
Abstract
Banisteriopsis caapi is the basic ingredient of ayahuasca, a psychotropic plant tea used in the Amazon for ritual and medicinal purposes, and by interested individuals worldwide. Animal studies and recent clinical research suggests that B. caapi preparations show antidepressant activity, a therapeutic effect that has been linked to hippocampal neurogenesis. Here we report that harmine, tetrahydroharmine and harmaline, the three main alkaloids present in B. caapi, and the harmine metabolite harmol, stimulate adult neurogenesis in vitro. In neurospheres prepared from progenitor cells obtained from the subventricular and the subgranular zones of adult mice brains, all compounds stimulated neural stem cell proliferation, migration, and differentiation into adult neurons. These findings suggest that modulation of brain plasticity could be a major contribution to the antidepressant effects of ayahuasca. They also expand the potential application of B. caapi alkaloids to other brain disorders that may benefit from stimulation of endogenous neural precursor niches.
Similar content being viewed by others
Introduction
Ayahuasca is the Quechua name used to designate Banisteriopsis caapi, a jungle liana of the Malpighiaceae family that is native to the Amazon and Orinoco river basins1. The term is also applied to the tea that is obtained by infusing in water the stems of B. caapi, alone or in combination with other plants2. The ayahuasca tea is a central element in the ancient shamanic practices and rites of passage of the indigenous inhabitants of northwestern South America. More recently, ayahuasca has become a central sacrament in the rituals of Brazilian syncretic religious groups. These so-called “ayahuasca religions” have popularized ayahuasca use with the expansion of their activities to North America and Europe3.
Ayahuasca has a complex chemistry and pharmacology. B. caapi contains high amounts of harmine and tetrahydroharmine (THH) and to a lesser degree harmaline, three indole alkaloids with β-carboline structure4. These alkaloids are reversible monoamine-oxidase-A (MAO-A) inhibitors5, while THH can also inhibit serotonin reuptake6. Although B. caapi can be the sole ingredient of the tea7, up to 100 different plants have been described as admixtures to ayahuasca. These plants contain a wide variety of psychotropic substances such as nicotine (from Nicotiana spp.), scopolamine (from Brugmansia spp.), caffeine (from Ilex guayusa and Paullinia yoco), cocaine (from Erythoxylum coca) and N,N-dimethyltryptamine (DMT, from Psychotria viridis and Diplopterys cabrerana)2, 8.
In recent years, non-traditional use of ayahuasca has expanded worldwide, especially the version of the tea that combines B. caapi with P. viridis, due to the visionary effects induced by DMT, a psychedelic serotonin-2A agonist9. This expansion and reports of health benefits derived from its use have stimulated research into the pharmacology and therapeutic potential of ayahuasca10. Two recent studies found that a single dose of ayahuasca rapidly reduced depressive symptoms in treatment-resistant patients11, 12. Remarkably, clinical improvement was maintained for up to three weeks. With the resurgence of research on psychedelics13, the β-carbolines of B. caapi have been considered to play a minor role in the overall pharmacology of ayahuasca preparations. However, this is probably a narrow view. B. caapi is the common ingredient to all ayahuasca brews, and chemical analyses have shown that while the β-carbolines are present in all ayahuasca samples, this is not always the case for DMT14, 15. For instance, an ayahuasca sample from the Brazilian Church União do Vegetal, a group known to regularly combine B. caapi with P. viridis in their ayahuasca, was found to contain no DMT at all16.
Due to their ubiquitous presence in ayahuasca, it can be hypothesized that the β-carbolines contribute to the CNS effects of the tea. Studies in animals have shown that harmine has antidepressant effects in behavioral animal models of depression17, 18. Responses after harmine in the forced swim and open field tests are analogous to those obtained with ayahuasca infusions prepared from B. caapi and P. viridis and containing DMT19, 20. These findings suggest that DMT is not essential for the behavioral responses observed in animals. Additionally, in contrast with more traditional antidepressants such as imipramine, harmine increases BDNF levels in the hippocampus after both acute and chronic administration18, 21. These data suggest that harmine and potentially the other β-carbolines present in B. caapi contribute to the therapeutic effects of ayahuasca observed in clinical studies involving patients with depression11, 12.
At the cellular level, antidepressant drug action has been linked to the ability of drugs to stimulate adult neurogenesis22. Neurogenesis is the process of generating functional neurons from progenitor cells. In the adult brain of mammals, neurogenesis occurs in two main niches: the subventricular zone (SVZ) of the lateral ventricle and the subgranular zone of the dentate gyrus of the hippocampus (SGZ). Neural stem cells in these areas can be induced to asymmetrically divide, generating new stem cells, and astrocytes, oligodendrocytes or neurons23, 24. These newly generated neurons have the capacity to migrate and integrate into existing neural circuits. The activity and phenotypic fate of neural stem cells is determined by both endogenous and exogenous factors25. Beyond understanding the mechanisms of adult neurogenesis, we are ultimately interested in its therapeutic capacity. Specifically, despite their various mechanism of action, clinically-effective antidepressants share the common feature of inducing neural stem cell proliferation and differentiation into new neurons26,27,28.
Here, we investigated the capacity of the three main β-carbolines present in B. caapi to induce neurogenesis in vitro using neural progenitor cells from adult mice. We decided to focus on the alkaloids found in all ayahuasca brews, rather than on nicotine, DMT, scopolamine or other psychoactive compounds that can be present in ayahuasca following the addition of the admixture plants mentioned above. To this end neurospheres were prepared from stem cells obtained from both the SVZ and the SGZ and treated with harmine, THH, harmaline, and harmol. The latter is present only in small amounts in B. caapi but is readily formed in vivo in humans via O-demethylation of harmine, which undergoes extensive first-pass metabolism4, 29, 30. As shown below, results showed that the B. caapi β-carboline alkaloids present in ayahuasca directly regulate proliferation, migration and differentiation of neural stem cells.
Results
B. caapi β-carbolines control the activity of neural progenitors
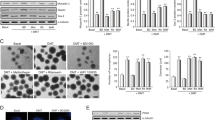
We isolated neural stem cells from the SVZ and the SGZ and we cultured them as free-floating neurospheres. Neurospheres from adult tissue are characterized by self-renewal and multipotent differentiation. To study the “stemness” of cultured neurospheres, we analyzed the expression of the following proteins: (a) musashi-1, a marker of undifferentiation; (b) nestin, an intermediate filament protein characteristic of neural stem/progenitor cells; and (c) “sex determining region Y-box 2” (SOX-2), a transcription factor essential for maintaining self-renewal and pluripotency of undifferentiated stem cells. We treated the neurosphere cultures for 7 days under proliferative conditions with each of the four β-carbolines: harmol, harmine, harmaline and tetrahydroharmine, all at 1 µM concentration. After treatment, we isolated the proteins and performed Western blots. Figure 1 clearly shows significant reductions in the amount of musashi-1, nestin and SOX-2 in the SVZ (musashi: F(4,15) = 49.87, p < 0.001; nestin: F(4,15) = 50.105, p < 0.001; SOX-2: F(4,15) = 121.684, p < 0.001) and SGZ (musashi: F(4,15) = 32.819, p < 0.001; nestin: F(4,15) = 141.556, p < 0.001; SOX-2: F(4,15) = 22.572, p < 0.001) neurospheres following treatment with the test compounds. These results indicated that the β-carbolines promote a loss of “stemness” or undifferentiated state of the neurospheres.

Effects of ayahuasca β-carboline alkaloids on stemness of cultured adult neurospheres. Representative Western blots and bar graphs showing expression levels of the precursor cell markers musashi-1, nestin and SOX-2 after treatment with each of the four alkaloids tested (1 µM). Values in bar graphs indicate mean ± SD of the quantification of at least three independent experiments corresponding to four different cellular pools. The left side of the image shows results for the subventricular zone (SVZ) of the brain. The right side of the image shows results for the subgranular zone of the hippocampus (SGZ). *p ≤ 0.05; **P ≤ 0.01; ***p ≤ 0.001 indicate significant results in the post-hoc pair-wise comparisons (Bonferroni) versus non-treated (basal) cultures.
B. caapi β-carbolines induce proliferation and growth in neurosphere cultures
We analyzed next whether the treatment with test compounds modulated proliferation capacity in the neurosphere. For this purpose, we grew free floating neurospheres under proliferative conditions, i.e., in the presence of epidermal growth factor (EFG) and fibroblast growth factor (FGF) for 7 days, with the addition of saline (control) or each of the four tested β-carbolines. As shown in Fig. 2, addition of the alkaloids to the cultures markedly increased the rate of formation and the size of the neurospheres.

Effects of ayahuasca β-carboline alkaloids on adult neurosphere formation. (a) Representative phase-contrast micrographs showing the number and size of neurospheres after 7 days in culture in the presence of each of the four alkaloids tested (1 µM). The number and diameter of at least 50 neurospheres was determined in control and treated cultures. Scale bar = 100 μm. (b) Bar graphs showing results as mean values ± SD of the quantification of at least three independent experiments corresponding to four different cellular pools. The left side of the image shows results for the subventricular zone (SVZ) of the brain. The right side of the image shows results for he subgranular zone of the hippocampus (SGZ). *p ≤ 0.05; **p ≤ 0.01; ***p ≤ 0.001 indicate significant results in the post-hoc pair-wise comparisons (Bonferroni) versus non-treated (basal) cultures.
After 7 days of β-carboline treatment, the number of SVZ-derived neurospheres was significantly higher than in the vehicle-treated cultures (F(4,15) = 466.512, p < 0.001). Analogous results were observed for neurospheres derived from the SGZ (F(4,15) = 1226.445, p < 0.001). Regarding the size of the neurospheres, significant increases were observed for both SVZ- and SGZ-derived neurospheres after β-carboline treatment, as compared with non-treated cultures (SVZ: F(4,15) = 126.858, p < 0.001); SGZ: F(4,15) = 172.617, p < 0.001). In view of these results, we next analyzed the expression of Ki67, a marker of dividing cells, and “proliferating cell nuclear antigen” (PCNA) (Fig. 3).

Effects of ayahuasca β-carboline alkaloids on adult neural stem cells proliferation. (a) Representative immunofluorescence images showing the expression of the cellular marker for proliferation ki67 (green) in neurospheres derived from the subventricular zone (SVZ) of the lateral ventricle and the subgranular zone (SGZ) of the hippocampus. SVZ-derived neurospheres are shown in two panels showing the central part of the sphere (left) and the distal migration site (right). Single images from SGZ-derived neurospheres show the whole neurosphere, including the central and distal areas. DAPI was used for nuclear staining. Scale bar = 50 μm. (b) Representative Western blots of ki67 and the proliferating cell nuclear antigen (PCNA) levels in neurospheres treated for 7 days with each of the four alkaloids tested (1 µM). Bar graphs show the results of the quantification analyses. Each bar indicates relative protein levels expressed as mean ± SD of the quantification of at least three independent experiments corresponding to four different cellular pools. The left side of the image shows results for the subventricular zone (SVZ) of the brain. The right side of the image shows results for he subgranular zone of the hippocampus (SGZ). *p ≤ 0.05; **p ≤ 0.01 indicate significant results in the post-hoc pair-wise comparisons (Bonferroni) versus non-treated (basal) cultures.
As expected, treatment of the cultures with the β-carbolines resulted in an increase in the number of Ki67- (SVZ: F(4,15) = 514.405, p < 0.001); SGZ: F(4,15) = 51.976, p < 0.001) and PCNA-positive cells (SVZ: F(4,15) = 7.905, p < 0.001); SGZ: F(4,15) = 42.213, p < 0.001), indicating a direct effect of these compounds on proliferation. These results indicated that the β-carbolines actively stimulate the proliferation and growth of neurospheres from both the SVZ and the SGZ by controlling the activity of neural progenitors.
B. caapi β-carbolines increase neural stem cell migration
In addition to proliferation and differentiation, neurogenesis involves the migration of neural stem cells and their integration into functional circuits. We thus tested next the effects of the β-carbolines on precursor migration. We treated the neurospheres with each of the test compounds and we monitored migration of the newly formed cells using livescanning microscopy for 48–72 h. The results are presented in Supplementary Figure 1 (and in the Supporting Information Videos 1–5). As shown therein, β-carboline treatment resulted in significant increases in migration capacity (SVZ: F(4,15) = 349.872, p < 0.001); SGZ: F(4,15) = 1203.666, p < 0.001) in comparison with basal (control). Neural stem cells moved long distances out of the neurosphere body in the presence of the four tested compounds. On the contrary, cells in the control cultures remained close to the neurosphere core.
B. caapi β-carbolines induce differentiation of neural stem cells
We investigated the capacity of the β-carbolines to promote cell differentiation into any of the three different cellular types that form the central nervous system: astrocytes, neurons and/or oligondendrocytes. We analyzed the formation of the different cell types using immunocytochemistry. Neurosphere cultures prepared from the SVZ and the SGZ were adhered to a substrate and cultured under differentiation conditions, that is, in the absence of growth factors and in the presence of 1% fetal bovine serum. The β-carbolines were added to the medium and, after 3 days in culture, we evaluated the expression of Tuj1- and MAP-2-positive cells by immunocytochemistry analysis (Fig. 4a) and we quantified these proteins by Western blot (Fig. 4b). After the three-day period, vehicle-treated cultures only showed a few positively-stained cells for Tuj-1 or MAP-2. On the contrary, in the β-carboline-treated cultures, the number of both Tuj1- (SVZ: F(4,15) = 16.295, p < 0.001); SGZ: F(4,15) = 45.997, p < 0.001) and MAP-2-positive cells (SVZ: F(4,15) = 41.631, p < 0.001); SGZ: F(4,15) = 22.951, p < 0.001) was significantly increased. These results indicate that the test compounds induce the differentiation of neural stem cells towards mature neurons.

Ayahuasca β-carboline alkaloids promote stem cell differentiation towards a neuronal phenotype. After 7 days on culture in the presence of harmol, harmine, harmaline and tetrahydroharmine (THH), free floating neurospheres derived from the adult subgranular (SGZ) and subventricular (SVZ) zone were adhered on coated coverslips and allowed to differentiate for 3 days in the presence of alkaloids at 1 µM. (a) Representative immunofluorescence images showing the expression of the neuronal markers β-III-Tubulin (TuJ-1 clone, green) and MAP-2 (red) in neurospheres. DAPI was used for nuclear staining. SVZ-derived neurospheres are shown in two panels showing the central part of the sphere (left) and the distal migration site (right). Single images from SGZ-derived neurospheres show the whole neurosphere, including the central and distal areas. Scale bar = 50 μm. (b) Representative Western blots of β-tubulin and MAP-2. Quantification analyses are also shown. Results are the mean ± SD of the quantification of at least three independent experiments corresponding to four different cellular pools. The left side of the image shows results for the subventricular zone (SVZ) of the brain. The right side of the image shows results for he subgranular zone of the hippocampus (SGZ). *p ≤ 0.05; **p ≤ 0.01 indicate significant results in the post-hoc pair-wise comparisons (Bonferroni) versus non-treated (basal) cultures.
Concerning the formation of astrocytes or oligodendrocytes, Fig. 5 shows that most of the test compounds did not induce the differentiation of neural precursors towards an oligondedrocyte phenotype. Results showed that only the harmol- and harmaline-treated SVZ cultures presented some scattered CNPase-positive cells (Fig. 5a). These results were further confirmed by quantification of the Western blots (Fig. 5b; SVZ: F(4,15) = 7.853, p < 0.001); SGZ: F(4,15) = 1.472, p > 0.05)). In the case of GFAP-positive cells, a high number was observed in basal conditions (Fig. 5) and this number was increased after β-carboline treatment (SVZ: F(4,15) = 43.838, p < 0.001); SGZ: F(4,15) = 136.268, p < 0.001), suggesting that these compounds also promote the differentiation of astroglial cells or astrocyte-like radial cells.

Ayahuasca β-carboline alkaloids promote astrogliogenesis. Neurospheres derived from the adult subgranular (SGZ) and subventricular (SVZ) zone were cultured in the presence of harmol, harmine, harmaline and tetrahydroharmine (THH). After 7 days neurospheres were adhered on coated coverslips and allowed to differentiate for 3 days in the presence of alkaloids at 1 µM. (a) Neurosphere immunofluorescence images showing in green the expression of CNPase (oligodendrocyte marker) and glial fibrillary acidic protein (GFAP, red) that stains astrocytes. SVZ-derived neurospheres are shown in two panels showing the central part of the neurosphere (left) and the distal migration site (right). Single images from SGZ-derived neurospheres show the distal part of the neurosphere. DAPI was used for nuclear staining. Scale bar = 50 μm. (b) Representative Western blots of CNPase and GFAP. Quantification analyses are also shown. Results are the mean ± SD of the quantification of at least three independent experiments corresponding to four different cellular pools. The left side of the image shows results for the subventricular zone (SVZ) of the brain. The right side of the image shows results for he subgranular zone of the hippocampus (SGZ). *p ≤ 0.05; **p ≤ 0.01 indicate significant results in the post-hoc pair-wise comparisons (Bonferroni) versus non-treated (basal) cultures.
Discussion
Here we showed that adult neural stem cell activity is regulated by harmine, THH, and harmaline, the most abundant alkaloids in B. caapi and ayahuasca, and by harmol, the main metabolite of harmine in humans31. Using an in vitro model of adult neurogenesis, we found that all four β-carbolines stimulated the proliferation and migration of progenitor cells and promoted their differentiation predominantly into neurons.
The four compounds tested effectively promoted proliferation, migration, and differentiation of progenitor cells obtained from the SVZ and the SGZ, the two main niches of adult neurogenesis in rodents. The β-carbolines increased the number and size of primary neurospheres, induced the loss of the neurospheres’ undifferentiated state, and promoted subsequent cell migration and differentiation mainly into a neuronal phenotype, as indicated by the positive expression of β-III-tubulin and MAP2, but also into astroglial cells. Taken together, these three effects indicate that B. caapi alkaloids have the capacity to regulate the expansion and fate of stem cell populations.
Analysis of the proliferation stage showed that all four β-carbolines increased the number and size of neurospheres, the number of Ki-67-stained cells, and the amounts of Ki-67 and PCNA protein as measured by Western blot. Our results for the effect of harmine on proliferation are in line with a previous study showing harmine-induced increase in mitosis in cultured chick embryo cells32, and in human neural progenitor cells33. To our knowledge, the effects of harmaline and THH on neurogenesis have not been studied before. While harmaline is only present in small amounts in B. caapi, THH is the second most abundant β-carboline in the plant4, 29. Additionally, THH shows more consistent plasma levels between individuals and studies than harmine, which is rapidly degraded to harmol when taken orally29, 34. The latter, formed in vivo by O-demethylation of the parent compound29, showed proliferative effects of similar magnitude to those of harmine.
Our results showed that B. caapi β-carbolines promoted cellular migration and differentiation, suggesting that these alkaloids not only act as mitogens for neural stem cells, but also modulate cellular fate. The largest effects on migration were observed for harmaline and THH. Increased migration capacity is relevant in certain conditions such as brain injury, where stem cell niches are far from the damaged area35,36,37. All tested compounds also promoted cellular differentiation. Neural stem cells are known to differentiate into neurons, astrocytes, and oligodendrocytes23, 24. The observed increases in Tuj-1 and MAP-2 protein expression indicated differentiation predominantly toward a neuronal phenotype. In the SVZ both proteins were equally expressed after each of the four treatments. However, in the SGZ harmine administration did not influence Tuj-1 levels, a marker of immature neurons, but significantly increased the expression of MAP-2, suggesting a larger impact on neuronal maturation.
All the above indicate that B. caapi β-carbolines facilitate neurogenesis at multiple levels. This capacity is of interest, since in pathological conditions the replacement of neurons may be optimized by acting simultaneously on various processes38, 39. The effect of the β-carbolines on cellular proliferation and differentiation is not unique to these compounds, having been observed for endogenous molecules such as leukotriene B440, BMPs41, the growth factors EGF/FGF242, and NGF/BDNF/bFGF43, and the transcription factors Lmx1a and Lmx1b44. However, the fact that the β-carbolines also stimulated migration highlights the versatility of these exogenous compounds, as they can promote the three processes involved in full adult neurogenesis.
A likely possible explanation for the observed effects of β-carbolines in neurogenesis is the increase in monoamine levels caused by MAO inhibition. With this said, we must acknowledge that the magnitude of the neurogenic effects was similar for the four compounds, despite harmol and THH being inhibitors that are between a hundred and a thousand times weaker than harmine or harmaline5. Moreover, the role of monoamines in neurogenesis is not fully understood. Knocking out the 5-HT1A receptor in mice impaired neurogenesis after fluoxetine but not after imipramine, indicating that neurogenesis was independent from elevated serotonin levels45. In another study, the authors reported the unexpected finding that serotonin depletion actually promoted hippocampal neurogenesis instead of decreasing it46. In a recent paper, harmine, but not the MAO inhibitor pargyline, stimulated proliferation of human neural progenitor cells in vitro 33. Harmine effects were mediated through inhibition of the DYRK1A kinase rather than through MAO inhibition. This opens the possibility that the β-carbolines tested here regulated stem cell fate via DYRK1A or other alternative mechanisms. To our knowledge, the inhibitory effects of harmaline, tetrahydroharmine and harmol on DYRK1A has not been examined. Other potential molecular targets for the neurogenic effects of small molecules include the modulation of the GSK-3β/β-catenin pathway 47, upregulation of the brain-derived neurotrophic factor48, increased levels of vascular endothelial growth factor49; and glucocorticoid receptor activation50. Future studies should assess whether B. caapi β-carbolines interact with one or more of these pathways.
Our findings have relevant therapeutic implications. The association between neurogenesis and anti-depressant activity is well documented45, 51. Enhanced hippocampal neurogenesis reduces depression-like behaviors in animals51. Furthermore, clinically effective antidepressants stimulate neurogenesis, independent of their chemical structure and mechanism of action. To cite a few examples, chronic treatment with the serotonin reuptake inhibitor fluoxetine increases neurogenesis in rats52, 53, as does chronic treatment with the selective MAO-A inhibitor pirlindole53. The association between neurogenesis and antidepressant effect is not limited to rodents and pharmacological interventions. Electroconvulsive therapy in primates also stimulates proliferation of neural precursors in the hippocampus and their differentiation into neurons54. Hippocampal neurogenesis appears to be necessary for antidepressant action. Irradiation of the SGZ of the hippocampus in mice prevents the neurogenic and behavioral effects of fluoxetine and imipramine45.
In humans, two recent clinical studies have demonstrated rapid and long-lasting antidepressant effects after a single ayahuasca dose in patients who did not respond to conventional treatment11, 12. The therapeutic potential of ayahuasca is an area of increasing research interest beyond depression55. Alterations in adult neurogenic niches have been associated with a number of pathologies affecting the central nervous system56,57,58,59. Stimulation of these niches is currently being investigated as a novel therapeutic strategy for neuropsychiatric disorders60,61,62. Regular ayahuasca use has been associated decreases in problematic alcohol, cocaine and opiate consumption, indicating anti-addiction properties for B. caapi preparations63, 64. These potential anti-addictive properties are particularly relevant if we acknowledge the notorious difficulty of treating substance use disorders. Drug-dependent patients not only show functional deficits in reward processing and cognitive control, but also structural alterations in brain gray and white matter65.
Our study has a series of limitations that need to be acknowledged. Ayahuasca brews contain other active compounds that were not tested here. A popular version of ayahuasca in the USA and Europe contains DMT, a serotonergic psychedelic9. It is possible that DMT may have contributed to the antidepressant effects reported in clinical studies using ayahuasca11, 12. This contribution could be due to both brain plasticity mediated by 5-HT2A receptor activation66 and to the profound psychological experiences induced by psychedelics67. While studying DMT in the neurogenesis model was not an objective of the present investigation, it could be assessed in a future study, comparing it with other 5-HT2A agonists such as psilocybin or LSD. Although several animal studies have already shown that harmine improves behavioral measures of depression17, 18, future studies could ideally test the four compounds assessed here for both in vivo neurogenesis and behavioral improvement. Finally, future research could also use positive controls to compare the potency of the B. caapi β-carbolines with that of other antidepressants, such as SSRIs and MAO inhibitors.
In conclusion, here we showed that the β-carboline alkaloids present in B. caapi, the plant source of the ayahuasca tea, promote neurogenesis in vitro by stimulating neural progenitor pool expansion, and by inducing cellular migration and differentiation into a neuronal phenotype. The stimulation of neurogenic niches in the adult brain may substantially contribute to the antidepressant effects reported for ayahuasca in recent clinical studies. The versatility and full neurogenic capacity of the B. caapi β-carbolines warrant further investigation of these compounds. Their ability to modulate brain plasticity indicates their therapeutic potential for a broad range of psychiatric and neurologic disorders.
Methods
Ethics
Animals used in this study were cared for following the animal experimental procedures previously approved by the “Ethics Committee for Animal Experimentation” of the Instituto de Investigaciones Biomédicas and carried out in accordance with the European Communities Council, directive 2010/63/EEC and National regulations, normative 53/2013.
Drugs
Harmine and harmaline were obtained from commercial sources (Sigma-Aldrich). Harmol and tetrahydroharmine were obtained by synthesis following the procedures described below.
Synthesis of harmol and tetrahydroharmine
1-Methyl-9H-pyrido[3,4-b]indol-7-ol hydrobromide (harmol·HBr)
To a solution of harmine (151 mg, 0.71 mmol, Sigma-Aldrich) in acetic acid (3 mL) hydrobromic acid (3 mL, 47% aqueous solution) was added. The mixture was heated at 160 °C during 25 min in a microwave oven and then allowed to cool to room temperature. After few minutes at 25 °C, needles started to form and harmol·HBr·H2O (180 mg, 85% yield) was recovered by filtration as a yellow solid of mp 250–251 °C. 1H NMR (400 MHz, D2O) δ 7.62 (d, J = 5.4 Hz, 1 H), 7.39 (d, J = 5.4 Hz, 1 H), 7.18 (d, J = 8.4 Hz, 1 H), 6.26 (d, J = 8.5 Hz, 1 H), 5.96 (s, 1 H), 2.31 (s, 3 H). 13C NMR (101 MHz, D2O) δ 158.55, 143.69, 134.75, 132.07, 131.27, 127.49, 122.97, 112.87, 112.18, 111.48, 95.12, 14.77. HPLC purity 100% (230 to 400 nm). HRMS (ESI+): m/z calcd for C12H10N2O: 198.07931, found: 198.07934. Anal. Calcd for C12H10N2O·HBr·H2O (297.15): C, 48.50; H, 4.41; N, 9.43. Found: C 48.08; H 4.29; N 9.52.
7-Methoxy-1-methyl-2,3,4,9-tetrahydro-1H-pyrido[3,4-b]indole hydrochloride (tetrahy-droharmine·HCl, THH·HCl)
Sodium borohydride (680 mg, 18.00 mmol) was added in portions to a stirred solution of harmaline (1.54 g, 7.20 mmol, Sigma-Aldrich) in water (40 mL) at 0 °C and the mixture was acidified until pH 2 with aqueous HCl (2 M). After 75 min at room temperature, the mixture was made alkaline with NaOH (10% aqueous solution), extracted with dichloromethane (3 × 20 mL) and evaporated to dryness. Solid was dissolved in isopropyl alcohol, heated and then treated with an excess of concentrated HCl until a precipitate appeared. After 3 days at 4 °C, THH·HCl was collected by filtration (1.40 g, 77% yield) as a white solid of mp 201–202 °C. 1H NMR (400 MHz, DMSO) δ 10.98 (s, 1 H), 7.34 (d, J = 8.6 Hz, 1 H), 6.85 (d, J = 2.2 Hz, 1 H), 6.68 (dd, J = 8.6, 2.2 Hz, 1 H), 4.68 (q, J = 6.8 Hz, 1 H), 3.76 (s, 3 H), 3.59 (dt, J = 12.4, 4.7 Hz, 1 H), 3.37–3.27 (m, 1 H), 2.97–2.81 (m, 2 H), 1.59 (d, J = 6.7 Hz, 3 H). 13C NMR (101 MHz, DMSO) δ 155.93, 136.95, 129.62, 120.17, 118.75, 108.97, 105.14, 94.67, 55.19, 48.42, 40.40, 18.30, 17.24. HPLC purity 98% (230 to 400 nm). Anal. Calcd for C13H16N2O·HCl (252.74): C, 61.78; H, 6.78; N, 11.08. Found: C 61.46; H 6.86; N 10.84.
Adult precursor isolation
Neural stem cells (NSCs) were isolated from the subgranular zone (SGZ) of the hippocampus and the subventricular zone (SVZ) of the lateral ventricle of adult C57BL/6 mice (3 months of age) and prepared following previously described methods68. A total number of 40 animals were used throughout the whole study, divided in four pools of 10 animals each. Every pool was individually used to perform all the experiments described in this section. Briefly, tissue was carefully dissected, dissociated in DMEM medium with glutamine, gentamicin and fungizone and then digested with 0.1% trypsin-EDTA + 0.1% DNAase + 0.01% hialuronidase for 15 min at 37 °C. The isolated stem cells were seeded into 12-well dishes at a density of ~40,000 cells per cm2 in DMEM/F12 (1:1) containing 10 ng/mL epidermal growth factor (EGF), 10 ng/mL fibroblast growth factor (FGF) and N2 medium.
Neurosphere cultures and treatments
NSCs were cultured under standard conditions and in the presence of growth factors (EGF and FGF) for a week, in 12-well dishes. After this time, small neural progenitor-enriched growing spheres known as neurospheres (NS) were formed. At this point, with all NS having the same stage and size, cultures were treated daily for 7 days with vehicle or 1 μM solutions of each of the four β-carbolines tested: harmol, harmine, harmaline and tetrahydroharmine (THH). The effective dose of compounds was chosen based on our previous studies on β-carboline analogs69 and the 7-day period was selected as a standard time frame used previously by our own and other research groups to test the neurogenic potential of drugs37, 50, 68, 70. The 1 μM concentrations chosen for the β-carbolines was lower than the 7.5 μM found to induce optimal proliferation in a previous in vitro study with harmine33 but closer to the plasma concentrations of 0.52–1.52 μM reported for harmine and THH, respectively, in a recent study involving ayahuasca administration to healthy humans71. At these concentrations, none of the tested β-carbolines affected the viability of the cultured cells. Proliferation and growth analysis was assessed from these cultures and the number and diameter of NS were quantified using the Nikon Digital Sight, SD-L1 software. All the NS contained in ten wells per condition were counted. Some of these proliferating NS were used for immunoblotting analysis, while others were seeded onto poly-L-lysine precoated-6-well plates, and/or -coverslips and cultured in the presence of β-carboline alkaloids (1 μM) under proliferation conditions (medium containing 1% fetal bovine serum and without exogenous growth factors). Once neurospheres were differentiated (72 h), those grown on coated 6-well plates were used for immunoblotting and those on coverslips for immunocytochemical analysis. The remaining NS were cultured for 48–72 h under differentiation conditions onto μ-Slide 8-well plates (Ibidi, Martinsried, Germany) and used for migration assays.
Immunocytochemistry
Cells were processed for immunocytochemistry as previously described69. Briefly, NS grown on glass were previously fixed 15 minutes at room temperature in 4% paraformaldehyde and incubated at 37 °C for 1 h with primary antibodies directed against ki67 (1/200, rabbit, Abcam), β-III-tubulin (1/400, TuJ-1 clone; rabbit; Abcam), MAP-2 (1/200, mouse; Sigma), CNPase (rabbit, Covance) and GFAP (1/500, mouse; Sigma). After several rinses in PBS, samples were then incubated with Alexa-488 goat anti-rabbit and Alexa-647 goat anti-mouse antibodies (1/500, Molecular Probes) for 45 min at 37 °C. Staining of nuclei was performed using 4′,6-diamidino-2-phenylindole (DAPI, 1/500). Finally images were acquired in a LSM710 laser scanning spectral confocal microscope (Zeiss). Confocal microscope settings were adjusted to produce the optimum signal-to-noise ratio.
Immunoblot analysis
Differentiated cultured NS were re-suspended in ice-cold cell lysis buffer (Cell Signaling Technology) with protease inhibitor cocktail (Roche) and incubated for 15–30 min on ice. A total amount of 30 µg of protein was loaded on a 10% or 12% SDS-PAGE gel and transferred to nitrocellulose membranes (Protran, Whatman). The membranes were blocked in Tris-buffered saline with 0.05% Tween-20 and 5% skimmed milk or 4% BSA (MAP-2 blots), incubated with primary and secondary antibodies, and washed according to standard procedures. Primary antibodies used were: musashi-1 (1/500, rabbit; Abcam), nestin (1/500, rabbit; Abcam), SOX-2 (1/1000, mouse, Cell Signaling), ki67 (1/500, rabbit, Abcam), PCNA (1/500, mouse, Millipore), β-III-tubulin (1/1000, TuJ clone; mouse; Covance), MAP-2 (1/500, mouse; Sigma), CNPase (1/500, rabbit, Covance), GFAP (1/1000, mouse; Sigma) and α-tubulin (1/5000, mouse; Sigma). Secondary peroxidase-conjugated (1/1000) donkey anti-rabbit (Amersham Biosciences, GE Healthcare), or rabbit anti-mouse antibodies (Jackson Immunoresearch) were used. Values in figures are the average of the quantification of at least three independent experiments each of them corresponding to four different cellular pools.
Cellular migration assay
Free-floating single spheres were individually picked up with a pipette and plated onto μ-Slide 8-well plates as previously described70. Images were acquired with a Cell Observer system from Zeiss (Jena, Germany) using a Zeiss Observer Z1 microscope, equipped with a Cascade 1 K camera, a monitorized X/Y stage, and a Module S incubator with equipment for temperature and CO2 control. During all experiments cells were kept in a humidified atmosphere of 5% CO2 in air at 37 °C. Axio Vision Rel. 4.8 software (Zeiss) for time-lapse imaging and cell tracking was used. Phase contrast images of cells were taken every 60 min using a × 4 objective (Achromatic, Zeiss). Cell migration was scored in at least 10 independent experiments per condition. The farthest distance of cell migration was calculated from the edge of the sphere. Resulting movies were collected and exported as avi files and are shown at 5 frames per second (See Supporting Videos 1–5).
Statistical analysis
Data were analyzed using a one-way ANOVA with treatment as factor (control, harmol, harmine, harmaline, THH). Significant results in the ANOVA (p < 0.05) were followed by post-hoc pair-wise test (Bonferroni). The SPSS statistical software package (version 20.0) for Windows (Chicago, IL, USA) was used for all statistical analyses.
References
Schultes, R. E. The botany and chemistry of hallucinogens. (Thomas, 1980).
McKenna, D. & Riba, J. New World Tryptamine Hallucinogens and the Neuroscience of Ayahuasca. Curr. Top. Behav. Neurosci. doi:10.1007/7854_2016_472 (2017).
Tupper, K. W. The globalization of ayahuasca: harm reduction or benefit maximization? Int. J. Drug Policy 19, 297–303 (2008).
Riba, J. Human Pharmacology of Ayahuasca. (Autonomous University of Barcelona, 2003).
Buckholtz, N. S. & Boggan, W. O. Monoamine oxidase inhibition in brain and liver produced by beta-carbolines: structure-activity relationships and substrate specificity. Biochem. Pharmacol. 26, 1991–1996 (1977).
Buckholtz, N. S. & Boggan, W. O. Inhibition by beta-carbolines of monoamine uptake into a synaptosomal preparation: structure-activity relationships. Life Sci. 20, 2093–2099 (1977).
Spruce, R. Notes of a botanist on the Amazon and Andes 2 (Macmillan, 1908).
Ott, J. Pharmacotheon: entheogenic drugs, their plant sources and history. (Natural Products Co, 1993).
Ray, T. S. Psychedelics and the human receptorome. PloS One 5, e9019 (2010).
Domínguez-Clavé, E. et al. Ayahuasca: Pharmacology, neuroscience and therapeutic potential. Brain Res. Bull. 126, 89–101 (2016).
Osório, F. de L. et al. Antidepressant effects of a single dose of ayahuasca in patients with recurrent depression: a preliminary report. Rev. Bras. Psiquiatr. São Paulo Braz. 1999 37, 13–20 (2015).
Sanches, R. F. et al. Antidepressant Effects of a Single Dose of Ayahuasca in Patients With Recurrent Depression: A SPECT Study. J. Clin. Psychopharmacol. 36, 77–81 (2016).
Sessa, B. Shaping the renaissance of psychedelic research. Lancet Lond. Engl. 380, 200–201 (2012).
Rivier, L. & Lindgren, J. E. ‘Ayahuasca’, the South American hallucinogenic drink: an ethnobotanical and chemical investigation. Econ. Bot. 26, 101–129 (1972).
McKenna, D. J., Towers, G. H. & Abbott, F. Monoamine oxidase inhibitors in South American hallucinogenic plants: tryptamine and beta-carboline constituents of ayahuasca. J. Ethnopharmacol. 10, 195–223 (1984).
Callaway, J. C. Various alkaloid profiles in decoctions of Banisteriopsis caapi. J. Psychoactive Drugs 37, 151–155 (2005).
Farzin, D. & Mansouri, N. Antidepressant-like effect of harmane and other beta-carbolines in the mouse forced swim test. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 16, 324–328 (2006).
Fortunato, J. J. et al. Acute harmine administration induces antidepressive-like effects and increases BDNF levels in the rat hippocampus. Prog. Neuropsychopharmacol. Biol. Psychiatry 33, 1425–1430 (2009).
Pic-Taylor, A. et al. Behavioural and neurotoxic effects of ayahuasca infusion (Banisteriopsis caapi and Psychotria viridis) in female Wistar rat. Behav. Processes 118, 102–110 (2015).
Lima, L.-M. et al. Les effets de l’ayahuasca sur le système nerveux central: étude comportementale. Phytothérapie 5, 254–257 (2007).
Fortunato, J. J. et al. Chronic administration of harmine elicits antidepressant-like effects and increases BDNF levels in rat hippocampus. J. Neural Transm. Vienna Austria 1996 117, 1131–1137 (2010).
Yun, S., Reynolds, R. P., Masiulis, I. & Eisch, A. J. Re-evaluating the link between neuropsychiatric disorders and dysregulated adult neurogenesis. Nat. Med. 22, 1239–1247 (2016).
Gage, F. H. Mammalian neural stem cells. Science 287, 1433–1438 (2000).
Temple, S. Stem cell plasticity–building the brain of our dreams. Nat. Rev. Neurosci. 2, 513–520 (2001).
Gonçalves, J. T., Schafer, S. T. & Gage, F. H. Adult Neurogenesis in the Hippocampus: From Stem Cells to Behavior. Cell 167, 897–914 (2016).
Chen, S.-J. et al. Antidepressant administration modulates neural stem cell survival and serotoninergic differentiation through bcl-2. Curr. Neurovasc. Res. 4, 19–29 (2007).
Boldrini, M. et al. Antidepressants increase neural progenitor cells in the human hippocampus. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 34, 2376–2389 (2009).
Sachs, B. D. & Caron, M. G. Chronic fluoxetine increases extra-hippocampal neurogenesis in adult mice. Int. J. Neuropsychopharmacol. Off. Sci. J. Coll. Int. Neuropsychopharmacol. CINP 18 (2015).
Riba, J. et al. Human pharmacology of ayahuasca: subjective and cardiovascular effects, monoamine metabolite excretion, and pharmacokinetics. J. Pharmacol. Exp. Ther. 306, 73–83 (2003).
Riba, J., McIlhenny, E. H., Bouso, J. C. & Barker, S. A. Metabolism and urinary disposition of N,N-dimethyltryptamine after oral and smoked administration: a comparative study. Drug Test. Anal. 7, 401–406 (2015).
Riba, J., McIlhenny, E. H., Valle, M., Bouso, J. C. & Barker, S. A. Metabolism and disposition of N,N-dimethyltryptamine and harmala alkaloids after oral administration of ayahuasca. Drug Test. Anal. 4, 610–616 (2012).
Hämmerle, B. et al. Transient expression of Mnb/Dyrk1a couples cell cycle exit and differentiation of neuronal precursors by inducing p27KIP1 expression and suppressing NOTCH signaling. Dev. Camb. Engl. 138, 2543–2554 (2011).
Dakic, V. et al. Harmine stimulates proliferation of human neural progenitors. PeerJ 4, e2727 (2016).
Callaway, J. C. Fast and slow metabolizers of Hoasca. J. Psychoactive Drugs 37, 157–161 (2005).
Arvidsson, A., Collin, T., Kirik, D., Kokaia, Z. & Lindvall, O. Neuronal replacement from endogenous precursors in the adult brain after stroke. Nat. Med. 8, 963–970 (2002).
Sun, L., Lee, J. & Fine, H. A. Neuronally expressed stem cell factor induces neural stem cell migration to areas of brain injury. J. Clin. Invest. 113, 1364–1374 (2004).
Morales-Garcia, J. A. et al. Phosphodiesterase 7 inhibition induces dopaminergic neurogenesis in hemiparkinsonian rats. Stem Cells Transl. Med. 4, 564–575 (2015).
Alvarez-Buylla, A. & Lim, D. A. For the long run: maintaining germinal niches in the adult brain. Neuron 41, 683–686 (2004).
Lie, D. C., Song, H., Colamarino, S. A., Ming, G. & Gage, F. H. Neurogenesis in the adult brain: new strategies for central nervous system diseases. Annu. Rev. Pharmacol. Toxicol. 44, 399–421 (2004).
Wada, K. et al. Leukotriene B4 and lipoxin A4 are regulatory signals for neural stem cell proliferation and differentiation. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 20, 1785–1792 (2006).
Chen, H.-L. & Panchision, D. M. Concise review: bone morphogenetic protein pleiotropism in neural stem cells and their derivatives–alternative pathways, convergent signals. Stem Cells Dayt. Ohio 25, 63–68 (2007).
Bressan, R. B. et al. EGF-FGF2 stimulates the proliferation and improves the neuronal commitment of mouse epidermal neural crest stem cells (EPI-NCSCs). Exp. Cell Res. 327, 37–47 (2014).
Chen, S.-Q., Cai, Q., Shen, Y.-Y., Cai, X.-Y. & Lei, H.-Y. Combined use of NGF/BDNF/bFGF promotes proliferation and differentiation of neural stem cells in vitro. Int. J. Dev. Neurosci. Off. J. Int. Soc. Dev. Neurosci. 38, 74–78 (2014).
Yan, C. H., Levesque, M., Claxton, S., Johnson, R. L. & Ang, S.-L. Lmx1a and lmx1b function cooperatively to regulate proliferation, specification, and differentiation of midbrain dopaminergic progenitors. J. Neurosci. Off. J. Soc. Neurosci. 31, 12413–12425 (2011).
Santarelli, L. et al. Requirement of hippocampal neurogenesis for the behavioral effects of antidepressants. Science 301, 805–809 (2003).
Song, N.-N. et al. Reducing central serotonin in adulthood promotes hippocampal neurogenesis. Sci. Rep. 6, 20338 (2016).
Hui, J. et al. Fluoxetine Regulates Neurogenesis In Vitro Through Modulation of GSK-3β/β-Catenin Signaling. Int. J. Neuropsychopharmacol 18 (2015).
Taliaz, D., Stall, N., Dar, D. E. & Zangen, A. Knockdown of brain-derived neurotrophic factor in specific brain sites precipitates behaviors associated with depression and reduces neurogenesis. Mol. Psychiatry 15, 80–92 (2010).
Warner-Schmidt, J. L. & Duman, R. S. VEGF is an essential mediator of the neurogenic and behavioral actions of antidepressants. Proc. Natl. Acad. Sci. USA 104, 4647–4652 (2007).
Anacker, C. et al. Antidepressants increase human hippocampal neurogenesis by activating the glucocorticoid receptor. Mol. Psychiatry 16, 738–750 (2011).
Hill, A. S., Sahay, A. & Hen, R. Increasing Adult Hippocampal Neurogenesis is Sufficient to Reduce Anxiety and Depression-Like Behaviors. Neuropsychopharmacology 40, 2368–2378 (2015).
Marcussen, A. B., Flagstad, P., Kristjansen, P. E. G., Johansen, F. F. & Englund, U. Increase in neurogenesis and behavioural benefit after chronic fluoxetine treatment in Wistar rats. Acta Neurol. Scand. 117, 94–100 (2008).
Morais, M. et al. The effects of chronic stress on hippocampal adult neurogenesis and dendritic plasticity are reversed by selective MAO-A inhibition. J. Psychopharmacol. Oxf. Engl. 28, 1178–1183 (2014).
Perera, T. D. et al. Antidepressant-induced neurogenesis in the hippocampus of adult nonhuman primates. J. Neurosci. Off. J. Soc. Neurosci. 27, 4894–4901 (2007).
Frood, A. Ayahuasca psychedelic tested for depression. Nature. doi:10.1038/nature.2015.17252 (2015).
Baker, S. A., Baker, K. A. & Hagg, T. Dopaminergic nigrostriatal projections regulate neural precursor proliferation in the adult mouse subventricular zone. Eur. J. Neurosci. 20, 575–579 (2004).
Höglinger, G. U. et al. Dopamine depletion impairs precursor cell proliferation in Parkinson disease. Nat. Neurosci. 7, 726–735 (2004).
Borta, A. & Höglinger, G. U. Dopamine and adult neurogenesis. J. Neurochem. 100, 587–595 (2007).
Lennington, J. B. et al. Midbrain dopamine neurons associated with reward processing innervate the neurogenic subventricular zone. J. Neurosci. Off. J. Soc. Neurosci. 31, 13078–13087 (2011).
Luskin, M. B., Zigova, T., Soteres, B. J. & Stewart, R. R. Neuronal progenitor cells derived from the anterior subventricular zone of the neonatal rat forebrain continue to proliferate in vitro and express a neuronal phenotype. Mol. Cell. Neurosci. 8, 351–366 (1997).
Gage, F. H., Kempermann, G., Palmer, T. D., Peterson, D. A. & Ray, J. Multipotent progenitor cells in the adult dentate gyrus. J. Neurobiol. 36, 249–266 (1998).
Curtis, M. A., Faull, R. L. M. & Eriksson, P. S. The effect of neurodegenerative diseases on the subventricular zone. Nat. Rev. Neurosci. 8, 712–723 (2007).
Fábregas, J. M. et al. Assessment of addiction severity among ritual users of ayahuasca. Drug Alcohol Depend. 111, 257–261 (2010).
Thomas, G., Lucas, P., Capler, N. R., Tupper, K. W. & Martin, G. Ayahuasca-assisted therapy for addiction: results from a preliminary observational study in Canada. Curr. Drug Abuse Rev. 6, 30–42 (2013).
Vaquero, L. et al. Cocaine addiction is associated with abnormal prefrontal function, increased striatal connectivity and sensitivity to monetary incentives, and decreased connectivity outside the human reward circuit. Addict. Biol., doi:10.1111/adb.12356 (2016).
Vollenweider, F. X. & Kometer, M. The neurobiology of psychedelic drugs: implications for the treatment of mood disorders. Nat. Rev. Neurosci. 11, 642–651 (2010).
Griffiths, R. R. et al. Psilocybin produces substantial and sustained decreases in depression and anxiety in patients with life-threatening cancer: A randomized double-blind trial. J. Psychopharmacol. Oxf. Engl. 30, 1181–1197 (2016).
Morales-Garcia, J. A. et al. Phosphodiesterase7 Inhibition Activates Adult Neurogenesis in Hippocampus and Subventricular Zone In Vitro and In Vivo. Stem Cells 35, 458–472 (2017).
de la Fuente Revenga, M. et al. Neurogenic Potential Assessment and Pharmacological Characterization of 6-Methoxy-1,2,3,4-tetrahydro-β-carboline (Pinoline) and Melatonin-Pinoline Hybrids. ACS Chem. Neurosci. 6, 800–810 (2015).
Morales-Garcia, J. A. et al. Glycogen synthase kinase 3 inhibition promotes adult hippocampal neurogenesis in vitro and in vivo. ACS Chem. Neurosci. 3, 963–971 (2012).
Schenberg, E. E. et al. Acute Biphasic Effects of Ayahuasca. PloS One 10, e0137202 (2015).
Acknowledgements
This work was supported by the MINECO (Grants SAF2010-16365 and SAF2014-52940-R to A.P-C and grants SAF2012-31035 and SAF2015-64948-C2-1-R to M.I.R.-F.), and was partially financed with FEDER funds. J.R. received funding from the Beckley Foundation. CIBERNED is funded by the Instituto de Salud Carlos III. J.A.M-G. is a post-doctoral fellow from CIBERNED.
Author information
Authors and Affiliations
Contributions
A.P.-C. and J.R., conceived the project, designed and supervised the research, analyzed the data and wrote the manuscript. J.A.M.-G., planned the project, performed experimental work, analyzed data and wrote the manuscript. M.F.-R., contributed to the data analysis and wrote the manuscript. S.A.-G., performed experimental work. M.I.R.-F. and A.F. participated in experimental design and data interpretation.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Morales-García, J.A., de la Fuente Revenga, M., Alonso-Gil, S. et al. The alkaloids of Banisteriopsis caapi, the plant source of the Amazonian hallucinogen Ayahuasca, stimulate adult neurogenesis in vitro . Sci Rep 7, 5309 (2017). https://doi.org/10.1038/s41598-017-05407-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-05407-9
This article is cited by
-
Neurobiological research on N,N-dimethyltryptamine (DMT) and its potentiation by monoamine oxidase (MAO) inhibition: from ayahuasca to synthetic combinations of DMT and MAO inhibitors
Cellular and Molecular Life Sciences (2024)
-
Long-term ayahuasca use is associated with preserved global cognitive function and improved memory: a cross-sectional study with ritual users
European Archives of Psychiatry and Clinical Neuroscience (2024)
-
The effects of ketamine and classic hallucinogens on neurotrophic and inflammatory markers in unipolar treatment-resistant depression: a systematic review of clinical trials
European Archives of Psychiatry and Clinical Neuroscience (2023)
-
Peripheral modulation of antidepressant targets MAO-B and GABAAR by harmol induces mitohormesis and delays aging in preclinical models
Nature Communications (2023)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
