
Abstract
Below-ground soil microorganisms can modulate above-ground plant-insect interactions. It still needs to be determined whether this is a direct effect of single species or an indirect effect of shifts in soil microbial community assemblages. Evaluation of the soil microbiome as a whole is critical for understanding multi-trophic interactions, including those mediated by volatiles involving plants, herbivorous insects, predators/parasitoids and microorganisms. We implemented a regulated system comprising Nerium oleander plants grown in soil initially containing a sterile/non sterile inoculum, herbivore Aphis nerii and predator Chrysoperla carnea. After aphid attack, plants emitted a characteristic blend of volatiles derived from two biosynthetic classes: fatty acid catabolites and aromatic-derived products. Three aliphatic compounds were mainly detected in plants grown in the inoculated microbial soil, a blend which was preferentially chosen by C. carnea adult females. The contrasting effect of the initial inocula was attributed to the different microbial consortia developed in each treatment. We argue that differences in the relative abundance of the active microbial communities in the rhizosphere correlate with those in the emission of selected volatile compounds by attacked plants. The mechanisms involved in how the functional soil microbiome modulates inducible indirect defence of plants are discussed.
Similar content being viewed by others

Introduction
As part of the evolutionary adaptation process, phytophagous insect-plant interactions occur according to the behavioural choices of insects and the development of physical and chemical plant defences1. In addition, volatile plant secondary metabolites possibly function as signals in communications with other organisms in the environment. In this context, the plant-mediated effects on predator-prey and host-parasitoid interactions in tri-trophic systems of herbivore-induced plant volatiles (HIPVs) have been well-documented2. The release of HIPVs, generally a mixture of green-leaf volatiles, terpenes and aromatic compounds, among others3, may signal the presence of potential prey or hosts and therefore can be exploited by natural enemies to locate the prey organism. Pioneering plant studies show how chewers, sap feeders and herbivore egg deposition induce the production of volatiles attractive to entomophagous arthropods4,5,6,7.
In addition to indirect interactions, plants act as a link between above- and below-ground communities. It is well known that soil-borne non-pathogenic microbes can modulate plant-insect above-ground interactions via plant growth promotion or induced systemic resistance by triggering biochemical changes in the primary plant metabolism8. From a multi-trophic perspective, the tri-trophic role of plant secondary chemistry has been shown to be central to an understanding of various aspects of trophic phenomena, including top-down and bottom-up regulation of herbivores. In recent years, several studies have investigated the effects of below-ground microbes on the third trophic level organisms via changes in HIPV emission9. However, most studies focusing on modifications in plant volatile emission through interactions with soil microorganisms mainly address plant interactions with single species of non-pathogenic microbes, which have a neutral, synergistic or antagonistic effect on plant secondary chemistry10, 11. Given the significant impact of microbial community diversity and richness on plant signalling pathways, the root microbiome as a whole needs to be considered in relation to many aspects of plant immunity12, 13 such as induced indirect defence and insect population dynamics. Nevertheless, to date, no overall trend has emerged in relation to the effects of increased microbial complexity on microbe-plant-insect interactions, with some evidence showing a significant, limited or zero impact on above-ground herbivores14.
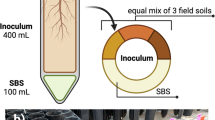
In this study, we aim to evaluate the role of the rhizosphere microbiome in HIPV production and its impact on above-ground community interactions. In a case study, we designed a system-based model with three trophic levels, with Nerium oleander as host plant, Aphis nerii as phloem-sap feeder and Chrysoperla carnea as predator (Fig. 1). The plants were grown for 3 months in sterilized soil with a microbial inoculant and were then infested with A. nerii. The HIPVs produced were sampled using solid-phase micro-extraction (SPME) and were measured by GC-MS. A Y-tube olfactometer was used to investigate the orientational response of C. carnea to the plant volatiles. Using a metagenomic approach, we analyzed the composition and diversity of entire and active microbial communities.

Regulated olfactometer system comprising Nerium oleander plants grown in a potting soil initially containing a sterile (Control)/non sterile (Vermicompost) inoculum, the herbivorous Aphis nerii and the generalist predator Chrysoperla carnea.
Results
Chrysoperla carnea behaviour
Model selection indicated that the most parsimonious model only takes account of fixed factor treatment without considering block and assay variables (AICc = 167.21, ΔAICc = 2, R2 = 0.25). Selected model estimated 2.55 C. carnea individuals chose the branch connected with the sterilised vermicompost treatment, whereas the non-sterilized vermicompost treatment was chosen by a predicted total average of 4.32 C. carnea individuals. This means that 37.12% of individuals chose the Control treatment, while 62.88% selected the Vermicompost treatment (Fig. 2). The results indicate a clear preference of female C. carnea adults for the volatile compounds produced under vermicompost soil treatment conditions.

Percentage of Chrysoperla carnea moving towards plants of Nerium oleander grown in a potting soil initially containing a sterile (Control)/non sterile (Vermicompost) inoculum and after Aphis nerii attack.
Volatiles
The capacity of N. oleander to emit volatiles has been previously described15, 16. However, the volatile blend emitted by N. oleander plants under non-attack conditions was quantitatively insufficient to be detected by SPME-GC in our experiment. Depending on the potting soil media used, N. oleander plants damaged by A. nerii under laboratory conditions emitted a different variety of volatile organic compounds 3 days after infestation (Fig. 3). The 2-decanone, 2-dodecanone and tetradecane compounds were mostly detected when plants were grown in the microbial inoculated soil (Vermicompost treatment).

Volatiles (mean ± SE) emitted by Nerium oleander plants grown in a potting soil initially containing a sterile (Control)/non sterile (Vermicompost) inoculum and after attack by Aphis nerii. V 1 : 4-methoxy-benzaldehyde (P = 0.600, F = 0.323), V 2 : benzothiazole (P = 0.627, F = 0.276), V 3 : benzyl alcohol (P = 0.091, F = 4.902), V 4 : dichlorobenzoic acid (P = 0.090, F = 4.962), V 5 : alkylbenzenes (P = 0.403, F = 0.873), V 6 : 1,2,4-trimethylbenzene (P = 0.831, F = 0.052), V 7 : 2-decanone (P = 0.001, F = 62.381), V 8 : 2-undecanone (P = 0.830, F = 0.053), V 9 : 2-dodecanone (P = 0.001, F = 63.562), V 10 : tetradecane (P = 0.005, F = 33.091).
Nerium oleander dry weight and chemical characteristics of soil samples
After harvest, no differences in the dry weight of N. oleander plants between the two treatments were detected (Control = 62.91 g, Vermicompost = 64.57 g; F = 0.213, P = 0.656). Detailed information on potting soil characteristics can be found as Supplementary Table S1. As expected, fractional sterilization had no significant effect on the intrinsic chemical properties of the vermicompost17. No significant differences were detected between the two treatments, with values for pH, C:N ratio and macro and micronutrients found to be comparable in Control and Vermicompost soils at the end of the experiment.
Composition of active and whole microbial communities
After 3 months of incubation, the two microbial communities (from a sterile and natural vermicompost) showed different taxonomical structures when inoculated in the same sterile soil. We retrieved over 160,000 bacterial and fungal operational taxonomic units (OTUs) from potting soils previously surveyed by pyrosequencing rRNA gene and gene transcript amplicons.
Bacterial communities
Eight dominant phyla (abundance > 1%) were present in the rhizosphere soil samples, accounting for more than 99% of all bacterial sequences (Fig. 4). The total bacterial community was dominated by Firmicutes (Control 79.49%, Vermicompost 46.07%), Actinobacteria (Control 12.14%, Vermicompost 16.52%) and Proteobacteria (Control 4.18%, Vermicompost 20.79%). Its composition differed significantly between treatments, with, most notably, a single phylum, Firmicutes, accounting for almost 80% of the entire bacterial population in the Control treatment. On closer analysis, no particular differences between treatments in terms of genus or species were detected, although relative abundance within bacterial classes did vary across treatments (Supplementary Fig. S1). For example, we observed differences between relative abundances of orders Bifidobacteriales (97% Control, 54% Vermicompost) and Actinomycetales (6% Control, 35% Vermicompost) in Actinobacteria, genera Lactobacillus (94% Control, 76% Vermicompost) and Clostridium (72% Control, 48% Vermicompost) in Bacilli and Clostridia, respectively, and between relative abundances of orders Pseudomonadales (62% Control, 28% Vermicompost) and Xanthomonadales (28% Control, 46% Vermicompost) in Gammaproteobacteria.

Relative abundance of total (16S rRNA gene) and active (16S rRNA transcript) bacteria in potting soil initially containing a sterile (Control)/non sterile (Vermicompost) inoculum.
With respect to the active bacterial community, the differences in relative abundance at class level were less marked between treatments than those detected in the total bacterial community (Fig. 4). The phylum Verrucomicrobia, which was detected at the DNA level with insufficient relative abundance (<1%), was identified at the RNA level with some differences observed in relative abundance at the order level (Supplementary Figs S2, S3). We also detected differences between relative abundances of orders Actinomycetales in Actinobacteria and order Caulobacteriales in Alfaproteobacteria (21% Control, 9% Vermicompost) as examples. The richness indices showed a similar comparative trend in terms of predicting the number of OTUs in all soil samples, displaying those from the Vermicompost treatment the highest alpha diversity -Chao1 and Faith’s Phylogenetic Diversity- values at both the DNA and RNA levels (Supplementary Fig. S4). Beta diversity patterns in the four datasets (Vermicompost DNA, Control DNA, Vermicompost RNA, Control RNA) were examined using qualitative and quantitative similarity indices at the putative species level (Table 1). The Sorensen-Dice distance and Bray-Curtis dissimilarity indices, based on presence/absence and abundance data, respectively, showed a high degree of differentiation among bacterial communities, particularly between total and active populations in treatment Control. At the RNA level, the Bray-Curtis dissimilarity index also displayed a certain degree of similarity between treatments at the RNA level. UniFrac distances, incorporating information on relative relatedness of community members through the inclusion of the phylogenetic distances between organisms, reflected a similar trend. Both the weighted (quantitative) and unweighted (qualitative) variants of UniFrac point to a medium-to-high degree of dissimilarity in bacterial population structure both in and between treatments at the DNA and RNA levels. It is important to note that the number of denoised sequences (62.160 and 49.730 per sample of DNA and RNA, respectively) is considered an enough sequencing depth to describe a trend in both alpha and beta diversity18.
Fungal communities
Three dominant phyla accounting for over 99% of all fungal sequences were present in the rhizosphere soil samples. The number of assigned reads was 152.643, 136.583, 128.412 and 137.465 for Control-DNA, -RNA, and Vermicompost-DNA and -RNA sets, respectively. The total and active fungal communities were dominated by two phyla, Ascomycota and Chytridiomycota, with remarkable differences between the two treatments (Fig. 5). While neither total nor active communities ascribed to Ascomycota changed significantly following each treatment (Control DNA 91.52%, Vermicompost DNA 67.93%; Control RNA 97.75%, Vermicompost RNA 74.07%), Chytridiomycota varied to some extent in terms of relative abundance in treatment Control at the RNA level (Control DNA 7.52%, Vermicompost DNA 30.30%; Control RNA not detected, Vermicompost RNA 22.45%). Fungal species belonging to phylum Basiodiomycota were only detected at the DNA level. In class terms, the main differences between treatments at the DNA level were found with respect to phylum Ascomycota, particularly in classes Sordaromycetes (Control 8.37%, Vermicompost 44.39%) and Pezizomycetes (Control 80.2%, Vermicompost 17.2%). The latter was also the predominant active fungal class in both treatments, particularly in rhizosphere soil Control (relative abundance 92%). Class Chytridiomycetes accounted for all Chytridiomycota fungi at the DNA and RNA level.

Relative abundance of total (18S rRNA gene) and active (18S rRNA transcript) fungi in potting soil initially containing a sterile (Control)/non sterile (Vermicompost) inoculum.
Microbial data analysis
Analysis of similarity (ANOSIM) was used to statistically determine the effects of the initial inoculum on final rhizosphere microbial community structure after 3 months. This analysis generated an R value of 1, indicating total separation between soils at either the DNA (P = 0.098) or RNA (P = 0.094) level. Similarity percentage (SIMPER) analyses show the major peaks corresponding to the differences between the two treatments. With respect to total bacterial and fungal communities (Table 2), the OTUs assigned to 18 soil microbial classes accounted for 90% of total dissimilarity (Bray-Curtis dissimilarity = 26.5%). The members of classes Deltaproteobacteria, Betaproteobacteria and Acidobacteria accounted for roughly 25% of community variation between the two soils. With regard to active microbial communities (Bray-Curtis dissimilarity = 21.7%), 11 soil microbial classes accounted for 90% of total variation across different treatments, with Pedosphaerae (Verrucomicrobiota) and Acidobacteria, contributing to 25% of the dissimilarity between the two groups, as the most discriminant species (Table 3). To determine whether community composition affects volatile blend composition, canonical correspondence analysis (CCA) was performed. To test our hypothesis in this multitrophic system, volatiles were regarded as dependent analytical factors. CCA analysis carried out to examine the effect of total and active community composition on volatile emission resulted in a species-volatiles correlation of 0.999 and significant axes in both cases (trace = 0.234, F = 60.185, P = 0.0020), with CCA axis 1and axis 2 accounting for over 98% of variance. The results show two distinct groups corresponding to the two soils, each positively correlated with specific microbial classes at either DNA or RNA level (Fig. 6).

CCA triplot analysis of volatiles emitted by Nerium oleander plants grown in a potting soil initially containing a sterile (C: Control)/non sterile (VC: Vermicompost) inoculum and after attack by Aphis nerii. Arrows indicate the orientations of total (a) and active (b) rhizosphere microbiome. V 1 : 4-methoxy-benzaldehyde, V 2 : benzothiazole, V 3 : benzyl alcohol, V 4 : dichlorobenzoic acid, V 5 : alkylbenzenes, V 6 : 1,2,4-trimethylbenzene, V 7 : 2-decanone, V 8 : 2-undecanone, V 9 : 2-dodecanone, V 10 : tetradecane.
Discussion
This study is based on the “integration” approach which uses previous evidence concerning the role of the below-ground rhizosphere microbiome at above-ground third trophic level. We tested N. oleander plants grown in a potting soil initially containing a sterile/non-sterile olive waste vermicompost as inoculum. When attacked by A. nerii after 3 months of growth, SPME fiber analysis revealed the presence in both treatments of a characteristic blend of plant volatiles derived from two biosynthetic classes: fatty acid catabolites and aromatic-derived products. The composition of the aromatic blend was, in qualitative and quantitative terms, generally similar in both treatments, with comparable relative amounts of 4-methoxy-benzaldehyde, benzothiazole, benzyl alcohol, dichlorobenzoic acid, alkylbenzenes and 1,2,4-trimethylbenzene (Supplementary Fig. S5). Many volatile compounds containing an aromatic ring produced by the shikimate pathway have been described in a wide range of plant species regarded as HIPVs. For instance, 4-methoxy-benzaldehyde has been described in herbivore-infested Arabidopsis thaliana plants19; in aphid species, perception of this plant-specific volatile component assists olfactory discrimination between host and non-host plants20. The release of insect-induced benzothiazole in rice and sunflower plants damaged by Tibraca limbativentris and Euschistus heros has also been described21. Benzyl alcohol has been identified in Camellia sinensis and Coffea canephora under attack from different insect feeding guilds22 and even under mechanical damage conditions23. Dichlorobenzoic acid has been described as a functional analogue of the defence hormone salicylic acid, which acts as a bioactive plant defence-inducing compound24. Many different alkyl benzenes have also been retrieved from infested plants, especially in the presence of high densities of fungi25. Finally, for example, 1,2,4-trimethyl benzene has been characterized as a Brassica oleracea HIPV when infested with Pieris rapae larvae26. The volatile blends of fatty acid derivatives contained four aliphatic compounds: the methyl ketones 2-decanone, 2-undecanone and 2-dodecanone, as well as the alkane tetradecane, all low-molecular-weight substances previously classified as HIPVs. The results obtained by Lozano et al.27 indicate that 2-decanone is involved in attracting the parasitoids Dendrosoter protuberans and Cheiropachus quadrum to their host Phloeotribus scarabaeoides. Some studies have revealed the role played by 2-undecanone in an integrated pest management strategy for tomato28 and solanaceous crops, the latter due to its capacity to alter Bactericera cockerelli behavior29. However, other studies have also shown that 2-undecanone emissions do not differ between Brassica rapa plants exposed/not exposed by Pieris brassicae to leaf herbivory30. Hervibore-induced 2-dodecanone has been detected as a tomato volatile blend component31. The alkane tetradecane, a known semiochemical for many arthropods32, has also been identified as a volatile biomarker indicating damaged flower head tissue33. The most remarkable finding of our study was that three aliphatic compounds were mainly detected after an aphid attack in plants grown in the inoculated microbial soil. Neither 2-decanone nor 2-dodecanone was detected in any of the plants of the Control treatment, at least above the detection limit of the method and in the experimental conditions described above. The other key volatile, tetradecane, was emitted by all the infected plants, either grown in Control or Vermicompost conditions, being the emission significantly higher in the last.
The biosynthesis and regulation of plant volatiles have been widely studied34. Herbivore damage usually elicits phytohormone-mediated changes in the expression of genes involved in biosynthetic pathways for the production of plant volatiles. Variables influencing plant volatile emissions are multiple, including abiotic and biotic characteristics such as plant species, variety, phenology, physiology and nutritional quality, environmental conditions such as light, temperature and moisture status, herbivore density and population growth, as well as biotic and abiotic soil stresses10. In our experiment, carried out under controlled environmental conditions, any possible impact of these stresses was limited. Plants of the same variety were grown under similar environmental conditions and under the same physico-chemical soil characteristics and no differences in plant dry biomass were detected between the two treatments. Also, even though the pre-reproductive period of a mature A. nerii can be less than 2 days35, no visual differences in aphid population size among treatments were observed similarly to that noted by other authors in comparable experiments36, notwithstanding that aphid number was not actually quantified.
Under all the standardized conditions, the contrasting effect of the initial inocula on shifts in the emission of the fatty acid-derived volatile blends could be mainly attributed to the different microbial consortia developed in each treatment. In this context, the main cause of the synthesis of fatty acid-derived volatiles has been identified as the lipoxygenase pathway which is positively regulated by phytohormones such as jasmonic acid37. In general, although phloem feeders activate the salicylic acid-dependent shikimic acid pathway, the important role played by jasmonic acid in defence against aphids has also been demonstrated38. Globally, phytohormone crosstalk has been shown to be considerably involved in the biosynthesis of plant volatiles, although the mechanism by which the microbe interacts with the plant, which subsequently affects plant-insect interaction, has not been fully elucidated39. Beneficial micro-organisms such as mycorrizae and rhizobacteria have been demonstrated to play a role in modulating plant-induced systemic resistance, which is mediated by phytohormone signaling40, 41. However, studies focusing on the role of soil microorganisms in the modification of plant volatile emission are still scarce. Previous findings have shown that non-pathogenic root-associated microbes can have a positive or a negative effect on the attraction of third trophic level organisms through changes in the composition of the blend of herbivore-induced plant volatiles14, 42. Despite the necessity for further research in order to elucidate the mechanisms underlying these contrasting effects, the role assigned to microorganisms involves changes in defence-related signaling. Given that stress responses share signalling pathways regulated by defence-related phytohormones43, soil organisms could have the capacity to modulate the synthesis of fatty acid-derived volatiles. From a multitrophic perspective, the differences in the emission of selected aliphatic compounds by plants under aphid attack initiate a volatile blend attractive to a generalist predator. Thus, C. carnea adult females exhibit some degree of preference for certain volatile blends. It is well known that volatile plant secondary metabolites such as terpenes and aromatic compounds can be detected by the olfactory system of C. carnea adults to locate suitable hosts44, 45. Although these adults are not predatory, some research evidences mechanisms to attract females and concentrated them locally in the field to increase egg-laying intensity, to get advantages of the potential of C. carnea larvae as biological control agents46. It is possible to infer from our study that C. carnea adult females respond to a volatile blend, in which the presence of aliphatic compounds is a determining factor.
However, most studies focusing on below- and above-ground interactions involve single microbes, which differ greatly from natural soil conditions, highlight the importance of evaluating the entire soil microbiome when studying microbe-plant-insect interactions. Little research has been devoted to the impact of the soil microbiome on primary metabolite production in plants, which, in turn, determines insect feeding behavior8. It is still important to determine whether the soil microbiome modulates the biosynthesis of secondary metabolites involved in plant-insect interactions. In this study, we aimed to assess whether alterations in the microbial community composition of a sterile soil following vermicompost amendment modify generalist predator responses to shifts in volatile compound emission induced by a phytophagous attack. The effects of the vermicompost-borne microbial community on both the entire and functional rhizospere microbiome structure were evident after three months. Microbiome composition structurally evolved in line with the composition of the inoculums, while the chemical characteristics of the potting soil remained unchanged. Canonical Correspondence Analysis was used to determine whether community composition affects volatile blend composition. The results show two distinct groups corresponding to the two soils, each positively correlated with specific microbial classes at either the DNA or RNA level. It was then necessary to determine whether the plant-mediated effects of microbes on above-ground herbivores are dependent on microbial community composition and the role, if any, played by the microbial physiological state. The soil samples and volatiles matched different microbial classes depending on the total or active nature of the microbial population. As mentioned above, communication is well known to occur between plants and microorganisms, in which their signalling molecules play an important role. Active microorganisms may then be critically involved in this molecular dialogue, whose precise mechanisms in herbivore-induced plant volatile emission still need to be determined. In this study, Vermicompost treatment samples were clustered with the aliphatic volatiles 2-decanone, 2-dodecanone and tretadecane, and positively correlated with active fungi ascribed to classes Chytridiomycetes and Agaricomycetes and active bacteria of classes Saprospirae, Deltaproteobacteria, Actinobacteria, Acidobacteria, Bacilli and Clostridia. Previous studies of the relationship between microbiome members and plant volatile induction by herbivory have described the effects of soil bacteria ascribed to classes Alpha- and Gamma-proteobacteria, as well as endophytic fungi, but not those of the taxa mentioned above42, 47, 48, although it should be noted that these studies focus on the role of single species. However, from a metagenomics perspective, we did not find any remarkable differences in the structure of Alpha- and Gamma-proteobacteria bacterial communities, with virtually all the bacterial species ascribed to these classes sharing the soils of the two treatments (Supplementary Fig. S3).
Under our experimental conditions, we found some evidence to show the contribution of a group of functional rhizosphere bacteria and fungi to the diversity of plant volatile patterns. Given the absence of previous data on the role of free-living classes of fungi in indirect induced plant defence, we focused on a more in-depth analysis of the bacterial population. In this regard, we were unable to identify any differences in bacterial patterns between Vermicompost and Control rhizospheres, with both soils appearing to share virtually all bacterial species ascribed to the discriminant taxa, particularly at the RNA level (Supplementary Figs S2, S3). Although some authors have debated the key role played by rare species within the entire microbiome in plant-herbivore interactions35, we cannot attribute any critical function to species ascribed to those classes identified as discriminant between treatments. On the contrary, using a metagenomic approach, we showed that the principal difference mainly related to the relative abundance of functional microorganisms. In this regard, the estimators of alpha diversity49 indicated higher within-community diversity in the Vermicompost rhizosphere with respect to that found in the Control treatment. The Chao 1 index50 and the Faith’s Phylogenetic Diversity51, based upon the number of rare OTUs and expressing the number of tree units found in a sample, respectively, confirmed that phylogenetic and functional structures of the bacterial communities differed between treatments. In addition, the beta diversity patterns, especially those including the phylogenetic distances between organisms, pointed to a medium-to-high degree of dissimilarity between the bacterial population structure in Vermicompost and Control rhizospheres.
Concluding Remarks
In this study, the effects of below-ground microbes on indirect plant defences were evaluated. We aimed to determine whether alterations in the microbial community composition of a sterile soil following vermicompost amendment modify plant-insect interactions through shifts in volatile compound emission. We found that differences in the composition of the rhizosphere active microbiome correlate with those in the emission of selected aliphatic compounds by plants under aphid attack, which initiates a volatile blend attractive to a generalist predator. Although further research is required, our results suggest that functional interactions between soil microbes play a significant role in regulating the biosynthesis of volatile plant secondary metabolites. The functional soil microbiome is a factor which needs to be investigated in order to assess whether the plant-mediated effects of soil microorganisms on above-ground herbivores are species- or population structure-dependent.
Materials and Methods
Plants
The plant species used in the experiments was 3-month-old oleander (Nerium oleander L. Apocynaceae) grown in a greenhouse nursery (25–30 °C, 60–80% RH, 16:8 h L:D). The roots were washed by being dipped in sterile water to remove soil and were then individually transplanted to a pot filled with the potting soil mixture. At harvest (90 days after planting), the plants were removed and separated into shoots and roots. The shoots were oven-dried at 60 °C for 48 h and their mass determined on an analytical balance.
Potting soil
A mixture of sand and loamy clay soil (1:1 v:v), previously sterilized by fractional sterilization (tyndallization; 100 °C, 60 min, 3 days), was used as potting medium. The soil was a calcareous loam (Typic Xerorthent)52 collected from an agricultural field (0–20 cm in depth) in Granada, Spain. Tyndallization involves killing vegetative cells and some spores at the initial heating stage; additional heat resistant spores germinate and are killed at a later heating stage. This low temperature sterilization technique preserves soil structure and quality more effectively than autoclaving at 121 °C53. The tyndallized soil characteristics were as follows: 0.9 g kg−1 SOC (soil organic carbon), 1.6 g kg−1 total N, pH (H2O) 7.5.
We used a vermicompost from olive-mill waste produced at the EEZ-CSIC facility (Granada, Spain), as described in Vivas et al.54, as microbial inoculum. In order to attain a soil organic carbon content of 30 g kg−1, 1,000 g of the soil mixture was placed in 2-l black pots and thoroughly mixed with the vermicompost at a rate corresponding to 50 g kg−1 (Vermicompost treatment). Soil amended with the same amount of vermicompost sterilized by tyndallization was used as control (Control treatment). Soil moisture content was adjusted to approximately 60% and maintained at this level by irrigation with sterilized deionized water during the experiment. Three replicates per treatment were arranged in randomized blocks in the greenhouse (25 °C, 60–80% RH, 16:8 h L:D).
Predators
The Chrysoperla carnea Steph. (Neuroptera: Chrysopidae) larvae were supplied by Koppert Biological Systems (La Mojonera, Almería, Spain). Larvae were individually reared in Petri dishes and fed on Ephestia kuehniella Zell. (Lepidoptera: Pyralidae) eggs. Upon emergence, C. carnea adults were collected daily and kept in boxes (28 cm diameter, 15 cm high) with an ovipositing surface; they were then fed on honey:pollen (1:1, v-v) and mineral water and maintained in a controlled environment cabinet at 25 °C, 50–60% RH and 16:8 h L:D for 2–3 days. Adult C. carnea were sexed by examining the ventral abdominal tip surface. Only females were used for bioassays.
Phytophagous insects
Aphis nerii Boy. (Homoptera: Aphididae) adults were taken from 20-y-old N. oleander plants located in Gójar, Granada, Spain. 20 individuals were reared on 1-y-old oleander plants maintained in a chamber at 25 °C, 50–60% RH and 16:8 h L:D. The plants were covered with fine mesh netting to prevent A. nerii emigration. When the plants were badly damaged by aphids, they were replaced by fresh plants after aphid migration to the healthy plants. Approximately 7 generations of aphids were produced before being used for the experiments.
Experimental design
A closed-system Y-tube olfactometer (ID 3 cm; stem 10 cm, arms 8 cm; stem-arm angle 130°) was used to assess choice of predator C. carnea between the two treatments after a 3-month N. oleander growing period (Fig. 1). Two glass chambers (40 × 40 × 140 cm), sufficiently large to accommodate aboveground plant tissues were connected from the top to the Y-shaped glass tubing of the olfactometer by transparent polytetrafluoroethylene. An SPME fiber was inserted into each arm of the olfactometer in order to collect volatiles. Using air pressure, synthetic pure air, at an airflow rate of 1.2 l min−1 per channel55, was drawn in though the bottom of the chambers.
Firstly, uninfested plants were tested in the chambers connected to the olfactometer, and the volatiles were recovered. To infest N. oleander, 20 wingless A. nerii adults were introduced at the top of the plant. After inoculation, the plants were re-introduced into the glass chambers. The aphids were allowed to feed for 48 h, and tests were conducted three days after inoculation.
Behavioural tests of predator C. carnea were carried out under artificial light between 09:00 and 18:00 h at 28 ± 2 °C. A white circular paperboard arena was placed around the olfactometer to prevent visual disturbances. Adult C. carnea females were inserted into the single branch of the olfactometer and were left to choose between the two branches of the device, with a maximum observation period of 5 min. If the insects, which were used only once and then discarded, did not attain a length of at least 4 cm along the arm connected to the test chambers, they were excluded from the data analysis. To rule out directional bias, the olfactometer was washed in hot water, rinsed in sterilized deionized water and dried in an oven at 60 °C before each experiment. The Y-tube was also rotated 180° after each test. The position of the chambers was changed after every two tests. The pairwise experiment on the behaviour of two plants was repeated three times every other day. In each assay, approximately 7 C. carnea females were used. A total of 22 behavioural assays were carried out with C. carnea females from three different rearing groups (blocks).
Volatiles
The volatiles emitted by N. oleander were sampled using SPME on day 3 for 9 h from 09:00 to 18:00 h. The SPME fibers (50/30 μm DVB/CAR/PDMS Stableflex 23Ga, Autosampler, 3pk, SUPELCO, Bellefonte, PA, USA) were preconditioned prior to analysis at 250 °C for 30 min. After the equilibration period, the fibers were exposed to the headspace of each Y-tube olfactometer arm. After completion of sampling, the fiber was withdrawn into the needle and inserted into the GC–MS system injection port in splitless injection mode at an injector temperature of 250 °C.
Gas chromatography (GC) analyses were conducted on a Varian 450-GC gas chromatograph fitted with a 1079 injector in split/splitless mode, a CTC Analytics CombiPal refrigerated autosampler and a Varian 240 Ion Trap as a mass spectrometer detector. A FactorFour VF-5ms (30 m × 0.25 mm × 0.25 μm) fused silica capillary column was also used.
The initial gas chromatography oven temperature was 50 °C for 5 min which was then increased to 260 °C at 10 °C min−1. It was then raised to a temperature of 300 °C at 30 °C min−1, which was maintained for 1 min with an injection of 1 μl (300 °C) in splitless mode (1 min). The carrier gas was He at 1 ml min−1. Electron impact ionization and detection in full scan (m/z 40 to 450) modes were carried out. The transfer line and trap temperatures were 290 and 210 °C, respectively. Peaks were identified by comparing the volatile sample mass spectra with spectra in the NIST08 Mass Spectral Database (MS Workstation 6.9.1. software). When necessary, the retention index (RI) was calculated for each volatile compound using the retention times of a homologous series of n-alkanes and by comparing the RI with that of compounds analyzed under similar conditions in the literature to confirm the identity of volatile compounds.
Rhizosphere soil samples
Rhizosphere soil was collected in two steps. First, the root system was separated from the bulk soil by gentle shaking, and the remaining soil was then removed from the roots by more vigorous shaking. Soil still adhering to the roots was removed using a sterile dissecting probe and collected for use as rhizosphere soil. Root-associated soil samples from each pot were placed in separate polyethylene bags and immediately stored at −80 °C for subsequent molecular analyses.
16S/18S rRNA gene sequence analysis
For each rhizosphere soil sample replicate, total DNA was separately extracted from four 1 g subsamples using the bead-beating method following the manufacturer’s instructions for the MoBio UltraClean Soil DNA Isolation kit (MoBio Laboratories, Solana Beach, CA, USA). The extracts were pooled and further concentrated at 35 °C to a final volume of 20 μl, with the aid of a Savant Speedvac® concentrator. Total RNA was extracted from four 2 g subsamples of each replicate according to the manufacturer’s instructions for the MoBio RNA PowerSoil Total RNA Isolation kit (MoBio laboratories, Solana Beach, CA, USA). To remove residual DNA, the DNase I enzyme was added using the Roche RNase-Free DNase set (Roche Applied Science, Penzberg, Germany) according to the manufacturer’s instructions. The extracts were pooled and further concentrated at 35 °C to a final volume of 80 μl with the aid of a Savant Speedvac® concentrator. The cDNA was synthesized from 1–2 μg of total RNA-DNase using the Transcriptor High Fidelity cDNA Synthesis Kit according to the manufacturer’s instructions (Roche Applied Science, Penzberg, Germany). The synthesis reaction was carried out at 50 °C for 30 min. The concentration and quality of the final DNA/RNA/cDNA samples were checked by a Nanodrop® ND-100 spectrometer (Nanodrop Technologies, Wilmington, DE, USA).
The metagenome and transcriptome libraries of the bacterial 16S rRNA gene were generated in pooled soil samples using the primers S-D-Bact-0341-b-S-17/S-D-Bact-0785-a-A-21 reported by Klindworth et al.56 (non-underlined sequences) and were fused with underlined Illumina adapter overhang nucleotide sequences. To amplify V3-V4 hypervariable regions of the 16S rRNA gene, the following primer sequences were used: 5′-TCGTCGGCAGCGTCAGATGTGTATAAGAGACAGCCTACGGGNGGCWGCAG-3′ and 5′ GTCTCGTGGGCTCGGAGATGTGTATAAGAGACAGGACTACHVGGGTATCTAATCC-3′. The amplified region was approximately 464 bp. For each library, triplicate soil PCR products with unique indexes were mixed in equal nanogram quantities and sequenced on the Illumina MiSeq platform using a 2 × 250 nucleotide paired-end protocol (Era7 Bioinformatics, Granada, Spain).
Metagenomic and transcriptomic analyses of the fungal 18S rRNA gene were performed on the degenerate primers 563 f (5′-GCCAGCAVCYGCGGTAAY-3′) and 1132r (5′-CCGTCAATTHCTTYAART-3′) designed by Hugerth et al.57. To prepare libraries for Illumina sequencing, primers 563 f and 1132r were fused with the Illumina adapter overhang nucleotide sequences. The primers were used to amplify the V4 region of the 18S rRNA gene, with the amplicon expected to measure approximately 569 bp. For each library, triplicate soil PCR products with unique indexes were mixed in equal nanogram quantities and sequenced on the Illumina MiSeq platform using a 2 × 300 nucleotide paired-end protocol (Era7 Bioinformatics, Granada, Spain).
The resulting sequences from 16S and 18S rRNA gene libraries were assessed and filtered according to base quality using the FASQC tool (http://www.bioinformatics.babraham.ac.uk/projects/fastqc/). The quality-filtered Illumina paired-end reads were merged using FLASH58. The non-merged reads considered to be invalid for analysis were discarded. The successfully merged fragments were assigned to a taxonomic tree node based on sequence similarity to 16S and 18S rRNA genes extracted from the RNA central database (http://rnacentral.org/) which includes rRNAs from a wide range of major databases such as SILVA, GreenGenes, RDP, RefSeq and ENA. The NCBI taxonomy was used; for taxonomic assignment, we used the MG7 method59 which is based on exhaustive BLAST comparison of each read, covering almost the whole length of the sequence, against the 16S and 18S ribosomal RNA database. This method discarded chimeric sequences as no-hit reads, given that the reference database only contains 16S and 18S sequences and chimeric sequences have a BLAST no-hit result. In addition to chimeric sequences, the majority of no-hits detected in all the samples correspond to phiX reads that are used as controls in the Illumina library preparation protocol in the sequencing experiment. The bacteria detected in the samples were identified on the basis of the BLAST results. Taxonomic assignment was carried out using the BBH (best BLAST hit) and LCA (Lowest Common Ancestor) assignment paradigms. Taxonomic profiling was studied at the sample level60. An additional filtering step was carried out exclusively for the 18S rRNA dataset, with only reads belonging to the Kingdom Fungi being kept for downstream analyses.
All original Illumina sequence data were deposited in the Sequence Read Archive (SRA) service of the European Bioinformatics Institute (EBI) database (BioProject ID: PRJNA313153, accession numbers SRX1795432, SRX1795541, SRX1798892, SRX1798901, SRX1795397, SRX1795517, SRX1798890, SRX1798895).
Chemical analyses
Air-dried rhizosphere soil samples were used to determinate chemical properties. Total N and SOC were determined with the aid of the Leco-TruSpec CN elemental analyzer (LECO Corp., St Joseph, MI, USA). Total mineral content was determined by the digestion method with HNO3 65%:HCl 35% (1:3; v-v) followed by analysis using inductively coupled plasma optical emission spectrometry (ICP-OES) (ICP 720-ES, Agilent, Santa Clara, USA).
Data analysis
The results of the chemical and volatile analyses were the means of 3 replicates. The data were subjected to factorial analysis of variance (ANOVA) using PAST (Paleontological Statistics) software program v3.1461.
Data on the behaviour of C. carnea were analysed to account for differences between treatments by using generalized linear mixed models (GLMMs). A Poisson error structure and log-link function were used to build these models, with the response variable being the count of C. carnea females located at the end of each Y-tube olfactometer branch62. We generated a set of models composed of different combinations of the fixed “treatment” and “block” factors and the random “assay” factor. By their nature, blocks should be regarded as a random factor; however, because this variable contains only three levels, a fixed factor is recommended63. We also tested a set of models with the block variable treated as a random factor and obtained the same results. The most complex of the eight plausible models we constructed, containing all possible combinations of the variables mentioned above, was the following:
where \(\alpha \) represents the intercept of the model; \({\beta }_{1}\) is the estimated value of the treatment effect; \({\beta }_{2}\) is the estimated value of the block effect; and \({\varepsilon }_{{assay}}\) is the estimated error associated with the assays carried out. Alternative models were compared using the Akaike Information Criterion (AICc) corrected for small sample size64. Models showing a difference in AICc > 2 indicate that the worst model has virtually no support and can be ruled out. The selected model was tested to account for unsuited error structure approach with DHARMa package ver. 0.1.3 written for the R environment65, given that the error structure chosen was appropriate for this type of analysis.
An additional analysis of bacterial sequences was carried out using QIIME v1.9.166. The raw files from Illumina paired-end sequencing (R1 and R2) were merged. Quality filtering was then performed using Phred67, 68, with a Phred quality score of Q20. The FASTA files obtained were brought together in a single file. With the aid of UCLUST69, an OTU clustering procedure was performed with a 97% similarity threshold. To facilitate further analysis, a representative set of sequences was selected. An OTU table in biom format70 was obtained to further analyse alfa and beta diversity using different metrics. For the phylogenetic analysis of the representative set of sequences, an alignment using PyNAST71 was carried out, the highly variable regions of sequences were removed and, finally, the phylogenetic tree was obtained with the aid of FastTree72. The phylogenetic tree in newick format is necessary to calculate alfa diversity using the PD_whole_tree metric (Faith’s Phylogenetic Diversity) and beta diversity using the unweighted_unifrac and weighted_unifrac methods.
Analysis of similarity (ANOSIM) and similarity percentage (SIMPER) analyses were performed on total and active microbial communities (OTUs, 16S and 18S ribosomal amplicon pyrosequencing) using PAST software v3.14. Distance indices were calculated with the aid of the Bray-Curtis method. Statistical significance was computed by permutation of group membership with 9,999 replicates. ANOSIM generated an R value, whose magnitude indicates the degree of separation between groups, with a score of 1 indicating total separation and 0 no separation.
The relationship between plant volatile blends and rhizosphere microbiome composition (OTUs, 16S and 18S ribosomal amplicon pyrosequencing) was determined by canonical correspondence analysis in CANOCO 4.573. OTU patterns were fitted to volatile blends by using the Monte Carlo permutation test (499 permutations) at a 95% confidence level. Community similarities were graphed by using an ordination triplot technique with scaling focused on inter-sample differences.
References
Karban, R. The ecology and evolution of induced resistance against herbivores. Functional Ecology 25, 339–347, doi:10.1111/j.1365-2435.2010.01789.x (2011).
Pichersky, E., Noel, J. P. & Dudareva, N. Biosynthesis of plant volatiles: nature’s diversity and ingenuity. Science 311, 808–811 (2006).
Dicke, M. Plant phenotypic plasticity in the phytobiome: a volatile issue. Current Opinion in Plant Biology 32, 17–23, doi:10.1016/j.pbi.2016.05.004 (2016).
Dicke, M. Volatile spider-mite pheromone and host-plant kairomone, involved in spaced-out gregariousness in the spider mite Tetranychus urticae. Physiological Entomology 11, 251–262 (1986).
Turlings, T. C. J., Tumlinson, J. H. & Lewis, W. J. Exploitation of herbivore-induced plant odors by host-seeking parasitic wasps. Science 250, 1251–1253 (1990).
Potting, R. P. J., Vet, L. E. M. & Dicke, M. Host microhabitat location by stem-borer parasitoid Cotesia flavipes: the role of herbivore volatiles and locally and systemically induced plant volatiles. Journal of Chemical Ecology 21, 525–539 (1995).
Tamiru, A. et al. Maize landraces recruit egg and larval parasitoids in response to egg deposition by a herbivore. Ecology Letters 14, 1075–1083 (2011).
Badri, D. V., Zolla, G., Bakker, M. G., Manter, D. K. & Vivanco, J. M. Potential impact of soil microbiomes on the leaf metabolome and on herbivore feeding behavior. New Phytologist 198, 264–273, doi:10.1111/nph.12124 (2013).
Pineda, A., Soler, R., Pozo, M. J., Rasmann, S. & Turlings, T. C. J. Editorial: Above-belowground interactions involving plants, microbes and insects. Frontiers in Plant Science 6, 318, doi:10.3389/fpls.2015.00318 (2015).
Gols, R. Direct and indirect chemical defences against insects in a multitrophic framework. Plant, Cell and Environment 37, 1741–1752 (2014).
Gadhave, K. R., Hourston, J. E. & Gange, A. C. Developing Soil Microbial Inoculants for Pest Management: Can One Have Too Much of a Good Thing? Journal of Chemical Ecology 42, 348–356 (2016).
Mendes, R. et al. Deciphering the Rhizosphere Microbiome for Disease-Suppressive Bacteria. Science 332, 1097–1100 (2011).
Berendsen, R. L., Pieterse, C. M. J. & Bakker, P. A. H. M. The rhizosphere microbiome and plant health. Trends in Plant Science 17, 478–486 (2012).
Pineda, A., Zheng, S. J., van Loon, J. J. A., Pieterse, C. M. J. & Dicke, M. Helping plants to deal with insects: the role of beneficial soil-borne microbes. Trends in Plant Science 15, 507–514 (2010).
Owen, S. M., Boissard, C. & Hewitt, C. N. Volatile organic compounds (VOCs) emitted from 40 Mediterranean plant species: VOC speciation and extrapolation to habitat scale. Atmospheric Environment 35, 5393–5409 (2001).
Holopainen, J. K. & Blande, J. D. Where do herbivore-induced plant volatiles go? Frontiers in Plant Science 4, 185, doi:10.3389/fpls.2013.00185 (2013).
Trevors, J. Sterilization and inhibition of microbial activity in soil. Journal of Microbiological Methods 26, 53–59 (1996).
Lundin, D. et al. Which sequencing depth is sufficient to describe patterns in bacterial α- and β-diversity? Environmental Microbiology Reports 4, 367–372 (2012).
Lin, Y. et al. Volatiles from plants induced by multiple aphid attacks promote conidial performance of Lecanicillium lecanii. PLoS One 11(3), e0151844, doi:10.1371/journal.pone.0151844 (2016).
Visser, J. H. Host odour perception in phytophagous insects. Annual Review of Entomology 31, 121–144 (1986).
Melo Machado, R. C., Sant’Ana, J., Blassioli-Moraes, M. C., Laumann, R. A. & Borges, M. Herbivory-induced plant volatiles from Oryza sativa and their influence on chemotaxis behaviour of Tibraca limbativentris stal. (Hemiptera: Pentatomidae) and egg parasitoids. Bulletin of Entomological Research 104, 347–356 (2014).
Cai, X. M., Sun, X. L., Dong, W. X., Wang, G. C. & Chen, Z. M. Herbivore species, infestation time, and herbivore density affect induced volatiles in tea plants. Chemoecology 24, 1, doi:10.1007/s00049-013-0141-2 (2014).
Cruz-López, L., Díaz-Díaz, B. & Rojas, J. C. Coffee volatiles induced after mechanical injury and beetle herbivory attract the coffee berry borer and two of its parasitoids. Arthropod-Plant Interactions 10, 151, doi:10.1007/s11829-016-9417-0 (2016).
Bektas, Y. & Eulgem, T. Synthetic plant defense elicitors. Frontiers in Plant Science 5: Article 804, doi:10.3389/fpls.2014.00804 (2015).
Cotes, B. et al. Habitat selection of a parasitoid mediated by volatiles informing on host and intraguild predator densities. Oecologia 79, 151–162, doi:10.1007/s00442-015-3326-2 (2015).
Poelman, E. H. et al. Field parasitism rates of caterpillars on Brassica oleracea plants are reliably predicted by differential attraction of Cotesia parasitoids. Functional Ecology 23, 951–962 (2009).
Lozano, C. et al. Response of parasitoids dendrosoter protuberans and Cheiropachus quadrum to attractants of Phloeotribus scarabaeoides in an olfactometer. Journal of Chemical Ecology 26, 791–799 (2010).
Farrar, R. R., Kennedy, G. & Kashyap, R. K. Influence of life history differences of two tachinid parasitoids of Helicoverpa zea (Boddie) (Lepidoptera: Noctuidae) on their interactions with glandular trichome/methyl ketone-based insect resistance in tomato. Journal of Chemical Ecology 18, 499–514 (1992).
Davidson, M. M. et al. A volatile compound, 2-undecanone, increases walking, but not flying, tomato potato psyllid movement toward an odour source. New Zealand Plant Protection 67, 184–190 (2014).
Pierre, P. S. et al. Differences in volatile profiles of turnip plants subjected to single and dual herbivory above- and belowground. Journal of Chemical Ecology 37, 368–377 (2011).
Raghava, T., Ravikumar, P., Hegde, R. & Kush, A. Spatial and temporal volatile organic compound response of select tomato cultivars to herbivory and mechanical injury. Plant Science 179, 520–526 (2010).
El-Sayed, A. M. The Pherobase: Database of Pheromones and Semiochemicals http://www.pherobase.com (2016).
Beck, J. J. et al. In-field volatile analysis employing a hand-held portable GC–MS: emission profiles differentiate damaged and undamaged yellow Starthistle flower heads. Phytochemical Analysis 26, 395–403, doi:10.1002/pca.2573 (2015).
Dudareva, N., Klempien, A., Muhlemann, J. K. & Kaplan, I. Biosynthesis, function and metabolic engineering of plant volatile organic compounds. New Phytologist 198, 16–32 (2013).
Murugan, K., Jeyabalan, D., Kumar, N. S., Nathan, S. S. & Sivaramakrishnan, S. Influence of host plant on growth and reproduction of Aphis nerii and feeding and prey utilization of its predator Menochilus sexmaculatus. Indian Journal of Experimental Biology 38, 598–603 (2000).
Hol, W. H. G. et al. Reduction of rare soil microbes modifies plant-herbivore interactions. Ecology Letters 13, 292–301 (2010).
Rouster, J., Leah, R., Mundy, J. & Cameron-Mills, V. Identification of a methyl jasmonate-responsive region in the promoter of a lipoxygenase 1 gene expressed in barley grain. The Plant Journal 11, 513–523, doi:10.1046/j.1365-313X.1997.11030513.x (1997).
Clavijo McCormick, A. Can plant–natural enemy communication withstand disruption by biotic and abiotic factors? Ecology and Evolution 6, 8569–8582, doi:10.1002/ece3.2567 (2016).
Pangesti, N., Pineda, A., Pieterse, C. M. J., Dicke, M. & van Loon, J. J. A. Two-way plant mediated interactions between root-associated microbes and insects: from ecology to mechanisms. Frontiers in Plant Science 4, 414, doi:10.3389/fpls.2013.00414 (2013).
Sabine, C., Jung, S. C., Martinez-Medina, A., Lopez-Raez, J. A. & Pozo, M. J. Mycorrhiza-Induced Resistance and Priming of Plant Defenses. Journal of Chemical Ecology 38, 651–664 (2012).
Pineda, A., Zheng, S.-J., van Loon, J. J. A. & Dicke, M. Rhizobacteria modify plant–aphid interactions: a case of induced systemic susceptibility. Plant Biology 14, 83–90 (2012).
Pineda, A. et al. Non-pathogenic rhizobacteria interfere with the attraction of parasitoids to aphid-induced plant volatiles via jasmonic acid signalling. Plant, Cell & Environment 36, 393–404, doi:10.1111/j.1365-3040.2012.02581.x (2013).
Pineda, A., Dicke, M., Pieterse, C. M. J. & Pozo, M. J. Beneficial microbes in a changing environment: are they always helping plants to deal with insects? Functional Ecology 27, 574–586, doi:10.1111/1365-2435.12050 (2013).
Flint, H. M., Salter, S. S. & Walters, S. Caryophyllene: an attractant for the green lacewing. Environmental Entomology 8, 1123–1125 (1979).
Tóth, M. et al. Phenylacetaldehyde: a chemical attractant for common green lacewings (Chrysoperla carnea s.l.; Neuroptera: Chrysopidae). European Journal of Entomology 103, 267–271 (2006).
Koczor, S., Szentkirályi, F., Fekete, Z. & Tóth, M. Smells good, feels good: oviposition of Chrysoperla carnea-complex lacewings can be concentrated locally in the field with a combination of appropriate olfactory and tactile stimuli. Journal of Pest Science 90, 311–317 (2017).
Schausberger, P., Peneder, S., Jurschik, S. & Hoffmann, D. Mycorrhiza changes plant volatiles to attract spider mite enemies. Functional Ecology 26, 441–449, doi:10.1111/j.1365-2435.2011.01947.x (2012).
Pangesti, N. et al. Rhizobacterial colonization of roots modulates plant volatile emission and enhances the attraction of a parasitoid wasp to host-infested plants. Oecologia 178, 1169–1180 (2015).
Whittaker, R. H. Evolution and Measurement of Species Diversity. Taxon 21, 213–251 (1972).
Chao, A. Nonparametric Estimation of the Number of Classes in a Population. Scandinavian Journal of Statistics 11, 265–270 (1984).
Faith, D. P. Conservation evaluation and phylogenetic diversity. Biological Conservation 61, 1–10 (1992).
Soil Survey Staff. Soil taxonomy. A basic system of soil classification for making and interpreting soil surveys. U.S. Department of Agriculture, Natural Resources Conservation (1999).
Wolf, D. C., Dao, T. H., Scott, H. D. & Lavy, T. L. Influence of sterilization methods on selected soil microbiological, physical, and chemical-properties. Journal of Environmental Quality 18, 39–44 (1989).
Vivas, A., Moreno, B., García-Rodriguez, S. & Benítez, E. Assessing the impact of composting and vermicomposting on bacterial community size and structure, and microbial functional diversity of an olive-mill waste. Bioresource Technology 100, 1319–1326 (2009).
Reddy, G. V. P. Plant volatiles mediate orientation and plant preference by the predator Chrysoperla carnea Stephens (Neuroptera: Chrysopidae). Biological Control 25, 49–55 (2002).
Klindworth, A. et al. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Research 41: e1, doi:10.1093/nar/gks808 (2013).
Hugerth, L. W. et al. Systematic design of 18S rRNA gene primers for determining eukaryotic diversity in microbial consortia. PLoS One 9(4), e95567 (2014).
Magoč, T. & Salzberg, S. L. FLASH: fast length adjustment of short reads to improve genome assemblies. Bioinformatics 27, 2957–2963 (2011).
Alekhin, A. et al. MG7: Configurable and scalable 16S metagenomics data analysis. bioRxiv 027714, doi:10.1101/027714 (2015).
Pasari, J. R., Levi, T., Zavaleta, E. S. & Tilman, D. Several scales of biodiversity affect ecosystem multifunctionality. Proceedings of the National Academy of Sciences 110, 10219–10222 (2013).
Hammer, Ø., Harper, D.A.T. & Ryan, P. D. Past: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4, 9 pp. (2001).
Davison, A. C. & Richard, I. Comparison of models for olfactometer data. Journal of Agricultural, Biological and Environmental Statistics 16, 157–169 (2010).
Zuur, A.F., Leno, E.N., Walker, N., Saveliev, A. A. & Smith, G. M. Mixed effects models and extensions in ecology with R (Springer, 2009).
Burnham, K. P. & Anderson, D. R. Model selection and multimodel inference: a practical information-theoretic approach (Springer-Verlag, 2002).
R Development Core Team. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria http://www.R-project.org (2016).
Caporaso, J. G. et al. QIIME allows analysis of high-throughput community sequencing data. Nature Methods 7, 335–336 (2010).
Ewing, B. & Green, P. Base-calling of automated sequencer traces using phred. II. Error probabilities. Genome research 8, 186–194 (1998).
Ewing, B., Hillier, L., Wendl, M. C. & Green, P. Base-calling of automated sequencer traces usingPhred. I. Accuracy assessment. Genome research 8, 175–185 (1998).
Edgar, R. C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461 (2010).
McDonald, D. et al. The Biological Observation Matrix (BIOM) format or: how I learned to stop worrying and love the ome-ome. GigaScience 1, 7 (2012).
Caporaso, J. G. et al. PyNAST: a flexible tool for aligning sequences to a template alignment. Bioinformatics 26, 266–267 (2010).
Price, M. N., Dehal, P. S. & Arkin, A. P. FastTree 2 - Approximately maximum-likelihood trees for large alignments. Plos One 5(3), doi:10.1371/journal.pone.0009490 (2010).
ter Braak, C. T. & Šmilauer, P. CANOCO reference manual and CanoDraw for Windows user’s guide: software for canonical community ordination (version 4.5). Section on Permutation Methods. Microcomputer Power New York (2002).
Acknowledgements
This work was supported by ERDF-cofinanced grant RECUPERA 2020 from the Spanish Ministry of Economy and Competitiveness-CSIC and Project AGR 1419 from Junta de Andalucía, Spain. E. Rodríguez held a postdoctoral contract (DOC-INIA program) granted by Spanish National Institute for Agricultural and Food Research and Technology (INIA) and the European Social Fund. The volatiles and chemical analyses were made at the Scientific Instrumentation Service, EEZ-CSIC, Granada, Spain. We would also like to thank Dr. Fernando Reyes from Fundación Medina for his help in discussing the results of the analysis of volatiles and Michael O´Shea for assisting in the translation of the original manuscript into English.
Author information
Authors and Affiliations
Contributions
E.B., B.M., E.R. and M.C. conceived the study and planned the experiments. E.B., B.M., D.P. and D.A. performed the experiments. E.B., B.M., D.P., M.C. and E.R. analysed data. R.N., M.R., D.A. and E.R. contributed data. E.B., B.M. and D.P. wrote the manuscript. All authors interpreted the results and reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Benítez, E., Paredes, D., Rodríguez, E. et al. Bottom-up effects on herbivore-induced plant defences: a case study based on compositional patterns of rhizosphere microbial communities. Sci Rep 7, 6251 (2017). https://doi.org/10.1038/s41598-017-06714-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-06714-x
This article is cited by
-
Modulation of volatile emissions in olive trees: sustained effect of Trichoderma afroharzianum T22 on induced plant defenses after simulated herbivory
Biology and Fertility of Soils (2024)
-
Ethnobotanical study on traditional use of local fruit varieties in Gargano Promontory (Apulia, Italy)
Genetic Resources and Crop Evolution (2022)
-
Improved 18S and 28S rDNA primer sets for NGS-based parasite detection
Scientific Reports (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
