Infect. Dis 204, 1217–1226 (2011).
Marchetti, G. et al. Skewed T-cell maturation and function in HIV-infected patients failing CD4+ recovery upon long-term virologically suppressive HAART. AIDS 24, 1455–1460 (2010).
Erikstrup, C. et al. T-cell dysfunction in HIV-1-infected patients with impaired recovery of CD4 cells despite suppression of viral replication. J. Acquir. Immune Defic. Syndr. 53, 303–310 (2010).
Massanella, M. et al. Increased ex vivo cell death of central memory CD4 T cells in treated HIV infected individuals with unsatisfactory immune recovery. J Transl Med 13, 230 (2015).
Méndez-Lagares, G. et al. Differential alterations of the CD4 and CD8 T cell subsets in HIV-infected patients on highly active antiretroviral therapy with low CD4 T cell restoration. J. Antimicrob. Chemother. 67, 1228–1237 (2012).
Nakanjako, D. et al. High T-cell immune activation and immune exhaustion among individuals with suboptimal CD4 recovery after 4 years of antiretroviral therapy in an African cohort. BMC Infect. Dis. 11, 43 (2011).
Robain, M., Carré, N., Dussaix, E., Salmon-Ceron, D. & Meyer, L. Incidence and sexual risk factors of cytomegalovirus seroconversion in HIV-infected subjects. The SEROCO Study Group. Sex Transm Dis 25, 476–480 (1998).
Webster, A. et al. Cytomegalovirus infection and progression towards AIDS in haemophiliacs with human immunodeficiency virus infection. Lancet 2, 63–66 (1989).
Weltevrede, M., Eilers, R., de Melker, H. E. & van Baarle, D. Cytomegalovirus persistence and T-cell immunosenescence in people aged fifty and older: A systematic review. Exp. Gerontol. 77, 87–95 (2016).
Huygens, A. et al. Functional Exhaustion Limits CD4+ and CD8+ T-Cell Responses to Congenital Cytomegalovirus Infection. J. Infect. Dis. 212, 484–494 (2015).
Hunt, P. W. et al. Valganciclovir reduces T cell activation in HIV-infected individuals with incomplete CD4+ T cell recovery on antiretroviral therapy. J. Infect. Dis 203, 1474–1483 (2011).
Gianella, S., Massanella, M., Wertheim, J. O. & Smith, D. M. The Sordid Affair Between Human Herpesvirus and HIV. J. Infect. Dis. 212, 845–852 (2015).
Gianella, S. & Letendre, S. Cytomegalovirus and HIV: A Dangerous Pas de Deux. J. Infect. Dis. 214(Suppl 2), S67–74 (2016).
Naeger, D. M. et al. Cytomegalovirus-specific T cells persist at very high levels during long-term antiretroviral treatment of HIV disease. PLoS ONE 5 (2010).
Appay, V. et al. Old age and anti-cytomegalovirus immunity are associated with altered T-cell reconstitution in HIV-1-infected patients. AIDS 25, 1813–1822 (2011).
Hsue, P. Y. et al. Increased carotid intima-media thickness in HIV patients is associated with increased cytomegalovirus-specific T-cell responses. AIDS 20, 2275–2283 (2006).
Parrinello, C. M. et al. Cytomegalovirus immunoglobulin G antibody is associated with subclinical carotid artery disease among HIV-infected women. J. Infect. Dis 205, 1788–1796 (2012).
Patel, E. U. et al. Elevated cytomegalovirus IgG antibody levels are associated with HIV-1 disease progression and immune activation. AIDS 31, 807–813 (2017).
Chidrawar, S. et al. Cytomegalovirus-seropositivity has a profound influence on the magnitude of major lymphoid subsets within healthy individuals. Clin. Exp. Immunol 155, 423–432 (2009).
Derhovanessian, E. et al. Infection with cytomegalovirus but not herpes simplex virus induces the accumulation of late-differentiated CD4+ and CD8+ T-cells in humans. J. Gen. Virol. 92, 2746–2756 (2011).
Almanzar, G. et al. Long-term cytomegalovirus infection leads to significant changes in the composition of the CD8+ T-cell repertoire, which may be the basis for an imbalance in the cytokine production profile in elderly persons. Journal of Virology 79, 3675–3683 (2005).
Moro-García, M. A. et al. Relationship between functional ability in older people, immune system status, and intensity of response to CMV. Age (Dordr) 34, 479–495 (2012).
Bajwa, M. et al. Functional Diversity of Cytomegalovirus-Specific T Cells Is Maintained in Older People and Significantly Associated With Protein Specificity and Response Size. J. Infect. Dis. 214, 1430–1437 (2016).
Lelic, A. et al. The polyfunctionality of human memory CD8+ T cells elicited by acute and chronic virus infections is not influenced by age. PLoS Pathog. 8, e1003076 (2012).
Minami, R., Takahama, S., Kaku, Y. & Yamamoto, M. Addition of maraviroc to antiretroviral therapy decreased interferon-γ mRNA in the CD4+ T cells of patients with suboptimal CD4+ T-cell recovery. J. Infect. Chemother 23, 29–34 (2017).
Casetti, R. et al. HIV-Specific CD8 T Cells Producing CCL-4 Are Associated With Worse Immune Reconstitution During Chronic Infection. J. Acquir. Immune Defic. Syndr. 75, 338–344 (2017).
Sallusto, F., Lenig, D., Förster, R., Lipp, M. & Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 401, 708–712 (1999).
Roff, S. R., Noon-Song, E. N. & Yamamoto, J. K. The Significance of Interferon-γ in HIV-1 Pathogenesis, Therapy, and Prophylaxis. Front Immunol 4, 498 (2014).
Williams, M. A., Tyznik, A. J. & Bevan, M. J. Interleukin-2 signals during priming are required for secondary expansion of CD8+ memory T cells. Nature 441, 890–893 (2006).
Sachdeva, N. et al. Poor lymphoproliferative responses with low proportion of Gag-specific CD8 TEMRA cells in HIV-1-infected patients showing immunological and virological discordance despite prolonged suppression of plasma viremia. Viral Immunol. 23, 49–61 (2010).
Stone, S. F., Price, P. & French, M. A. Cytomegalovirus (CMV)-specific CD8+ T cells in individuals with HIV infection: correlation with protection from CMV disease. J. Antimicrob. Chemother. 57, 585–588 (2006).
Gianella, S. et al. Shedding of HIV and human herpesviruses in the semen of effectively treated HIV-1-infected men who have sex with men. CLIN INFECT DIS 57, 441–447 (2013).
Lachmann, R. et al. Polyfunctional T cells accumulate in large human cytomegalovirus-specific T cell responses. Journal of Virology 86, 1001–1009 (2012).
Erlandson, K. M. et al. Physical function impairment of older, HIV-infected adults is associated with cytomegalovirus immunoglobulin response. AIDS Research and Human Retroviruses 31, 905–912 (2015).
Gianella, S. et al. Cytomegalovirus replication in semen is associated with higher levels of proviral HIV DNA and CD4+ T cell activation during antiretroviral treatment. Journal of Virology 88, 7818–7827 (2014).
Pitcher, C. J. et al. HIV-1-specific CD4+ T cells are detectable in most individuals with active HIV-1 infection, but decline with prolonged viral suppression. Nat. Med. 5, 518–525 (1999).
Champagne, P. et al. Skewed maturation of memory HIV-specific CD8 T lymphocytes. Nature 410, 106–111 (2001).
Appay, V. et al. Memory CD8+ T cells vary in differentiation phenotype in different persistent virus infections. Nat. Med 8, 379–385 (2002).
Harari, A., Vallelian, F., Meylan, P. R. & Pantaleo, G. Functional heterogeneity of memory CD4 T cell responses in different conditions of antigen exposure and persistence. J. Immunol. 174, 1037–1045 (2005).
Méndez-Lagares, G. et al. Severe immune dysregulation affects CD4+CD25(hi)FoxP3+ regulatory T cells in HIV-infected patients with low-level CD4 T-cell repopulation despite suppressive highly active antiretroviral therapy. Journal of Infectious Diseases 205, 1501–1509 (2012).
Hunt, P. W. et al. HIV-specific CD4+ T cells may contribute to viral persistence in HIV controllers. CLIN INFECT DIS 52, 681–687 (2011).
Cockerham, L. R. et al. CD4+ and CD8+ T cell activation are associated with HIV DNA in resting CD4+ T cells. PLoS ONE 9 (2014).
Chomont, N. et al. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat. Med 15, 893–900 (2009).
Douek, D. C. et al. HIV preferentially infects HIV-specific CD4+ T cells. Nature 417, 95–98 (2002).
Riou, C. et al. Differential impact of magnitude, polyfunctional capacity, and specificity of HIV-specific CD8+ T cell responses on HIV set point. Journal of Virology 88, 1819–1824 (2014).
Almeida, J. R. et al. Superior control of HIV-1 replication by CD8+ T cells is reflected by their avidity, polyfunctionality, and clonal turnover. J. Exp. Med. 204, 2473–2485 (2007).
Duvall, M. G. et al. Polyfunctional T cell responses are a hallmark of HIV-2 infection. Eur. J. Immunol. 38, 350–363 (2008).
Sylwester, A. W. et al. Broadly targeted human cytomegalovirus-specific CD4+ and CD8+ T cells dominate the memory compartments of exposed subjects. J. Exp. Med. 202, 673–685 (2005).
Weinberg, A. et al. In vitro cell-mediated immune responses of human immunodeficiency virus-infected and -uninfected individuals to whole cytomegalovirus antigens and their subunits. Clin. Vaccine Immunol. 15, 1398–1409 (2008).
Kern, F. et al. Cytomegalovirus (CMV) phosphoprotein 65 makes a large contribution to shaping the T cell repertoire in CMV-exposed individuals. J. Infect. Dis. 185, 1709–1716 (2002).
Kern, F. et al. Analysis of CD8 T cell reactivity to cytomegalovirus using protein-spanning pools of overlapping pentadecapeptides. Eur. J. Immunol. 30, 1676–1682 (2000).
Maecker, H. T. et al. Use of overlapping peptide mixtures as antigens for cytokine flow cytometry. Journal of Immunological Methods 255, 27–40 (2001).
Baran, J., Kowalczyk, D., Ozóg, M. & Zembala, M. Three-color flow cytometry detection of intracellular cytokines in peripheral blood mononuclear cells: comparative analysis of phorbol myristate acetate-ionomycin and phytohemagglutinin stimulation. Clin. Diagn. Lab. Immunol. 8, 303–313 (2001).
Petersen, C. M., Christensen, E. I., Andresen, B. S. & Møller, B. K. Internalization, lysosomal degradation and new synthesis of surface membrane CD4 in phorbol ester-activated T-lymphocytes and U-937 cells. Exp. Cell Res. 201, 160–173 (1992).
Roederer, M., Nozzi, J. L. & Nason, M. C. SPICE: Exploration and analysis of post-cytometric complex multivariate datasets. Cytometry 79A, 167–174 (2011).
Larsen, M. et al. Evaluating cellular polyfunctionality with a novel polyfunctionality index. PLoS ONE 7 (2012).
Larsen, M. http://www.funkycells.com/main (2016).
Acknowledgements
We are grateful to Anuska Llano for her help with the evaluation of specific response to technical staff of IrsiCaixa for sample processing and the clinical staff of Fundació Lluita contra la SIDA. We thank the IGTP Cytometry Core Facility and Marco Antonio Fernández for his contribution to this publication. The authors would like to thank all the individuals who were participants in the study. IrsiCaixa and IGTP are part of the CERCA Program/Generalitat de Catalunya. This work was supported by the EC11-045 project (Ministerio de Sanidad y Política Social) and the Spanish AIDS network ‘Red Temática Cooperativa de Investigación en SIDA’ (RD12/0017/0002). Grant RD12/0017/0002 is co-funded by the Spanish Instituto de Salud Carlos III (ISCIII) and the European Fund for Regional Development (FEDER). C.C. and J.B. are supported by the ISCIII and the Health Department of the Catalan Government (Generalitat de Catalunya).
Author information
Authors and Affiliations
Contributions
E.G.M. and E.G. performed the functional analysis. E.N. and B.C. recruited and provided clinical care for the participants, J.P. obtained the blood samples. V.U. performed the statistical analysis. E.N., B.C., J.B., M.M. and C.C. designed the study and developed the hypotheses. E.G.M. and C.C. analyzed and interpreted the data. E.G.M. and C.C. drafted the article. All authors contributed to the article’s preparation and approved its final version.
Corresponding author
Ethics declarations
Competing Interests
E.G.M., M.M., E.G., V.U., J.P., J.B., E.N. and C.C. report no disclosures. B.C. have served as a consultant to and/or have received research grant support from Gilead, Janssen, MSD, ViiV, BMS.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gómez-Mora, E., García, E., Urrea, V. et al. Preserved immune functionality and high CMV-specific T-cell responses in HIV-infected individuals with poor CD4+ T-cell immune recovery. Sci Rep 7, 11711 (2017). https://doi.org/10.1038/s41598-017-12013-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-12013-2
This article is cited by
-
The forces driving clonal expansion of the HIV-1 latent reservoir
Virology Journal (2020)
-
Cytomegalovirus-specific T-cells are associated with immune senescence, but not with systemic inflammation, in people living with HIV
Scientific Reports (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
