
Abstract
Much of the fossil record for dogs consists of mandibles. However, can fossil canid mandibles be reliably identified as dogs or wolves? 3D geometric morphometric analysis correctly classifies 99.5% of the modern dog and wolf mandibles. However, only 4 of 26 Ust’-Polui fossil mandibles, a Russian Arctic site occupied from 250BCE to 150CE, were identified as dogs and none of the 20 Ivolgin mandibles, an Iron Age site in southern Russia, were identified as dogs. Three of the Ust’-Polui mandibles and 8 of the Ivolgin mandibles were identified as wolves. In contrast, all 12 Ivolgin skulls and 5 Ust’-Polui skulls were clearly identified as dogs. Only the classification of the UP6571 skull as a dog (Dog Posterior Probability = 1.0) was not supported by the typical probability. Other evidence indicates these canids were domesticated: they were located within human dwellings, remains at both sites have butchery marks indicating that they were consumed, and isotope analysis of canid and human remains from Ust’-Polui demonstrate that both were consuming freshwater protein; indicating that the humans were feeding the canids. Our results demonstrate that the mandible may not evolve as rapidly as the cranium and the mandible is not reliable for identifying early dog fossils.
Similar content being viewed by others

Introduction
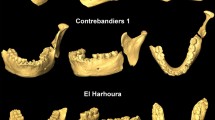
The date and location of dog domestication is a contentious issue whether the evidence being considered is genetic or morphological1,2,3,4,5,6,7,8,9,10,11,12,13,14. Previous research on canid fossil mandibles and skulls has employed Euclidean distances for identification1, 3, 7. We conducted a three-dimensional geometric morphometric analysis of fossil mandibles and skulls from Ust’-Polui, Ivolgin, and Alaska to determine whether these are dog or wolf fossils6, 15,16,17,18,19. The Ust’-Polui and Ivolgin sites date to the late Holocene, post-dating the advent of dog domestication by millennia16,17,18,19. The Ust’-Polui archaeological site is in Salekhard, Russia, in the Arctic (66.5501°N, 66.6028°E). Ust’-Polui has produced thousands of artifacts and faunal remains, including disarticulated skeletal elements from over 100 canids, nearly all of which were originally identified as dogs based on their relative small sizes compared to Arctic wolves18, 19. At least two of the canid skeletons found at this site were fully articulated burials (these canids were not available for the present analysis). Radiocarbon dating and dendrochronology indicate this fortified site was occupied from ~250BCE to 150CE by foragers18, 19. The Ivolgin site is in the steppe region of southern Russia near Ulan-Ude (51.7630°N, 107.47346°E). Ivolgin consisted of a series of earth ramparts and a wood stockade that surrounded over 50 wooden dwellings16, 17. This town was probably occupied by the Xiongnu, a confederation of Iron Age pastoral groups, from ~300BCE to 200CE16, 17. Scattered faunal remains were found throughout Ivolgin, with over 90% identified as domestic animals (sheep, cattle, pig, horse, goat, camel, yak, dog). At least 59 specimens from Ivolgin were previously identified as dogs based on size comparisons with southern Siberian wolves16. The only wild canid identified was fox16. At both Ust’-Polui and Ivolgin, some of the canid remains display butchery marks, indicating they were consumed by people. Additionally, we analyzed carbon-dated late Pleistocene canid fossils from Alaskan permafrost deposits that were all genetically classified as wolves20. Finally, Alaskan canids carbon-dated near 1600CE and genetically classified as dogs also were analyzed21. These two sets of Alaskan canid fossils were included to determine whether genetic identifications correspond with those based on morphology.
Traditional morphometric analyses of fossil canid mandibles and skulls aimed at identifying dogs depend mostly on one-dimensional measures of length and width1,2,3,4,5, 7. As demonstrated in our previous analysis of the canid skull6, distance measurements are problematic for multiple reasons including: autocorrelation, isometry, overlap between dogs and wolves (making identification impossible), spurious correlations in multivariate analyses, and non-normal distributions22. Capturing 3D coordinates from mandibles and skulls provides a more accurate representation of their inherent 3D shapes than one-dimensional distances and ratios (Supplementary Figure S1)23, 24. Geometric morphometric methods (GMM) are widely recognized as powerful and sophisticated diagnostic tools for investigating biological shape6, 15. Procrustes superimposition of the coordinate configurations removes information related to size by scaling all configurations to the same centroid size while translating and rotating the landmark configurations using a least-squares fit of homologous landmarks24, 25. Thus, Procrustes coordinates only contain information pertaining to shape, having removed information related to size, position, and rotation24, 25. Here we re-analyze the canid skulls and mandibles from the Ust’-Polui, Ivolgin, and Alaska sites using 3D GMM to assess whether they can be accurately identified as dogs or wolves. We compared the fossil mandibles and skulls to a large dataset of modern mesaticephalic (wolf-like) dogs and a global assemblage of both modern and fossil wolves. We hypothesize that the fossil skulls will be accurately identified as wolves or dogs; however, it remains to be seen if these fossil mandibles can be reliably categorized.
The form-space principal components analysis (PCA) revealed that modern dog and wolf mandibles are separated within the first three principal components, which account for 92.3% of the total mandible form-space variance (Fig. 1a). PC1 captures static allometry and is associated with overall size variation (rPC1 = 0.99, P < 0.001) from the large, robust jaws of wolves and dog breeds such as German Shepherds to the smaller mandibles of breeds like the Fox Terrier (Fig. 1b and Supplementary Video S1). Wolves have significantly larger mandibles than dogs (P < 0.001; permutation test, n = 10,000). However, due to their considerable variation in size, dogs overlap with wolves along PC1 (Fig. 1a). Dogs diverge from wolves along PCs 2 and 3, which are both independent of size (rPC2 = 0.002, rPC3 = 0.003). The curvature of the mandible distinguishes dogs from wolves along PC2 (Fig. 1b and Supplementary Video S2).

3D plot of PC1–3 mandible shape variation. Black: dogs, dark grey: Alaskan wolves, light grey: European wolves, dark red: Ivolgin fossils, green: Ust’-Polui fossils, purple: Pleistocene Alaskan wolves, cyan: 1600CE fossil dogs, orange: unknown Alaskan fossil canids, pink: 1600CE fossil wolf.
99.5% of modern canid mandibles were correctly identified as either dogs or wolves with 100% accuracy using a resampling procedure involving one-thousand iterations of a cross-validation Quadratic Discriminant Analysis (QDA) to ensure equal dog and wolf sample sizes, with posterior probability greater than 0.90 (Tau = 0.460, Wilks’ lambda = 0.12). Furthermore, the classifications were supported by the typical probabilities.
The fossil canid mandibles exhibit a wide variation in morphology; some are separated from both wolves and dogs at the positive end of PC1 while some are within the wolf or dog shape variation (Fig. 1a). The results from the classification procedure identified 4 of the 26 mandibles from Ust’-Polui as dogs and 3 of the mandibles as wolves (Table 1). The remaining 19 Ust’-Polui mandibles could not be statistically identified as dogs or wolves (Table 1). Within the Ivolgin population, 8 mandibles were classified as wolves and 12 were unidentifiable (Table 1). The unclassified Ust’-Polui and Ivolgin mandibles are not highly aberrant from dogs or wolves as demonstrated by their close proximity on the PCA plot (Fig. 1a). These unclassified mandibles do not share the formspace of either the dogs or the wolves. They are similar in size to smaller mesaticephalic dogs such as Fox Terriers (overlap on PC1) but the shape of their mandibles resembles the wolf mandibles (overlap on PC2). Our GMM analysis confirmed that the Alaskan Pleistocene canid mandibles are wolves (Table 1)20. Our results also verified that 2 of the Alaskan mandibles dated near 1600CE (AMNH30436 and AMNH70932) are dogs (Table 1)21. Our analysis classified the mandibles AMNH30482 and AMNH70963C as wolves despite the genetic identification by Leonard et al.21 of both specimens being dogs (Table 1). We also analyzed 7 mandibles that have not been carbon-dated nor genetically identified but were found at the same Alaskan site as the 1600CE mandibles. One of these, AMNH39381, was classified as a dog, and 6 of the mandibles were identified as wolves (Table 1).
We conducted a separate three-dimensional geometric morphometric analysis of the fossil skulls from Ivolgin, Ust’-Polui, and Alaska and compared them to modern mesaticephalic (wolf-like) dogs, ancient dogs, and the global assemblage of both modern and fossil wolves from our previous study6. The form-space PCA clearly shows the fossil canid skulls from Ust’-Polui and Ivolgin all lie within dog cranial shape variation and are clearly separated from wolf cranial shape variation within the first three principal components which account for 89.7% of the total skull form variance (Fig. 2a). The Ust’-Polui and Ivolgin fossil skulls all share with modern dogs forward-facing orbits and almost all display a pronounced angle between the forehead and the muzzle, a distinguishing feature of dog skulls (Fig. 2b)6. The results from the classification procedure identify all fossil skulls as dogs according to both the posterior and typical probabilities in 99.6% of the 1,000 resampling runs (Table 2). Only the classification of UP6571 as a dog via posterior probability (Dog Posterior Probability = 1.0) was not supported by the typical probability in more than 95% of the resampling runs (Table 2). The late Pleistocene Alaskan fossil canid skulls were all confirmed to be wolves except for AMNH30433 (wolf posterior probability = 0.87) and AMNH67157 (wolf posterior probability = 0.66). The Alaskan fossil canids from 1600CE were all identified as dogs in our analysis, corroborating the genetic identification of these specimens as dogs21.

3D plot of PC1–3 skull shape variation. Black: dogs, dark grey: Alaskan wolves, light grey: European wolves, dark red: Ivolgin fossils, green: Ust’-Polui fossils, purple: Pleistocene Alaskan wolves, cyan: 1600CE fossil dogs.
Surprisingly, only 15% of the mandible specimens from the Ust’-Polui site were classified as dogs despite the very high accuracy of this procedure in correctly assigning known specimens to their group. In addition, 69% of the mandible specimens from Ust’-Polui and 60% of the mandible specimens from Ivolgin were unclassified as either dogs or wolves. The unclassified mandibles are found outside the wolf mandible morphospace because they are smaller than the wolves and outside the dog mandible morphospace because of their wolf-like shape. However, all but one of the skull specimens from both sites were identified as dogs. Interestingly, we saw a similar pattern in the fossil canids from Alaska. Although 4 of 1600CE Alaskan fossil mandibles had been genetically identified as dogs21, our analysis only classified 2 of these mandibles as dogs (Table 1). Furthermore, the canids at Ust’-Polui and Ivolgin are suspected to be domestic dogs based on other criteria. Those at Ivolgin were found in association with remains of many other domestic fauna and in a fortified town occupied by a historically-documented pastoral society, some inside houses and waste pits. Stable carbon and nitrogen isotope analysis of bone collagen from 44 Ust’-Polui canid specimens, including 10 crania and 34 right scapulae, indicated these individuals have very negative δ13C values (mean δ13C = −25.8‰, s.d. = 0.8) and elevated δ15N values (mean δ15N = 13.9‰, s.d. = 0.8) (Supplementary Table S1; For isotope analysis methods see Supplementary Information). The canids’ isotope values are similar to those of two humans buried at Ust’-Polui (mean δ13C = −25.3‰, s.d. = 1.25; mean δ15N = 16.9‰, s.d. = 0.4; Supplementary Figure S2). Terrestrial herbivores such as reindeer (Rangifer tarandus) and elk (Alces alces) at the site have far more positive δ13C values (mean δ13C = −20.0‰, s.d. = 1.03), and much lower δ15N values (mean δ15N = 5.6‰, s.d. = 2.1). Bone collagen δ15N values show enrichment of 3–5‰ along the food chain, providing an indication of trophic level26, 27. The canids and humans at Ust’-Polui, all with δ15N values above 12‰, were regularly consuming food items with higher δ15N values than those of these large-bodied terrestrial herbivores. In the Arctic, such elevated δ15N values appear more consistent with dietary reliance on freshwater or marine fauna. Neither freshwater fish or marine mammals are well represented in our isotope data, but other studies show that Arctic freshwater fish have far more negative δ13C values than marine mammals and fish, ranging from around −30‰ to −20‰, with the two groups of marine fauna often having more positive values than terrestrial ungulates28,29,30,31,32,33,34,35. Offsets in δ13C between prey collagen and predator collagen are around 1‰26, 36. The very negative δ13C values for the canids and humans at Ust’-Polui suggest both were regularly consuming freshwater fish; remains of such fish are highly abundant at the site19. Such dietary patterns are inconsistent with those of wolves living in the Arctic, where water bodies are frozen for much of the year37, 38, but are consistent with people partially provisioning dogs with their own food items, a historically well-documented practice in many areas39.
The lack of consistent identification of the mandibles as either dogs or wolves could indicate several things. The mandibles seem unlikely to be from small wolves or hybrids because neither wolf nor hybrid crania were found at either site; all of the skulls from both sites were clearly identified as dogs. It also seems improbable that the mandibles are from some other type of canid, as one would expect their crania also to be present at these sites, both of which have been extensively excavated. Perhaps most telling, even some of the Late Holocene mandibles from Alaska were not morphologically identified, despite the identification of the skulls as dogs as well as their confirmation as dogs from genetic information21.
Contextual and dietary information from both Ivolgin and Ust’-Polui provide supporting evidence for the presence of dogs at these two sites. The mandibles from Ivolgin were found with remains other domestic animals within a fortified town. Analysis of the canid skulls at both Ust’-Polui and Ivolgin confirm that numerous dogs are present at both locations but failed to show the presence of wolves. Further, there are butchery marks on the canid remains at both of these sites, and the isotope analysis indicates that the canids at Ust’-Polui had similarly structured diets as the humans buried there, and these diets included freshwater fish.
Overall, these results indicate that the rate of evolutionary modification of the dog mandible may not keep pace with cranial shape change, and that the variation in mandible shape that differentiates modern dogs and wolves mostly emerged relatively late in the domestication process, perhaps with the advent of modern intensive breeding. Fossil mandibles from even Late Holocene dogs may be mistakenly classified as small wild canids and should not be relied on as the only evidence for specimen identification. Future studies comparing shape variation in canid skulls and mandibles may elucidate the lack of coevolution observed in our dataset.
Methods
Ct-scans of fossil mandibles from Ivolgin and Ust’-Polui were converted into Polygon files and digitized in IDAV Landmark software40 all other specimens were digitized by AGD with a Microscribe digitizer. Fossils include 26 mandibles from Ust’-Polui18, 19, 20 from Ivolgin16, 17, 8 late Pleistocene canids from Alaska20 (AMNH: 67168, 67169, 67179, 67202, 67224, 67228, 70944, 70958), 4 canids from near 1600CE Alaska21 (AMNH: 30436, 30482, 70932, 70963C), and another 7 canids that are likely from around 1600 CE Alaska (but have not been carbon-dated) (AMNH: 30474, 39381, 67242, 97104, 97105, AINS825, AINS840). 37 three-dimensional coordinates were captured from the mandibles of 121 North American wolves, 85 Eurasian wolves, and 240 adult dogs (only mesaticephalic breeds) (Supplementary Figure S1). Breeds in the mandible analysis include: Afghan Hound, Airedale Terrier, Akita Inu, Alaskan Malmute, Bloodhound, Borzoi, Boxer, Bull Terrier, Chesapeake Bay Retriever, Chow Chow, Cocker Spaniel, Dalmatian, Dingo, English Bulldog, English Setter, English Springer Spaniel, Foxterrier, French Mastiff, German Shepherd, German Spaniel, Golden Retriever, Greenland Dog, Greyhound, Irish Setter, Irish Wolfhound, Jura Laufhund, Jura Laufhund St. Hubert, Labrador Retriever, Nova Scotia Duck Tolling Retriever, Pharoh Hound, Poodle, Samojede, Scottish Deerhound, Shar Pei, Siberian Husky, Tervueren, Weimeraner, Whippet, and Wolfspitz.
Ct-scans of fossil skulls from Ivolgin and Ust’-Polui, as well as the following fossil specimens: Eliseevichi MAE 447/5298 (13,905 +/− 55 YBP; Epigravettian), Goyet (31,680 +/− 250 YBP), Trou Balleux (10,110 +/− 120 YBP), Shamanka II (7,372 +/− 47 YBP), and Ust’-Belaia (6,817 +/− 63 YBP) were converted into Polygon files and digitized in IDAV Landmark software40 all other specimens were digitized by AGD with a Microscribe digitizer. Fossils include 12 skulls from Ust’-Polui (UP: 1007, 1202, 1203, 2531, 4767, 5070, 5071, 6433, 6517, 6554, 6571, 6596)18, 19, 6 fossil skulls from Ivolgin (IV: 35470_1, 35470_2, 35470_4, 35470_5, 35470_7, 35470_8)16, 17, 4 late Pleistocene skulls from Alaska20 (AMNH: 30431, 30450, 67163, 97079), and 4 skulls from near 1600CE Alaska21 (AMNH: 30435, 30436, 67155a, 70932). 36 three-dimensional coordinates were recorded from skulls of 258 North American wolves, 57 European wolves, and 91 adult dogs (only mesaticephalic breeds). The following fossils were included in the wolf sample: Eliseevichi MAE 447/5298, Goyet, and Trou Balleux. The following fossils were included in the dog sample: Shamanka II, Ust’-Belaia, three Egyptian mummified dogs from the Saite–Ptolemaic period, and four Neolithic and one Gallo-Roman dog from France. For more details on the specimens used in the cranial analysis please see Drake et al.6.
The majority of dog specimens are housed in the Albert Heim Collection at the Natural History Museum in Berne, Switzerland. Most of the wolf specimens are from the University of Alaska Museum in Fairbanks, Alaska. Other specimens are from the Smithsonian Institution’s National Museum of Natural History in Washington, DC, the Museum of Vertebrate Zoology at the University of California in Berkeley, California, the Natural History Museum in Berne, Switzerland, and the Zoology Department at the Natural History Museum, London.
Geometric morphometric analysis23,24,25, 41 was conducted with the R programming language. Landmark software was used to warp a 3D Ct-scan of a wolf mandible and a wolf skull to the average shape of the known dog and wolf specimens and then warped along the PC axes40. Many of the methods used in this analysis are similar to those used in Drake et al.6. Here we detail any differences in methodology.
We used a resampling procedure to balance the sample sizes of the wolf and dog groups. A test developed by Anderson determined that eigenvalues from PC 9 onwards were nearly equal and therefore not useful for our analysis. We ran 1,000 iterations of the resampling procedure and in each round we used the Anderson test to determine whether the first 9 PCs were useful. If they were not, we eliminated that round and ran the resampling procedure again. For each successful round of resampling we used the 9 PCs to conduct a quantitative discriminant function analysis (QDFA) of dogs and wolves. The posterior probability and typical probabilities were calculated in the same manner as in Drake et al.6. For the analysis of the skulls we followed the same procedure, using PCs 1–6.
Data Availability
The datasets analysed during the current study are available from the corresponding author on reasonable request.
References
Davis, S. J. M. & Valla, F. R. Evidence for domestication of the dog 12,000 years ago in the Natufian of Israel. Nature 276, 608–610 (1978).
Germonpré, M., Lázničková-Galetová, M. & Sablin, M. V. Palaeolithic dog skulls at the Gravettian Předmostí site, the Czech Republic. J. Archaeol. Sci. 39.1, 184–202 (2012).
Ovodov, N. D. et al. A 33,000-year-old incipient dog from the Altai Mountains of Siberia: evidence of the earliest domestication disrupted by the Last Glacial Maximum. PLoS One 6(7), e22821, doi:10.1371/journal.pone.0022821 (2011).
Germonpré, M. et al. Fossil dogs and wolves from Palaeolithic sites in Belgium, the Ukraine and Russia: osteometry, ancient DNA and stable isotopes. J. Archaeol. Sci. 36, 473–490 (2009).
Sablin, M. V. & Khlopachev, G. A. The earliest Ice Age dogs: evidence from Eliseevichi I. Current Anthropology 43, 795–799 (2002).
Drake, A. G., Coquerelle, M. & Colombeau, G. 3D morphometric analysis of fossil canid skulls contradicts the suggested domestication of dogs during the late Paleolithic. Scientific Reports 5, 8299 (2015).
Germonpré, M., Lázničková-Galetová, M., Losey, R. J., Räikkönen, J. & Sablin, M. V. Large canids at the Gravettian Předmostí site, the Czech Republic: the mandible. Quaternary International 359, 261–279 (2015).
Frantz, L. A. F. et al. Genomic and archaeological evidence suggest a dual origin of domestic dogs. Science 352, 1228–1231 (2016).
Thalmann, O. et al. Complete Mitochondrial Genomes of Ancient Canids Suggest a European Origin of Domestic Dogs. Science 342, 871–874 (2013).
Wang, G. et al. The genomics of selection in dogs and the parallel evolution between dogs and humans. Nature Communications 4, 1860 (2013).
Druzhkova, A. S. et al. Ancient DNA analysis affirms the canid from Altai as a primitive dog. PloS One 8(no. 3), e57754, doi:10.1371/journal.pone.0057754 (2013).
Larson, G. et al. Rethinking dog domestication by integrating genetics, archeology, and biogeography. Proc. Natl. Acad. Sci. USA 109, 8878–8883 (2012).
Skoglund, P., Gotherstrom, A. & Jakobsson, M. Estimation of population divergence times from non-overlapping genomic sequences: examples from dogs and wolves. Mol. Biol. Evol. 28, 1505–1517 (2011).
Pang, J. F. et al. mtDNA data indicate a single origin for dogs south of Yangtze River, less than 16,300 years ago, from numerous wolves. Mol. Biol. Evol. 26, 2849–2864 (2009).
Rohlf, F. J. & Marcus, L. F. A revolution in morphometrics. Trends Ecol. Evol. 8, 129–132 (1993).
Davydova, A. The Ivolga Archaeological Complex, Volume 1, The Ivolga Fortress. (Asiatic Fund, 1995).
Brosseder, U. & Miller, B. K. Xiongnu Archaeology: Multidisciplinary Perspectives of the First Steppe Empire in Inner Asia (Vor-und Frühgeschichtliche Archäologie Rheinische Friedrich-Wilhelms-Universität Bonn, 2011).
Chernetsov, V. N. & Moszyńska, W. Prehistory of Western Siberia (Arctic Institute of North America, 1974).
Gusev, A. V. & Fedorova, N. V. Drevnee Sviatilishche Ust’-Polui: Konstruktsii, Deistviia, Artefakty. Itogi Issledovanii Planigrafii i Stratigrafii Pamiatnika: 1935–2012 gg. (Izd-vo GU “Beverone Izdatel’stvo”, 2012).
Leonard, J. A. et al. Megafaunal extinctions and the disappearance of a specialized wolf ecomorph. Current Biology 17, 1146–1150 (2007).
Leonard, J. A. et al. Ancient DNA evidence for Old World origin of New World dogs. Science 298, 1613–1616 (2002).
Atchley, W. R., Gaskins, C. T. & Anderson, D. Statistical properties of ratios. I. Empirical results. Syst. Biol. 25, 137–148 (1976).
Strauss, R. E. & Bookstein, F. L. The truss: body form reconstructions in morphometrics. Systematic Biology 31.2, 113–135 (1982).
Mitteroecker, P. & Gunz, P. Advances in geometric morphometrics. Evolutionary Biology 36(no. 2), 235–247 (2009).
Dryden, I. L. & Mardia, K. V. Statistical shape analysis (Wiley, 1998).
Bocherens, H. & Drucker, D. Trophic Level Isotopic Enrichment of Carbon and Nitrogen in Bone Collagen: Case Studies from Recent and Ancient Terrestrial Ecosystems. International Journal of Osteoarchaeology 13, 46–53 (2003).
Schoeninger, M. J. & DeNiro, M. J. 1984. Nitrogen and Carbon Isotopic Composition of Bone Collagen from Marine and Terrestrial Animals. Geochimica et Cosmochimica Acta 48, 625–39 (1984).
Dehn, L. A. et al. Feeding ecology of phocid seals and some walrus in the Alaskan and Canadian Arctic as determined by stomach contents and stable isotope analysis. Polar Biology 30, 167–181 (2006).
Matley, J. K., Fisk, A. T. & Dick, T. D. Foraging Ecology of Ringed Seals (Pusa Hispida), Beluga Whales (Delphinapterus Leucas) and Narwhals (Monodon Monoceros) in the Canadian High Arctic Determined by Stomach Content and Stable Isotope Analysis. Polar Research 34 (2015).
Klervia, J., Szpak, P., & Richards, M. P. Zinc Isotope Ratios as Indicators of Diet and Trophic Level in Arctic Marine Mammals. PLoS One 11 (2016).
Hoekstra, P. F. et al. Trophic transfer of persistent organochloride contaminants (OCs) within an Arctic marine food web from the southern Beaufor-Chukchi Seas. Environmental Pollution 124, 509–522 (2003).
Hoekstra, P. F. et al. Trophic ecology of bowhead whales (Balaena mysticetus) compared with that of other arctic marine biota as interpreted from carbon-, nitrogen-, and sulfur-isotope signatures. Canadian Journal of Zoology 80, 223–231 (2002).
Muir, D. et al. Bioaccumulation of PCBs and Chlorinated Pesticides in Seals, Fishes and Invertebrates from the White Sea, Russia. Science of The Total Environment, Persistent Organic Pollutants and Heavy Metal Contamination in the Russian Arctic Marine and Freshwater Environment 306, 111–31 (2003).
Hesslein, R. H., Capel, M. J., Fox, D. E. & Halfard, K. A. Stable isotopes of sulphur, carbon, and nitrogen as indicators of trophic level and fish migration in the Lower Mackenzie River Basin, Canada. Canadian Journal of Fisheries and Aquatic Sciences 48, 2258–2265 (1991).
Feige, N., Ehrich, D., Popov, I. Y. & Broekhuizen, S. Monitoring Least Weasels after a Winter Peak of Lemmings in Taimyr: Body Condition, Diet and Habitat Use. Arctic 65, 273–82 (2012).
DeNiro, M. J. & Epstein, S. Influence of Diet on the Distribution of Carbon Isotopes in Animals. Geochimica et Cosmochimica Acta 42, 495–506 (1978).
Spaulding, R. L., Krausman, P. R. & Ballard, W. B. Summer diet of gray wolves, Canis lupus, in northwestern Alaska. Canadian Field-Naturalist 112, 262–266 (1998).
Okarma, H. The trophic ecology of wolves and their predatory role in ungulate communities of forest ecosystems in Europe. Acta Theriologica 40, 335–386 (1995).
Guiry, E. J. Dogs as analogs in stable isotope-based human paleodietary reconstructions: a review and considerations for future use. J Archaeol Method Th 19, 351–376 (2012).
Wiley, D. F. et al. Evolutionary morphing. Proceedings of the IEEE Visualization 2005 (VIS’05), 431–438 (2005).
Bookstein, F. L. Morphometric Tools for Landmark Data: Geometry and Biology. (Cambridge University Press, 1991).
Acknowledgements
We thank C.P. Klingenberg for critical discussion of methodology. A. Drake and R. Losey were supported by a grant from the Social Sciences and Humanities Research Council of Canada grant (#SSHRC IG 435-2014-0075) and a European Research Council Grant to D. Anderson (#295458). M. Sablin acknowledges participation of ZIN RAS (state assignment № АААА-А17-117022810195-3) to this research.
Author information
Authors and Affiliations
Contributions
A.G.D., M.C., P.A.K., O.P.B., M.S., A.V.G., and R.J.L. designed the study. A.G.D., M.C., and R.J.L. wrote the paper. A.G.D., M.C., and L.S.F. performed the analytical work.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Drake, A.G., Coquerelle, M., Kosintsev, P.A. et al. Three-Dimensional Geometric Morphometric Analysis of Fossil Canid Mandibles and Skulls. Sci Rep 7, 9508 (2017). https://doi.org/10.1038/s41598-017-10232-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-10232-1
This article is cited by
-
Possible origins and implications of atypical morphologies and domestication-like traits in wild golden jackals (Canis aureus)
Scientific Reports (2023)
-
Wild or domestic? A 3D approach applied to crania to revisit the identification of mummified canids from ancient Egypt
Archaeological and Anthropological Sciences (2023)
-
Interrelations Between the Cranium, the Mandible and Muscle Architecture in Modern Domestic Dogs
Evolutionary Biology (2020)
-
How Does Masticatory Muscle Architecture Covary with Mandibular Shape in Domestic Dogs?
Evolutionary Biology (2020)
-
Bony labyrinth shape differs distinctively between modern wolves and dogs
Zoomorphology (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
