
Abstract
Extracellular adenosine, generated via the concerted action of CD39 and CD73, contributes to T-cell differentiation and function. Adenosine concentrations are furthermore influenced by adenosine deaminase binding protein CD26. Because aberrant T-cell phenotypes had been reported in anti-neutrophil cytoplasmic auto-antibody (ANCA)-associated vasculitis (AAV) patients, an impaired expression of these molecules on T-cells of AAV patients was hypothesized in the present study. While in AAV patients (n = 29) CD26 was increased on CD4+ lymphocytes, CD39 and CD73 were generally reduced on patients’ T-cells. In CD4+ cells significant differences in CD73 expression were confined to memory CD45RA- cells, while in CD4- lymphocytes differences were significant in both naïve CD45RA+ and memory CD45RA- cells. The percentage of CD4-CD73+ cells correlated with micro-RNA (miR)−31 expression, a putative regulator of factor inhibiting hypoxia-inducible factor 1 alpha (FIH-1), inversely with serum C-reactive protein (CRP) and positively with estimated glomerular filtration rate (eGFR). No correlation with disease activity, duration, and ANCA profile was found. It remains to be assessed if a decreased CD73 and CD39 expression underlies functional impairment of lymphocytes in AAV patients. Likewise, the relations between frequencies of CD4-CD73+ cells and serum CRP or eGFR require further functional elucidation.
Similar content being viewed by others

Introduction
There is compelling evidence that immune cell function and immune responses are heavily influenced by purinergic mediators. Following a variety of tissue insults, ATP is released into the extracellular space where it is rapidly converted to ADP, AMP and adenosine by the membrane associated ecto-nucleotidases CD39 and CD731,2. Deamination of adenosine finally results in the generation of inosine and ammonia. Adenosine deaminase (ADA) deficiency is associated with severe combined immunodeficiency (SCID) underscoring its role in immune regulation3. Human ADA1 can bind dipeptidyl peptidase IV (CD26) on T-cells and may counteract regulatory T-cell-mediated T-cell suppression4. The local expression of CD73, CD39 and CD26 thus controls concentrations of ATP, ADP, AMP, adenosine and inosine in the extracellular space5,6,7,8.
While ATP promotes inflammation, T-cell activation and Th17 differentiation, adenosine controls vascular permeability and exerts immunosuppressive effects9 mainly through activation of A2A receptors on immune cells10,11. Hypoxia strongly influences adenosine signalling to immune cells partly via stabilisation of hypoxia inducible factor-1α (HIF-1α), which in turn drives the induction of adenosine receptors and CD7312. Apart from hypoxia, CD73 is regulated by various cytokines, e.g. interferon(IFN)-β13, TGFβ, IFNγ, IL-6, and IL-1214, and by adenosine itself in a paracrine manner15. CD73 expression is influenced indirectly by factors that regulate factor inhibiting HIF-1α (FIH-1), e.g. micro-RNAs (miR)16,17,18,19. The importance of CD73 in cell mediated immunity has been appreciated amongst others in studies using CD73 deficient mice20 and models of auto-immune uveitis21. Other experimental studies have suggested CD73 as novel therapeutic target in cancer22,23, multiple sclerosis24, and chronic Toxoplasma gondii infection25.
Anti-neutrophil cytoplasmic auto-antibody (ANCA)-associated vasculitis (AAV), i.e. granulomatosis with polyangiitis (GPA), microscopic polyangiitis (MPA), eosinophilic granulomatosis with polyangiitis (EGPA) and renal-limited AAV26 share common histopathological findings of systemic small vessel vasculitis with fibrinoid necrosis of the vessel wall or necrotizing, crescentic glomerulonephritis27,28. While ANCA are believed to be directly involved in the pathogenesis of AAV, their presence suggests impaired regulation of immune tolerance. Because isotype switching from IgG1 to IgG4, the predominant IgG ANCA subclasses29, depends on T-cell cytokines, impaired regulation of immune tolerance in AAV patients may not be restricted to the B-cell compartment alone but likely involves T-cells as well. Indeed, aberrant T-cell phenotypes have been identified in AAV patients including expanded effector memory T-cells30, defective regulatory T-cells31 and expanded Th17 cells32. In keeping with these findings and the role of adenosine in immunity, we hypothesized that T-lymphocytes from AAV patients may disclose an altered expression of cell surface molecules involved in adenosine generation and turn-over. In addition, we assessed if this was associated with differential miR expression and to what extent an altered CD39 and CD73 expression correlated with clinical entities.
Results
CD73, CD39 and CD26 expression on T-cells of AAV patients
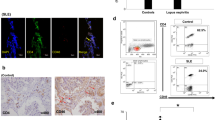
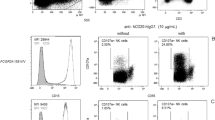
CD73, CD39 and CD26 expression were assessed by FACS on peripheral blood lymphocytes of AAV patients (n = 29, 2 samples were not eligible for FACS analysis) and healthy controls (HC, n = 12). Demographic and clinical characteristics are depicted in Table 1 and in methods. Both the frequency and median fluorescence intensity (MFI) of CD73+ lymphocytes were significantly reduced in AAV patients as compared to HC (Fig. 1a). This was found in both CD4- and CD4+ lymphocytes, albeit these differences were more pronounced in the former population (Fig. 1b). Differences in the frequency of CD73+ cells between patients and HC were significant within the CD4+ population only in the memory CD45RA- lymphocytes (Fig. 1c), while in the CD4- lymphocytes this was observed for both naïve CD45RA+ and memory CD45RA- lymphocytes (Fig. 1d). Figure 2 shows illustrative dot plots from an exemplary AAV patient and HC for CD73 expression in all lymphocyte subsets. Expression differences for CD39 between patients and HC were confined to the CD4- lymphocytes. CD4-CD45RA+ lymphocytes of patients showed a reduced frequency of CD39, yet CD39 MFI was increased on these cells. In contrast, CD39 MFI in the CD4-CD45RA- lymphocytes of patients was reduced with no significant change in the percentage of CD39+ cells between patients and HC (Table 2). Differences in CD26 expression were only observed in CD4+ lymphocytes, in both the naïve and memory subsets (Table 2). When MFI for CD73, CD39 and CD26 was ranked from high to low, it was found that in patients MFI of CD26 was higher than that of CD73 (CD26 > CD73 > CD39; 1498 ± 506.8, 1082 ± 361.5 and 615.4 ± 172.1, respectively), while in HC CD73 was expressed stronger compared to CD26 (CD73 > CD26 > CD39; 1459 ± 415.1, 1173 ± 206.5 and 679.6 ± 190.8, respectively).

Reduction of CD73 frequency in lymphocytes from AAV patients. (a) CD73 frequency and protein level (median fluorescence intensity, MFI, given in relative light units, RLU) were significantly reduced in the entire lymphocytic population from ANCA patients. (b) CD73+ cells were reduced both in the CD4+ and CD4- fraction in the patient cohort. (c) CD4+CD73+ populations were solely decreased in the memory (CD45RA-) subset from ANCA patients, while CD4- CD73+ cells were significantly diminished in the memory and the naïve (CD45RA+) subpopulation as well (d). AAV: patient group, HC: control group. Non-parametric two-tailed Mann-Whitney-U-test was used to test for significance; ns: not significant.

Exemplary CD73+ lymphocyte populations of one AAV patient (AAV, left column) compared to a healthy control (HC, right column). First row: all lymphocytes were gated, CD4 intensity is plotted against CD73. Only CD4+ lymphocytes (second row), and only CD4- lymphocytes (third row) were gated, CD45RA intensity is plotted against CD73. Colours indicate cell density in descending order: red, yellow, green, blue.
CD4-CD73+ cells correlate with inflammation and renal function of AAV patients
We next explored if changes in T-cell subsets correlate with clinical entities, e.g. relapse rate, disease duration (time since initial diagnosis) and activity, ANCA profile, serum CRP and renal function assessed by estimated glomerular filtration rate (eGFR). Because for CD73, CD39 and CD26 expression on T-cells the difference between patients and HC were the strongest for the CD4-CD73+ T-cell subset, we chose the frequency of CD4-CD73+ T-cells as independent variable in univariate analysis. While no significant variation was evident between patients with active disease and in remission or different ANCA profiles as well as disease duration and relapse rate, the frequency of CD4-CD73+ T-cells disclosed an inverse correlation with serum CRP (r = −0.508; P = 0.0049, Fig. 3a) and was positively correlated with eGFR (r = 0.492; P = 0.0067, Fig. 3b).

Correlation analysis of CD4-CD73+ lymphocytes with renal function and inflammation. (a) Frequency of CD4-CD73+ cells positively correlated with renal function assessed by eGFR; Spearman’s r = 0.492; P = 0.0067; R2 = 0.2137. (b) Frequency of CD4-CD73+ cells negatively correlated with CRP concentration in peripheral blood; Spearman’s r = −0.5081; P = 0.0049; R2 = 0.1657. Black lines indicate regression line.
miR analysis by TaqMan Low Density Arrays
Since expression of miRs may influence the expression of CD73 directly or indirectly through HIF-1α or its repressor factor inhibiting HIF-1α (FIH1)33,34 we analysed miR expression in peripheral blood mononuclear cells (PBMC) of 20 patients (GPA: n = 16, MPA: n = 4) and 12 controls by means of TaqMan Low Density Array. We observed 29 miRs to be expressed significantly different when comparing AAV patients and HC (Table 3), while the vast majority of these miRs were up-regulated (fold change ranging from 1.37–2.69 in patients), only one miR, i.e. miR-31, was down-regulated (fold change 0.31 in patients) and found to be the most significant amongst the 29 differently expressed miRs.
miR-31 correlates with lymphocytic CD73 frequency
As miR-31 was previously reported to be a direct target for FIH-1 mRNA33 and because HIF-1α is a transcription factor for CD7312, we performed qPCR for miR-31, HIF-1α and FIH-1 mRNA in our complete study cohort (AAV: n = 30, HC: n = 12). Indeed, we confirmed that miR-31 was significantly lower expressed in AAV patients (mean fold change ± s.d.: 0.55 ± 0.48), but unexpectedly, also its putative target FIH-1 mRNA was significantly down-regulated in the patient group (mean fold change ± s.d.: 0.87 ± 0.24) (Fig. 4a), whereas HIF-1α mRNA did not show significant discrepancy in the patient group (mean fold change ± s.d.: 0.81 ± 0.77, not significant). Nonetheless, the expression of miR-31 significantly correlated with the frequency of CD73+ T-cells when all study participants were analysed (Fig. 4b).

Significant down-regulation of miR-31 and its target FIH-1 mRNA in AAV patients. (a) miR-31 was repeatedly down-regulated in the patient group (AAV: n = 30, healthy controls: n = 12), so was its direct target FIH-1 mRNA as indicated by comparison of delta cycle threshold (ΔCT) values from qPCR between AAV patients and healthy controls. Non-parametric two-tailed Mann-Whitney-U-test was used to test for significance. (b) Quantity of miR-31 positively correlated with frequency of CD73+ lymphocytes in the entire study cohort (n = 40), Spearman’s r = 0.5433; P = 0.0003; R2 = 0.2838. Black lines indicate regression line.
Discussion
Our study demonstrates that the expression of CD73 and CD39, that are both implicated in the generation of extracellular adenosine, are down-regulated on T-cells of AAV patients, while the expression of CD26 on these cells is up-regulated. Since CD26 strongly associates with ADA, our findings may suggest an imbalance in T-cell mediated adenosine generation and turn-over in AAV patients. This may not be specific for AAV patients as serum ADA activity has also been reported to be increased in SLE patients35, while the expression of the adenosine-generating enzymes (CD39, CD73) seems to be decreased36,37. Most of the published studies on CD39 and CD73 expression on lymphocytes in relation to auto-immune diseases have focused on regulatory T-cells. In the current study, we demonstrate that CD39 and CD73 are expressed on all major T-cell subsets to a different extent and that differences between patients and HC in expression levels of these molecules unlikely are restricted to regulatory T-cells.
We are aware that CD73 expression on lymphocytes decreases with age38. Although this was also found in our study cohort, down-regulation of CD73 in AAV patients was still noticed when comparing individuals above 40 years of age in both groups.
We did not find correlations between CD73 expression and disease activity, relapse rate or time since initial diagnosis, yet, the relative number of CD4-CD73+ T-cells showed a correlation with serum CRP and renal function. An inverse correlation between CD73 and CRP has also been described in HIV patients. In these patients, depletion of CD4+CD73+ T-cells coincides with increased activation markers on CD4+ and CD8+ cells39 suggesting a role in persistent immune activation and inflammation. Nonetheless, we cannot exclude that CD39 and CD73 expression are influenced by immunosuppressive medication as has been demonstrated for methotrexate in the treatment of rheumatoid arthritis patients40,41 or azathioprine in inflammatory bowel disease patients42,43. Also rituximab and cyclophosphamide have been shown to modify peripheral T-cell subset frequencies44,45, but their effect on the expression of ecto-nucleotidases still remains elusive. Only four patients in complete remission, three presenting with normal and one with slightly reduced renal function, were treated with methotrexate and only two patients received mycophenolate mofetil. Even though AAV patients are relatively lymphopenic46, show persistent immune activation47 and may have signs of subclinical inflammation prior to relapses48, the cause of a diminished CD73+ T-cell population and its consequences in relation to inflammation and persistent immune activation remains to be assessed.
A relation between CD73 and reno-protection has been described in ischemia reperfusion injury (IRI) models in which adoptively transferred CD73+, but not CD73-deficient regulatory T-cells protect wild-type mice from kidney IRI49. Also, the glomerular filtration rate depends on CD73 derived adenosine as a main component of the tubulo-glomerular feedback50. In addition, CD73 deficiency appears to be associated with a vascular pro-inflammatory phenotype51, comprising the renal glomerular endothelium and interstitium20. Whether a decreased expression of CD73 in AAV patients is only confined to T-cells or also occurs in renal tissue is to our knowledge not known and subject of our currently ongoing research.
Inasmuch as miR expression, as part of epigenetic reprogramming, participates in T-cell activation and differentiation, dysregulation hereof may yield aberrant T-cell phenotypes contributing to auto-immunity52. In AAV patients, miR expression has been studied in the circulation53,54 and in peripheral blood mononuclear cells55. Amongst the significantly changed miR in our study, 3 miR are indirectly associated with CD73 expression, i.e. miR-31, miR-424 and miR-34a. While the latter 2 miR increase CD73 expression through HIF-1α up-regulation56 or inhibition of HIF-1α proteasomal degradation57, miR-31 might influence CD73 expression via targeting FIH-1 mRNA33. Since FIH-1 impedes the transcriptional activity of HIF-1α and thus transcription of CD7334, down-regulation of miR-31 may result in down-regulation of CD73. Indeed, the strong correlation with CD73 expression found in our study suggests a regulatory role for miR-31, yet this assumption awaits further experimental confirmation. It is noteworthy to mention that in addition to the findings of Li et al. on CD39 and CD73 expression on lymphocytes of SLE patients36,37, also in these patients, down-regulation of miR-31 has been reported.
Lymphocyte infiltrations in vascular lesions of AAV patients have been described in numerous studies58,59. Aberrant T-cell phenotypes, e.g. memory CD8+CD28- cells which are believed to produce IFNγ and TNFα60, can be found in these infiltrates. It is therefore tempting to speculate that a relative shortage of adenosine, potentially caused by changes in CD26, CD39 and CD73 expression, may drive the pro-inflammatory milieu in lesions of AAV patients. The expression of CD26 in nasal biopsies of patients with localized GPA has been reported to be high61, yet the expression of CD39 and CD73 has not been studied in lesions of AAV patients.
In conclusion, the findings of our study suggest that T-cells from AAV patients may disclose an impaired ability to generate adenosine as a consequence of reduced expression of adenosine generating ecto-nucleotidases (CD39 and CD73) while at the same time an increased expression of the adenosine deaminase binding protein CD26 may favour adenosine turn-over. Our study, however, does not provide experimental evidence that adenosine concentrations are low in lesions of AAV patients and that this in turn underlies the pro-inflammatory milieu of such lesions. Nonetheless, the correlations found with CRP and renal function indicate a potential role for adenosine, but warrants further functional studies to underpin the clinical relevance of our findings.
Methods
Recruitment of study participants
Biopsy proven AAV patients (n = 31) treated in our department and healthy controls (n = 12, male: n = 6, female: n = 6, mean age ± standard deviation: 55.5 ± 16.1 years) were investigated in this study. For detailed patient cohort characteristics see Table 1. Diagnosis of AAV followed the Chapel Hill Nomenclature and diagnostic criteria of the American College of Rheumatology. Disease activity was assessed using the Birmingham Vasculitis Activity Score (BVAS) for EUVAS studies modified from Luqmani et al. 1994 (QJM: monthly journal of the Association of Physicians) with active disease defined as BVAS > 0. Relapse was defined as recurrence of active disease requiring escalation of immunosuppressive treatment from maintenance therapy. Disease duration was calculated as the time from initial diagnosis to study enrolment, while time in remission was defined as the period between latest relapse and study inclusion. Relapse rate was calculated as number of relapses per month since first AAV manifestation.
Sample acquisition
Peripheral venous blood was drawn once from patients and healthy controls using 2,2′,2″,2″′-(Ethane-1,2-diyldinitrilo)-tetraacetic acid (EDTA) containing tubes as anticoagulant. Samples were processed immediately at room temperature and protected from daylight after acquisition. Measurement of C-reactive protein (CRP) and MDRD-eGFR in the patient group were part of clinical routine. All patients and healthy controls gave written informed consent. The local ethics committee (Medizinische Ethikkommission II der Universität Heidelberg am Universitätsklinikum Mannheim) approved the entire study (reference number 2011–339 N). All experiments were performed in accordance with relevant guidelines and regulations.
FACS analysis
Fourfold whole blood staining with directly conjugated antibodies for CD4 (Pacific Blue, clone RPA-T4), CD45RA (FITC, clone HI100), CD25 (APC, clone M-A251), CD39 (RPE, clone TU66), CD73 (RPE, clone AD2), and CD26 (RPE, clone M-A261) (all purchased from BD Biosciences, Heidelberg, Germany), was performed on 100 μl of each sample (due to technical reasons, 2 patients could not be analysed by FACS; AAV: n = 29, HC: n = 12). Optimal antibody concentrations were determined by serial dilution. After antibody incubation, red blood cells were lysed using BD FACS lysing solution (BD Biosciences, Heidelberg, Germany) followed by thorough washing with phosphate buffered saline (PBS, Sigma Aldrich) containing 2 mM EDTA (Sigma Aldrich) and 3% (w/v) bovine serum albumine (Serva Electrophoresis GmbH, Heidelberg, Germany) and finally fixation with BD cellFIX solution (BD Biosciences, Heidelberg, Germany). For compensation BD compBeads (BD Biosciences, Heidelberg, Germany) were stained in a similar manner as described above. Flow cytometry was performed on a BD FACS Canto II cytometer using negative and fluorescence-minus-one controls and data analysis was performed using FlowJo software version 5.2 (FlowJo LLC, Ashland, USA).
RNA isolation from PBMC
PBMC were isolated from whole blood using PolymorphPrepTM (Progen Biotechnik GmbH, Heidelberg, Germany) density centrifugation at 450 g for 40 min as previously described62. PBMC were collected on the interface, transferred to a new tube and thoroughly washed. Cell numbers were assessed on a Casy1 TT instrument (Schärfe System, Reutlingen, Germany). RNA was isolated using TRIzol (Invitrogen, Carlsbad, USA) according to the manufacturer’s instructions. After precipitation, the RNA pellet was dissolved in nuclease free water and quality and concentrations were assessed on an Infinite 200 plate reader (Tecan Trading AG, Männedorf, Switzerland). Only RNA isolates with an absorbance ratio (wavelength 260 nm/280 nm) ≥ 2.0 were used in the study (isolate from 1 patient did not meet quality criteria).
cDNA synthesis, TaqMan Low Density Arrays and qPCR
600 ng of RNA (AAV: n = 20, controls: n = 12) were reversely transcribed for TaqMan low density arrays using the TaqMan® MicroRNA Reverse Transcription Kit and MegaplexTM Pools (Applied Biosystems, Carlsbad, USA). RNA samples (AAV: n = 30, controls: n = 12) were reversely transcribed for qPCR using the High-Capacity cDNA Reverse Transcription Kit with RNase Inhibitor from Applied Biosystems. Reverse transcription was performed for both on a 2720 Thermal Cycler (Applied Biosystems, Carlsbad, USA).
MiR expression was assessed by TaqMan Low Density Arrays (Human Pool A, Applied Biosystems, Carlsbad, USA) using RNU48 and Mamm U6 as endogenous controls. Arrays were run on a 7900HT Fast Real-time PCR System (Applied Biosystems) with ROX as reference dye. Expression analysis was computed by Expression suite software (LifeTechnologies, Carlsbad, USA) applying an auto-threshold method. Cycle threshold (CT) values >32 were excluded from analysis.
Quantitative PCR was performed in triplicate using FAM labelled TaqMan assays and Fast Advanced Master Mix (both purchased from Applied Biosystems, Carlsbad, USA) on a StepOneplus thermocycler (for FIH-1 and HIF-1α mRNA, FIH-1: TaqMan assay Hs 00215495_m1, HIF-1α: TaqMan assay Hs 00153153_m1) or 7900HT Fast Real-time PCR System (for miR-31, hsa-miR-31–5p) setting ROX as reference dye. HPRT1 (TaqMan assay Hs 02800695_m1), and RNU48 (SNORD48, Homo sapiens small nucleolar RNA, C/D box 48) served as endogenous controls for FIH-1, HIF-1α mRNA and miR-31, respectively. Comparative CT values were calculated with Expression suite software (FIH-1 mRNA, LifeTechnologies, Carlsbad, USA), and RQ Manager software (miR-31, Applied Biosystems, Carlsbad, USA) by auto-threshold. Relative quantification was performed by using the ΔΔCT method with healthy controls as normaliser.
Statistical analysis
Statistics were calculated with Graphpad prism 5 software. Since the data failed to show homoscedasticity and did not meet criteria of Gaussian distribution, non-parametric two-sided Mann-Whitney-U-test was applied for comparison of two groups. Correlation analysis was performed using Spearman’s rank correlation coefficient. All data in figures are given as mean ± standard deviation (s.d.). A p-value ≤ 0.05 was considered as significant. The datasets generated and analysed during the current study are available from the corresponding author on reasonable request.
References
Heine, P. et al. The C-terminal cysteine-rich region dictates specific catalytic properties in chimeras of the ectonucleotidases NTPDase1 and NTPDase2. Eur J Biochem 268, 364–373, https://doi.org/10.1046/j.1432-1033.2001.01896.x (2001).
Strater, N. Ecto-5′-nucleotidase: Structure function relationships. Purinergic signalling 2, 343–350, https://doi.org/10.1007/s11302-006-9000-8 (2006).
Buckley, R. H. Molecular defects in human severe combined immunodeficiency and approaches to immune reconstitution. Annu Rev Immunol 22, 625–655, https://doi.org/10.1146/annurev.immunol.22.012703.104614 (2004).
Mandapathil, M. et al. CD26 expression and adenosine deaminase activity in regulatory T cells (Treg) and CD4( + ) T effector cells in patients with head and neck squamous cell carcinoma. Oncoimmunology 1, 659–669, https://doi.org/10.4161/onci.20387 (2012).
Brady, T. Adenosine deaminase. Biochem J 36, 478–484 (1942).
Dong, R. P. et al. Determination of adenosine deaminase binding domain on CD26 and its immunoregulatory effect on T cell activation. Journal of immunology 159, 6070–6076 (1997).
Zhong, J., Rao, X. & Rajagopalan, S. An emerging role of dipeptidyl peptidase 4 (DPP4) beyond glucose control: potential implications in cardiovascular disease. Atherosclerosis 226, 305–314, https://doi.org/10.1016/j.atherosclerosis.2012.09.012 (2013).
Cortes, A. et al. Moonlighting adenosine deaminase: a target protein for drug development. Med Res Rev 35, 85–125, https://doi.org/10.1002/med.21324 (2015).
Faas, M. M., Saez, T. & de Vos, P. Extracellular ATP and adenosine: The Yin and Yang in immune responses? Mol Aspects Med, doi:https://doi.org/10.1016/j.mam.2017.01.002 (2017).
Hasko, G. & Pacher, P. A2A receptors in inflammation and injury: lessons learned from transgenic animals. Journal of leukocyte biology 83, 447–455, https://doi.org/10.1189/jlb.0607359 (2008).
Hasko, G., Linden, J., Cronstein, B. & Pacher, P. Adenosine receptors: therapeutic aspects for inflammatory and immune diseases. Nat Rev Drug Discov 7, 759–770, https://doi.org/10.1038/nrd2638 (2008).
Synnestvedt, K. et al. Ecto-5′-nucleotidase (CD73) regulation by hypoxia-inducible factor-1 mediates permeability changes in intestinal epithelia. The Journal of clinical investigation 110, 993–1002, https://doi.org/10.1172/JCI15337 (2002).
Kiss, J. et al. IFN-beta protects from vascular leakage via up-regulation of CD73. European journal of immunology 37, 3334–3338, https://doi.org/10.1002/eji.200737793 (2007).
Regateiro, F. S. et al. Generation of anti-inflammatory adenosine by leukocytes is regulated by TGF-beta. European journal of immunology 41, 2955–2965, https://doi.org/10.1002/eji.201141512 (2011).
Narravula, S., Lennon, P. F., Mueller, B. U. & Colgan, S. P. Regulation of endothelial CD73 by adenosine: paracrine pathway for enhanced endothelial barrier function. Journal of immunology 165, 5262–5268, https://doi.org/10.4049/jimmunol.165.9.5262 (2000).
Hu, J. et al. The role of the miR-31/FIH1 pathway in TGF-beta-induced liver fibrosis. Clin Sci (Lond) 129, 305–317, https://doi.org/10.1042/CS20140012 (2015).
Wong, H. K. et al. The Cancer Genome Atlas Analysis Predicts MicroRNA for Targeting Cancer Growth and Vascularization in Glioblastoma. Mol Ther 23, 1234–1247, https://doi.org/10.1038/mt.2015.72 (2015).
Li, Y. et al. Hypoxia-inducible miR-182 enhances HIF1alpha signaling via targeting PHD2 and FIH1 in prostate cancer. Sci Rep 5, 12495, https://doi.org/10.1038/srep12495 (2015).
Li, G. et al. MicroRNA-125a-5p Contributes to Hepatic Stellate Cell Activation through Targeting FIH1. Cell Physiol Biochem 38, 1544–1552, https://doi.org/10.1159/000443095 (2016).
Blume, C. et al. Autoimmunity in CD73/Ecto-5′-nucleotidase deficient mice induces renal injury. PloS one 7, e37100, https://doi.org/10.1371/journal.pone.0037100 (2012).
Liang, D. et al. CD73 Expressed on gammadelta T Cells Shapes Their Regulatory Effect in Experimental Autoimmune Uveitis. PloS one 11, e0150078, https://doi.org/10.1371/journal.pone.0150078 (2016).
Hay, C. M. et al. Targeting CD73 in the tumor microenvironment with MEDI9447. Oncoimmunology 5, e1208875, https://doi.org/10.1080/2162402X.2016.1208875 (2016).
Montalban Del Barrio, I. et al. Adenosine-generating ovarian cancer cells attract myeloid cells which differentiate into adenosine-generating tumor associated macrophages - a self-amplifying, CD39- and CD73-dependent mechanism for tumor immune escape. J Immunother Cancer 4, 49, https://doi.org/10.1186/s40425-016-0154-9 (2016).
Mills, J. H. et al. CD73 is required for efficient entry of lymphocytes into the central nervous system during experimental autoimmune encephalomyelitis. Proceedings of the National Academy of Sciences of the United States of America 105, 9325–9330, https://doi.org/10.1073/pnas.0711175105 (2008).
Mahamed, D. A., Mills, J. H., Egan, C. E., Denkers, E. Y. & Bynoe, M. S. CD73-generated adenosine facilitates Toxoplasma gondii differentiation to long-lived tissue cysts in the central nervous system. Proceedings of the National Academy of Sciences of the United States of America 109, 16312–16317, https://doi.org/10.1073/pnas.1205589109 (2012).
Jennette, J. C. et al. 2012 revised International Chapel Hill Consensus Conference Nomenclature of Vasculitides. Arthritis and rheumatism 65, 1–11, doi:https://doi.org/10.1002/art.37715 (2013).
Berden, A. E. et al. Histopathologic classification of ANCA-associated glomerulonephritis. Journal of the American Society of Nephrology: JASN 21, 1628–1636, https://doi.org/10.1681/ASN.2010050477 (2010).
Jennette, J. C., Wilkman, A. S. & Falk, R. J. Anti-neutrophil cytoplasmic autoantibody-associated glomerulonephritis and vasculitis. The American journal of pathology 135, 921–930 (1989).
Brouwer, E. et al. Predominance of IgG1 and IgG4 subclasses of anti-neutrophil cytoplasmic autoantibodies (ANCA) in patients with Wegener’s granulomatosis and clinically related disorders. Clinical and experimental immunology 83, 379–386, https://doi.org/10.1111/j.1365-2249.1991.tb05647.x (1991).
Abdulahad, W. H., van der Geld, Y. M., Stegeman, C. A. & Kallenberg, C. G. Persistent expansion of CD4 + effector memory T cells in Wegener’s granulomatosis. Kidney international 70, 938–947, https://doi.org/10.1038/sj.ki.5001670 (2006).
Rimbert, M. et al. Decreased numbers of blood dendritic cells and defective function of regulatory T cells in antineutrophil cytoplasmic antibody-associated vasculitis. PloS one 6, e18734, https://doi.org/10.1371/journal.pone.0018734 (2011).
Gan, P. Y. et al. Th17 cells promote autoimmune anti-myeloperoxidase glomerulonephritis. Journal of the American Society of Nephrology: JASN 21, 925–931, https://doi.org/10.1681/ASN.2009070763 (2010).
Chen, T. et al. MicroRNA-31 contributes to colorectal cancer development by targeting factor inhibiting HIF-1alpha (FIH-1). Cancer Biol Ther 15, 516–523, https://doi.org/10.4161/cbt.28017 (2014).
Mahon, P. C., Hirota, K. & Semenza, G. L. FIH-1: a novel protein that interacts with HIF-1alpha and VHL to mediate repression of HIF-1 transcriptional activity. Genes Dev 15, 2675–2686, https://doi.org/10.1101/gad.924501 (2001).
Saghiri, R. et al. Serum adenosine deaminase activity in patients with systemic lupus erythematosus: a study based on ADA1 and ADA2 isoenzymes pattern. Rheumatol Int 32, 1633–1638, https://doi.org/10.1007/s00296-011-1836-8 (2012).
Li, D. M. et al. [Expression of FOXP3 in CD4+ CD39+ T cells of patients with systemic lupus erythematosus and dynamic observation of treatment with glucocorticoid]. Zhonghua Yi Xue Za Zhi 89, 1636–1638 (2009).
Li, D. M. et al. [The expression of CD73 in CD4+ regulatory T cells in patients with new-onset systemic lupus erythematosus]. Zhonghua Nei Ke Za Zhi 49, 772–775 (2010).
Guzman-Flores, J. M. et al. Expression of CD73 and A2A receptors in cells from subjects with obesity and type 2 diabetes mellitus. Immunobiology 220, 976–984, https://doi.org/10.1016/j.imbio.2015.02.007 (2015).
Schuler, P. J. et al. CD4(+)CD73(+) T cells are associated with lower T-cell activation and C reactive protein levels and are depleted in HIV-1 infection regardless of viral suppression. Aids 27, 1545–1555, https://doi.org/10.1097/QAD.0b013e328360c7f3 (2013).
Peres, R. S. et al. Low expression of CD39 on regulatory T cells as a biomarker for resistance to methotrexate therapy in rheumatoid arthritis. Proceedings of the National Academy of Sciences of the United States of America 112, 2509–2514, https://doi.org/10.1073/pnas.1424792112 (2015).
Figueiro, F. et al. Methotrexate up-regulates ecto-5′-nucleotidase/CD73 and reduces the frequency of T lymphocytes in the glioblastoma microenvironment. Purinergic signalling 12, 303–312, https://doi.org/10.1007/s11302-016-9505-8 (2016).
Haglund, S., Vikingsson, S., Almer, S. & Soderman, J. Combination treatment with 6-mercaptopurine and allopurinol in HepG2 and HEK293 cells - Effects on gene expression levels and thiopurine metabolism. PloS one 12, e0173825, https://doi.org/10.1371/journal.pone.0173825 (2017).
Haglund, S., Almer, S., Peterson, C. & Soderman, J. Gene expression and thiopurine metabolite profiling in inflammatory bowel disease - novel clues to drug targets and disease mechanisms? PloS one 8, e56989, https://doi.org/10.1371/journal.pone.0056989 (2013).
Zhao, Y. et al. Circulating T follicular helper cell and regulatory T cell frequencies are influenced by B cell depletion in patients with granulomatosis with polyangiitis. Rheumatology 53, 621–630, https://doi.org/10.1093/rheumatology/ket406 (2014).
Ahlmann, M. & Hempel, G. The effect of cyclophosphamide on the immune system: implications for clinical cancer therapy. Cancer Chemother Pharmacol 78, 661–671, https://doi.org/10.1007/s00280-016-3152-1 (2016).
Marinaki, S. et al. Abnormalities of CD4 T cell subpopulations in ANCA-associated vasculitis. Clinical and experimental immunology 140, 181–191, https://doi.org/10.1111/j.1365-2249.2005.02731.x (2005).
Kalsch, A. I., van der Woude, F. J. & Yard, B. A. The role of T lymphocytes in ANCA associated vasculitis: neglected or revisited? Clin Nephrol 64, 471–474, https://doi.org/10.5414/CNP64471 (2005).
Kalsch, A. I. et al. Use of highly sensitive C-reactive protein for followup of Wegener’s granulomatosis. The Journal of rheumatology 37, 2319–2325, https://doi.org/10.3899/jrheum.100302 (2010).
Kinsey, G. R. et al. Autocrine adenosine signaling promotes regulatory T cell-mediated renal protection. Journal of the American Society of Nephrology: JASN 23, 1528–1537, https://doi.org/10.1681/ASN.2012010070 (2012).
Castrop, H. et al. Impairment of tubuloglomerular feedback regulation of GFR in ecto-5′-nucleotidase/CD73-deficient mice. The Journal of clinical investigation 114, 634–642, https://doi.org/10.1172/JCI21851 (2004).
Koszalka, P. et al. Targeted disruption of cd73/ecto-5′-nucleotidase alters thromboregulation and augments vascular inflammatory response. Circulation research 95, 814–821, https://doi.org/10.1161/01.RES.0000144796.82787.6f (2004).
Zhao, M., Wang, Z., Yung, S. & Lu, Q. Epigenetic dynamics in immunity and autoimmunity. Int J Biochem Cell Biol 67, 65–74, https://doi.org/10.1016/j.biocel.2015.05.022 (2015).
Skoglund, C. et al. Circulating microRNA expression pattern separates patients with anti-neutrophil cytoplasmic antibody associated vasculitis from healthy controls. Clinical and experimental rheumatology 33, S-64–71 (2015).
Bertram, A. et al. Circulating ADAM17 Level Reflects Disease Activity in Proteinase-3 ANCA-Associated Vasculitis. Journal of the American Society of Nephrology: JASN 26, 2860–2870, https://doi.org/10.1681/ASN.2014050477 (2015).
Kurz, T. et al. A myelopoiesis gene signature during remission in anti-neutrophil cytoplasm antibody-associated vasculitis does not predict relapses but seems to reflect ongoing prednisolone therapy. Clinical and experimental immunology 175, 215–226, https://doi.org/10.1111/cei.12236 (2014).
Kaller, M. et al. Genome-wide characterization of miR-34a induced changes in protein and mRNA expression by a combined pulsed SILAC and microarray analysis. Mol Cell Proteomics 10(M111), 010462, https://doi.org/10.1074/mcp.M111.010462 (2011).
Ghosh, G. et al. Hypoxia-induced microRNA-424 expression in human endothelial cells regulates HIF-alpha isoforms and promotes angiogenesis. The Journal of clinical investigation 120, 4141–4154, https://doi.org/10.1172/JCI42980 (2010).
Wilde, B. et al. T cells in ANCA-associated vasculitis: what can we learn from lesional versus circulating T cells? Arthritis research & therapy 12, 204, https://doi.org/10.1186/ar2923 (2010).
Lintermans, L. L., Stegeman, C. A., Heeringa, P. & Abdulahad, W. H. T cells in vascular inflammatory diseases. Frontiers in immunology 5, 504, https://doi.org/10.3389/fimmu.2014.00504 (2014).
Komocsi, A. et al. Peripheral blood and granuloma CD4( + )CD28(-) T cells are a major source of interferon-gamma and tumor necrosis factor-alpha in Wegener’s granulomatosis. The American journal of pathology 160, 1717–1724, https://doi.org/10.1016/S0002-9440(10)61118-2 (2002).
Muller, A. et al. Localized Wegener’s granulomatosis: predominance of CD26 and IFN-gamma expression. The Journal of pathology 192, 113–120, https://doi.org/10.1002/1096-9896(2000)9999:9999 < ::AID-PATH656 > 3.0.CO;2-M (2000).
Degel, J. & Shokrani, M. Validation of the efficacy of a practical method for neutrophils isolation from peripheral blood. Clinical laboratory science: journal of the American Society for Medical Technology 23, 94–98 (2010).
Author information
Authors and Affiliations
Contributions
L.K., U.B., B.K.K., B.Y. and A.I.K. designed the study. L.K., B.Y., A.I.K., U.B., B.K.K. and S.P. participated in writing and revision of the manuscript. U.B., A.I.K., and B.K.K. recruited patients and provided clinical data. L.K., A.B., S.P., L.L. and M.H. performed experiments. L.K. performed statistical analysis.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kling, L., Benck, U., Breedijk, A. et al. Changes in CD73, CD39 and CD26 expression on T-lymphocytes of ANCA-associated vasculitis patients suggest impairment in adenosine generation and turn-over. Sci Rep 7, 11683 (2017). https://doi.org/10.1038/s41598-017-12011-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-12011-4
This article is cited by
-
CD73-mediated adenosine production by CD8 T cell-derived extracellular vesicles constitutes an intrinsic mechanism of immune suppression
Nature Communications (2021)
-
Multiple steps determine CD73 shedding from RPE: lipid raft localization, ARA1 interaction, and MMP-9 up-regulation
Purinergic Signalling (2018)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
