
Abstract
Reduction of seed dormancy mechanisms, allowing for rapid germination after planting, is a recurrent trait in domesticated plants, and can often be linked to changes in seed coat structure, in particular thinning. We report evidence for seed coat thinning between 2,000 BC and 1,200 BC, in southern Indian archaeological horsegram (Macrotyloma uniflorum), which it has been possible to document with high precision and non-destructively, through high resolution x-ray computed tomography using a synchrotron. We find that this trait underwent stepped change, from thick to semi-thin to thin seed coats, and that the rate of change was gradual. This is the first time that the rate of evolution of seed coat thinning in a legume crop has been directly documented from archaeological remains, and it contradicts previous predictions that legume domestication occurred through selection of pre-adapted low dormancy phenotypes from the wild.
Similar content being viewed by others
Introduction
Crop domestication is a co-evolutionary process which adapted plants to cultivation and facilitated the growth of larger sedentary human populations1, 2. The documentation of domestication in the archaeological record is often hampered by limited preservation of diagnostic morphology in archaeological plant specimens, with seed dispersal traits preserved in some cereals and grain size information in a wider range of taxa3, 4. Reduction of seed dormancy mechanisms, allowing for rapid germination after planting, is a recurrent trait in domesticated plants, and can often be linked to changes in seed coat structure, in particular thinning5, but this has been documented previously in sub-fossil material of only a few taxa, such as New World Chenopodium spp.4, 6, 7. In pulses (domesticated Fabaceae seed crops) reduction of seed dormancy is regarded as central to domestication as wild legumes have conspicuous dormancy related to thick, hard seed coats8,9,10,11. In some legumes, such as pea (Pisum sativum) the reduction in dormancy is linked to a conspicuous reduction in seed coat sculpturing9, but this is by no means universal across pulses. It has been inferred that only rare, wild plants that had reduced dormancy would have been suitable for early cultivators to select from the wild for cultivation8, 12, 13. However, archaeobotanical evidence documenting the presence of thin-seed coated, low dormancy phenotypes prior to or at the beginnings of cultivation has never been reported, nor have archaeobotanists previously documented evidence for the evolution, whether rapid or gradual, of pulse testa morphology. While this has been discussed largely in relation to Southwest Asian domesticates such a lentil (Lens culinaris) and pea8,9,10, 12, 13, it should apply equally across all pulses. Using a novel methodology to non-destructively document seed coat thickness around entire archaeological pulse seeds, we report data from the South Asia pulse crop horsegram (Macrotyloma uniflorum).
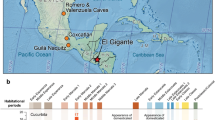
Horsegram is an annual native to South Asia14, where it has been an important crop since the beginnings of agriculture in many parts of South Asia. It is the most widely recovered pulse crop on prehistoric or early historic sites in India15. Today it is the fifth most widely cultivated grain legume in India, and arguably the most hardy, as well as a key vegetable protein source for hundreds of millions of rural inhabitants on the subcontinent16, 17. Evidence from the wild progenitor (Macrotyloma uniflorum var. stenocarpum) has been poorly studied and has never been described in the floristic studies of India. Thus, the regional origins of Horsegram still remain largely obscure18. Recent archaeobotanical sampling in the Deccan plateau of South India, a large, arid region featuring rich Neolithic period remains5, 18,19,20,21 has shown that a number of the earliest Southern Neolithic crop domesticates, including horsegram, appear to have been locally domesticated. Thus, based upon this evidence and limited assessment of herbarium specimens from the Botanical Survey of India, Pune an inferred possible South Indian origin has been postulated15, suggesting that wild horsegram was native to the semi-arid scrub savannah environments ranging from the Acacia thickets from the Aravalli hills in Rajasthan, the savannahs of the Southern Peninsula22, while the lower slopes of the western Himalayas are all also plausible foci of early cultivation or domestication (Fig. 1)23, 24. Previously, it had been simply assumed that all archaeological examples were domesticated crops, lending speculation to the question of where and when the domestication process took place25. In contrast, here we report findings for morphological change dating to this domestication process.

Map showing the distribution of early archaeological horsegram, indicating those sites included in the present study (Pa = Paithan, Pi = Piklihal, H = Hallur, S = Sanganakallu). Additional sites with horsegram recorded (1) Arikamedu (2) Veerapuram (3) Vikrampura Bangladesh (4) Ter (5) Saunphari (6) Perur (7) Noh (8) Nevasa (9) Ahirua Rajarampur (10) Brahmagiri (11) Ostapur (12) Inamgaon (13) Watgal (14) Tuljapur Garhi (15) Tokwa (16) Tekkalakota (17) Singanapalle (18) Sanghol (19) Rojdi (20) Rohira (21) Peddamudiyam (22) Paiyampalli (23) Paithan (24) Ojiyana (25) Narhan (26) Mithathal (27) Malhar (28) Ludwala (29) Kurugodu (30) Kunal (31) Kaothe (32) Kadebakele (33) Kayatha (34) Iinjedu (35) Hulas (36) Hiregudda (37) Hattibelagallu (38) Harirajpur (39) Bhagimohari (41) Banawali (42) Golbai Sassan (43) Apegaon (44) Adam (45) Daimabad (46) Kanmer (47) Lotehswar. Also shown the predicted original wild range. Map created using QGIS Development Team, QGIS 2.12.3- Lyon 2015. QGIS Geographic Information System. Open Source Geospatial Foundation Project. http://www.qgis.org/.
It may be that wild horsegram populations were more widespread during the mid-Holocene climatic wet phase, and the subsequent reduction of their availability, during the aridification that began in the later Fourth Millennium BC, may be connected to their domestication and the emergence of the Southern Neolithic (which coincide largely with the territory of the modern states of Karnataka and parts of Telangana); as these hunter/gatherer/foragers began to collect, concentrate and tend to patches of horsegram in their seasonal rounds15, 22, 26,27,28. Archaeobotanically, horsegram is widely reported and recovered from Chalcolithic and Neolithic sites, with the earliest known occurrence from the site of Khujhun, in the Vindhyan plateau29, 30, the Harappan site of Burthana Tigrana in Haryana31 and Southern Neolithic sites of Andhra and Karnataka29, 32, 33 (Fig. 1). In South India, horsegram is regarded as one of the co-staple crops in an indigenous package of crop domesticates that also included mungbean (Vigna radiata) and the millet, browntop millet (Brachiaria ramosa)26. Together these crops provided the economic base for the emergence of sedentary villages on the Indian peninsula between 5,000 and 4,000 years ago (3,000–2,000 BC)19, 34, 35.
Grain size increase is a trait used to measure rates of domestication which has been documented in several pulses including mungbean, lentil, pea, chickpea (Cicer arietinum), and soybean (Glycine max)4. Horsegram metrics from the Deccan region, as well as those from the western states of Gujarat and Rajasthan, indicate that trends towards size increase were underway before 2,000 BC36. The timing of size increase appears to finish somewhat earlier in the northwest and this distinct trend suggests that the domestication process was separate and may have begun earlier in northwestern India, perhaps from wild populations in the western Himalayas that were brought down to the Indo-Gangetic plains for cultivation. The current morphometric data on archaeological horsegram36 indicates that seed length and width are smaller in the earliest populations and appear to increase in both length and width around the middle of the Second Millennium BC, suggesting that domestication started by at least 4,000 years ago (2,000 BC) and size increase is evident by around 3,500 years ago (1,500 BC) and finishes by 3,000 years ago (1,000 BC). However, seed size change is sometimes regarded as an inaccurate indicator of domestication as this trait may evolve gradually, after, or take longer than, more essential domestication changes associated with seed shattering and germination5, 13, 37. There has been discussion as to whether or not size increase occurs during the initial stages of domestication or later during the process1, 38. Recent comparative studies suggest that in general seed size increase in pulses and other seed crops occurs during the same domestication episode that saw the evolution of other domestication traits4, 39.
Loss of germination inhibition, by contrast, is widely regarded as a key feature of domesticated legumes5, 8,9,10,11,12,13, 37. In wild pulses a thick and hard seed coat prevents germination, usually requiring physical damage to allow water to initiate germination. Studies of wild lentil and pea, for example, report germination rates of less than 10%8, 10, 12. This means that cultivation of wild-type legumes will offer very poor annual yields if plants are harvested whole and replanted annually. This has led to discussion of the possibility that rare, freely germinating mutants were selected from the wild8, rather than evolving under early domestication, and this process, should therefore be, a rapid, consciously-driven selection process12, 13. While lentil and pea have been the focus of most studies, and no field trials have been carried out specifically on wild horsegram, one expects parallel evolution in this trait across grain legume crops. However, documenting the evolution of this trait in archaeological seeds has been limited, due to the destruction of the seed coat in preserved archaeological specimens and because, when preserved, observation of the seed coat through conventional methods requires breaking and destroying archaeological specimens. We selected horsegram for this study as we have large assemblages of archaeological horsegram from Indian sites, many of which have wholly or partly preserved seed coats (Fig. 2a), which provided a good test case for methods that can be applied to legume seeds more widely.

(a) SEM of archaeological horsegram (from Sanganakallu) showing a radially fractured seed coat, (b,c) modern horsegram collected from Dharwad Market, India by author (DQF), a cross section showing cotyledons and seed coat, (d–f) examples of HRXCT images of archaeological horsegram including (d) Domesticated type from the site of Paithan dating to 0–400 AD (Lab code 70019) (e) Thicker wild type from the site of Hallur dating to 1,900 BC (Lab code 70018) (f) Semi-thin grade from the site of Sanganakallu dating to 1,400–1,250 BC (Lab code 70051).
Results
We conducted an experiment using high resolution x-ray computed tomography (HRXCT) to document preserved archaeological seed coats in their entirety and non-destructively. HRXCT and the use of synchrontrons for imaging biological and palaeobiological materials has proved useful in a wide range of studies40, 41. Our experiment is the first to try this method for documenting a domestication trait in archaeological seeds. A selection of twelve well-preserved, charred archaeological seeds were selected from four southern Indian sites, Paithan (Pa), Hallur (H), Sanganakallu (S), and Piklihal (Pi) (Fig. 1) representing periods between 2,000 BC to 500 AD (Neolithic to Early Historic periods), which provide a sequence that should capture much of the domestication process in this crop. These were examined at the UK National synchrotron. Three seeds from the early historic period, 0 to 500 AD, should represent fully domesticated forms after the evolutionary domestication process had finished15, and therefore represent a control for determining the amount of change that took place earlier. In addition, we measured seed length and width from 450 archaeological seeds from the same sites, combined with 36 published measurements in order to document size change over the same time scale and geography (Tables 1, S2 and S6). These data indicate a significant increase in the average and range of seed width between assemblage before and after ca. 1,700 BC (Fig. 3).

Change in seed size and seed coat thickness over time in Southern Indian archaeological horsegram. Top graph show mean and standard deviation of seed width (data: Table S2). Lower graph shows testa thickness mean and standard deviation (data: Table S3). Trend lines through Neolithic/Chalcolithic samples indicates the trend during the domestication process.
We were able to successfully, non-destructively image archaeological seeds allowing for the first time seed coats to be measured throughout the length and circumference of the seed. An additional result was that we were able to gauge the extent of internal tissue preservation within the seed (Fig. 2d–f)42. Our results so far indicate that there is considerable variation within individual archaeological specimens seed coat thickness (Mean values range from 23.2 to 10.6, with corresponding SD 13.8 and 2.8 and SEM 2.2 and 0.47) (Tables S3 and S4), and therefore single spot estimates of testa thickness may be misleading. Some of this variation may be due to differential shrinkage during charring on different parts of the same seed and post-charring damage leading to apparent thinning. This highlights the need for large numbers of testa thickness observations. Despite variation found across individual specimens, there is more variation that is statistically significant between specimens, and this variation indicates temporal trends over time (Figs 3, 5 and 6, Tables 2, 3 and S5).

Boxplot of archaeological Macrotyloma seed width divided into three periods (raw data in Table S6. X indicates the mean. Means between the early phase (before 1,700 BC) and second phase (1,700–1,200 BC) are significantly different (t-test for equal mean: p = 0.00059786). Kolmogorow-Smirnov test for equal distributions between these two phases is also significantly different (p = 0.0010299).

Boxplot of modern, uncharred testa thickness measured in 5 modern domesticated Macrotyloma uniflorum var. uniflorum populations. Raw data in Table S1.
The data indicates that we are able to document evidence for morphological evolution in terms of average testa thickness. Seeds can be grouped into two statistically significant testa size grades, thicker (wild type) seed coat that averages above >17 micrometres versus thin testa with averages between 15 and 10 micrometres. Differences between thick/intermediate and thin examples (thin: HLR98A4, SAN04 1130, PTN985, PTN985, PTN1036) are consistently significant to at least p = 0.18 and usually much higher levels (>×10−5) (Table S5). However, differences between intermediate and thick testa are only significant in pairwise comparisons between HLR98A-8 and PKL03B 130–160 (p = 0.019), or between PKL03B 130–160 and the other PKL specimens (p = 0.005 and p = 0.04). The whole dataset of thickness measurements is Log10 Normal, a common feature of biological and morphological data. Skewness is near 0 or positive with the exception of one slightly negative set (HLR98A-6). Thickness measurements do not fall below 6 micrometers (once charred), which can be regarded as the minimal testa thickness in this taxon. It also appears that the variability within a specimen tends to much greater in the thick and older, and hence more wild, examples, than amongst the intermediate or thinner specimens. This trend deserves further investigation on a larger number of specimens.
All of the thinner types are dated to 1,400 BC or more recent, while thicker testa grades are all older than 1,300 BC. Less statistically significant is an intermediate, or semi-thick grade (17–20 micrometers) versus a truly thick, wild type (>22 micrometers) (Fig. 6; Tables 3 and S5). Further samples should help to clarify how robust the thick versus intermediate distinction is. If these are considered, it can be seen that the thickest seed coats predate 1,700 BC, while the intermediate thicknesses have been found between 1,800 BC and 1,300 BC. This would suggest the hypothesis that selection for thinner seed coats, and by inference reduction of germination inhibition, was a stepped process. Of the four earliest specimens dating between 2,000–1,700 BC, three fall in the wild type grade and one in the semi-thin grade. Among the next five specimens dating between 1,700 and 1,250 BC, three are of the semi-thin grade and two of the thin/domesticated type. Finally, three early historic specimens, dating between AD 200 and 500 are all of the thin/domesticated grade. This indicates that the evolution of thinner seed coats took place during the Second Millennium BC and that seed coats were fixed in terms of thickness before the early centuries AD, although further research is needed to identify the morphological status of specimens between 1,250 and 1 BC. However, the directional trend through the Second Millennium BC indicates a rate of morphological change comparable to that estimated for seed coat thinning in North American archaeological Chenopodium 4, 43. Thus seed coat thinning evolved over a protracted period, as has been documented for other crop domestication traits.
Discussion
The different grades in testa thickness suggest that this trait is not controlled by a single underlying genetic factor but involves at least two sets of changes. This fits with observations in some other legumes, such as pea and hyacinth bean (Lablab purpureus) that show a spectrum of states from high dormancy to easily germinating10, 44. Therefore, the directional evolution of the seed coat took place under early cultivation alongside seed size increase (Figs 3 and 4), and this was not more rapid than other domestication traits that have been documented in grain crops4. In addition, it implies that cultivation and utilization of this pulse began despite wild dormancy, contrary to predictions proposed for other grain legumes8, 12, 13. Horsegram now provides a model for the archaeological documentation of grain legume domestication and affirms the utility of HRXCT for documenting seed coat thickness. This methodology has wider implications and utility for other domesticated crops as it can be applied to other archaeobotanical domesticates to non-destructively assess traits of domestication such as testa thickness and other morphometric properties connected with domestication. It is anticipated that this technique will propel forward research on the biogeographical origins, diversification and improvement of other pulses and domesticated crops and contribute to future issues of agricultural sustainability.

Boxplots of archaeological testa thickness measurements, colour-coded into suggested thick (wild), intermediate and thin (domesticated) testa grades. Boxed areas are the 25th and 75th percentile around the median. Outliers are indicated for measurements at more than 3 times the height of the box; all other measurements fall within the whiskers. Raw data in Table S4.
Methods
Archaeological seed specimens were individually mounted with clear nail varnish on the end of 18 mm cryo-pins (Moloceular Dimensions Limited, MD7-410) and placed in the beamline imaging chamber, Diamond Light Source beamline I-13-2, which is a 250 m long branch-line allowing relatively low energy and large field of view imaging. Samples were each run with 180° scan, with 4,000 projections at 0.15, 0.2 or 0.25 second exposure time each, with 10 darks and 10 flats, with an energy of 15 KeV. We found that 0.15 exposure was sufficient, but 0.2 second had improved resolution. Images were captured with the PCO 4000 detector and 4x objective. We also tried the 10x objective but found that 4x had sufficient magnification and better allowed for the whole seed to be captured.
Tomography was computed with the DAWN software run on the Diamond Light Source servers45, which produced between 2000 and 4000 thin section slices across each seed. Seed coat measurements were taken on a selection of 10 slices across the seed, evenly spaced, and 4 measurements were taken at approximately 30°, 70°, 120°, and 180° degrees around the seed circumference. Due to preservation differences between specimens some measurements are absent as part of the seed coat was missing. Measurements were taken with Leica software (LAS-EZ-V3-2-0) and were calibrated for each image based on the width to the cryo-pin.
Sites
All the synchrotron imaged archaeological horsegram chosen for this study were from South India. Specifically, Piklihal is a well-documented, multi-phase Southern Neolithic period site, consisting of a complex of occupation areas located in the Deccan plateau, with a well-established chronology, based upon radiocarbon dates dating from the Lower Neolithic to the Medieval period36. Sanganakallu is a Neolithic village settlement19, 34. Hallur is an early Chalcolithic site19, 26. Paithan is a much later, well-sampled Early Historic site46.
Dating
The date of specimens were inferred from their archaeological context. The Neolithic sites included in this study were all from South India and included Piklihal, Sanganakallu and Hallur which are all well dated by direct AMS dates on crop remains, coherent stratigraphic relationships and Bayesian statistical models of published absolute chronology19. Paithan is an Early Historic site in which the approximate age has been inferred from artefact types including coins.
Significance testing
Statistical significance of results of testa thickness measurements was performed in the software Past 3.11, 28 including both a one-way ANOVA that indicates the probability of equal means across all the samples is 1.7 × 10–35. Pairwise comparisons across the dataset were explored with the Tukey-Kramer test47, 48.
Future Research
Another round of beam time on beamline I-13 was awarded for May 2017 with Diamond Light Source. We will be targeting archaeological horsegram samples dating between 1,200 BC to 1 BC to fill in this lacuna and increase the sample size of archaeological horsegram testa measurements.
References
Purugganan, M. D. & Fuller, D. Q. The nature of selection during plant domestication. Nature 457, 843–848 (2009).
Larson, G. et al. Current perspectives and the future of domestication studies. PNAS 111, 6139–6146 (2014).
Tanno, K. & Willcox, G. How fast was wild wheat domesticated? Science 311, 1886 (2006).
Fuller, D. Q. et al. Convergent evolution and parallelism in plant domestication revealed by an expanding archaeological record. PNAS 111, 6147–6152 (2014).
Fuller, D. Q. & Allaby, R. Seed dispersal and crop domestication: shattering, germination and seasonality in evolution under cultivation. In Fruit Development and Seed Dispersal, Annual Plant Reviews, vol. 38, 238–295, Editor Lars Ostergaard (Wiley-Blackwell, Oxford, 2009).
Smith, M. L. How Ancient Agriculturalists Managed Yield Fluctuations through Crop Selection and Reliance on Wild Plants: An Example from Central India. Econ. Bot. 60, 39–48 (2006).
Bruno, M. C. A Morphological Approach to Documenting the Domestication of Chenopodium in the Andes. In Zeder, M. A., Bradley, D. G., Emshwiller, E. & Smith, B. D. (eds) Documenting Domestication: New Genetic and Archaeological Paradigms, 32–45 (University of California Press, Berkeley and Los Angeles, 2006).
Ladizinsky, G. Pulse domestication before cultivation. Econ. Bot. 41, 60–65 (1987).
Butler, A. Cryptic anatomical characters as evidence of early cultivation in the grain legumes (pulses). In Foraging and farming: the evolution of plant exploitation, 390–407 (Unwin Hyman, London, 1989).
Weeden, N. F. Genetic Changes Accompanying the Domestication of Pisum sativum: Is there a Common Genetic Basis to the ‘Domestication Syndrome’ for Legumes? Ann. Bot. 100, 1017–1025 (2007).
Smartt, J. Grain Legumes (Cambridge University Press, Cambridge, 1990).
Abbo, S., Rachamin, E. & Zehavi, Y. Experimental growing of wild pea in Israel and its bearing on Near Eastern plant domestication. Ann. Bot. 107, 1399–1404 (2011).
Aboo, S., Pinhasi van Oss, R. & Gopher, A. Plant domestication versus crop evolution: a conceptual framework for cereals and grain legumes. Trends Plant Sci. 19, 351–360 (2013).
Hooker, J. The Flora of British India, vol. 2 (L. Reeve & Co., London, 1879).
Fuller, D. Q. & Harvey, E. L. The archaeobotany of Indian pulses: identification, processing and evidence for cultivation. Environmental Archaeology 11, 219–246 (2006).
Kadam, S. S. & Salunkhe, D. K. Nutritional composition, processing, and utilization of horse gram and moth bean. Crit. Rev. Food Sci. Nutr. 22, 1–26 (1985).
Kachroo, P. & Arif, M. (eds) Pulse crops of India (Indian Council of Agricultural Research, New Delhi, 1970).
Fuller, D. Q. Fifty Years of Archaeobotanical Studies in India: Laying a Solid Foundation. In Indian Archaeology in Retrospect, vol. III of Archaeology and Interactive Disciplines, 247–364, Publications of the Indian Council for Historical Research (Manohar, New Dehli, 2002).
Fuller, D. Q., Boivin, N. & Korisettar, R. Dating the Neolithic of South India: New radiometric evidence for key economic, social and ritual transformations. Antiquity 81, 755–778 (2007).
Bellwood, P. The First Farmers (Blackwell, Oxford, 2005).
Balter, M. Seeking Agriculture’s Ancient Roots. Science 316, 1830–1835 (2007).
Fuller, D. Q. & Murphy, C. Overlooked but not forgotten: India as a center for agricultural domestication. General Anthropology 21, 1–8 (2014).
Kingwell-Banham, E. & Fuller, D. Q. Brown Top Millet: Origins and Development (2014).
Fuller, D. Q. & Harvey, E. L. The Archaeobotany of Indian Pulses: identification, processing and evidence for cultivation. Environmental Archaeology 11, 219–246 (2006).
Dhavalikar, M., Sankalia, H. & Ansari, Z. Excavations at Inamgaon. I (Parts i and ii). Tech. Rep., Deccan College, Pune (1988).
Fuller, D. Q., Korisettar, R., Venkatasu, P. & Jones, M. K. Early plant domestications in southern India: some preliminary archaeobotanical results. Veg. Hist. Archaeobot. 13, 115–129 (2004).
Asouti, E. et al. Early Holocene woodland vegetation and human impacts in the arid zone of the southern Levant. The Holocene 25, 1124–1139 (2015).
Murphy, C. & Fuller, D. Q. The Transition to Agricultural Production in India: South Asian Entanglements of Domestication. In Schug, G. R. & Walimbe, S. R. (eds) A Companion to South Asia in the Past, 344–357 (John Wiley & Sons, Inc., Hoboken, NJ, 2016).
Kajale, M. Current status of Indian palaeoethnobotany: introduced and indigenous food plants with a discussion of the historical and evolutionary development of Indian agriculture and agricultural systems in general. In New light on early farming- recent developments in palaeoethnobotany, 155–189 (Edinbugh University Press, Edinburgh, 1991).
Saraswat, K. Plant economy of Late Harappan at Hulas. Puratattva 23, 1–12 (1992).
Willcox, G. Some differences between crops of Near Eastern origin and those from the tropics. In South Asian Archaeology 1989, vol. 14 of Monographs in World Archaeology, 291–299 (Prehistory Press, Madison, 1992).
Fuller, D. Q., Willcox, G. & Allaby, R. G. Cultivation and domestication had multiple origins: arguments against the core area hypothesis for the origins of agriculture in the Near East. World Archaeol. 43, 628–652 (2011).
Kajale, M. D. Initial palaeoethnobotanical results from Neolithic Watgal, South India in relation to data from contemporary sites (1998).
Roberts, P. et al. Local diversity in settlement, demography and subsistence across the southern Indian Neolithic-Iron Age transition: site growth and abandonment at Sanganakallu-Kupgal. Archaeol. Anthropol. Sci. 8, 575–599 (2016).
Fuller, D. Q. Finding Plant Domestication in the Indian Subcontinent. Curr. Anthropol. 52, S347–S362 (2011).
Fuller, D. Q. & Murphy, C. The origins and early dispersal of horsegram (Macrotyloma uniflorum), a major crop of ancient India. Genet. Resour. Crop Evol. 1–21 (2017).
Sonnante, G., Hammer, K. & Pignone, D. From the cradle of agriculture a handful of lentils: history of domestication. Rend. Lincei. 20, 21–37 (2009).
Purugganan, M. & Fuller, D. Q. Archaeological data reveal slow rates of evolution during plant Domestication. Evolution 65, 171–183 (2010).
Moles, A. T. et al. Global patterns in seed size. Global Ecol. Biogeogr. 16, 109–116 (2007).
DeVore, M., Kenrick, P. & Pigg, K. Utility of high resolution X-ray computed tomography (HRXCT) for paleobotanical studies: an example using London Clay fruits and seeds. Am. J. Botany 93, 1848–1851 (2006).
Bertrand, L., Cotte, M. & Stampanoni. Development and trends in synchrotron studies of ancient and historical materials. Phys. Rep. 519, 51–96 (2012).
Zong, Y. et al. Selection for Oil Content During Soybean Domestication Revealed by X-Ray Tomography of Ancient Beans. Sci. Rep. 7 (2017).
Smith, B. Eastern North America as an independent center of plant domestication. PNAS 103, 12223–12228 (2006).
Maass, B. Changes in seed morphology, dormancy and germination from wild to cultivated hyacinth bean germplasm (Lablab purpureus: Papilionoideae). Genet. Resour. Crop Evol. 53, 1127–1135 (2005).
Basham, M. et al. Data Analysis WorkbeNch (DAWN). J. Synchrotron Radiat. 22 (Pt3), 853–858 (2015).
Fuller, D. Archaeobotany. In Transformations in Early Historic and Early Medieval India: Excavations at Paithan, Maharashtra 1996–1999, 289–327 (Archaeological Survey of India, New Delhi, 2013).
Hammer, O., Harper, D. & Ryan, P. PAST: Paleontological Statistics software package for education and data analysis. Palaeontol. Electronica 4, 1–9 (2001).
Tukey, J. Comparing Individual Means in the Analysis of Variance. Biometrics 5, 99–144 (1949).
Garcia-Graner, J. J. From Gathering to farming in semi-arid Northern Gujarat (India): A multi-proxy Approach. PhD, Universitat de Barcelona, Barcelona (2015).
Weber, S. A. Plants and Harappan subsistence: an example of stability and change from Rojdi (Westview Press, Boulder, 1991).
Vishnu-Mittre. Discussion. In Early history of agriculture, vol. B275 of Philos. Trans. R. Soc. Lond., B, Biol. Sci. 141 (1976).
Kajale, M. New light on agricultural plant economy during 1st millennium BC: Palaeobotanical study of plant remains from excavations at Veerapuram, District Kurnool, Andhra Pradesh. In Veerapuram: a type site for cultural study in the Krishna valley, vol. Appendix B, 15 (Birla Archaeological and Cultural Research Institute, Hyderabad, 1984).
Vishnu-Mittre. Palaeobotanical evidence from India. In Evolutionary studies in world crops: diversity and change in the Indian subcontinent, 3–30 (Cambridge University Press, Cambridge, 1974).
Acknowledgements
This research is part of the Comparative Pathways to Agriculture Project (ComPAg) funded by the European Research Council (grant 323842). Synchrotron imaging was supported by a grant of beam time from Diamond Light Source (www.diamond.ac.uk), project MT12082. We acknowledge Diamond Light Source for time on Beamline I-13 under Proposal MT12082. We thank Beamline Scientist Dr. Christoph Rau and staff at Branchline I-13 for their assistance. For modern reference materials we thank the Royal Botanic Gardens, Kew; the London Natural History Museum herbarium department, and USDA National germplasm collections.
Author information
Authors and Affiliations
Contributions
D.Q.F. and C.M. undertook all experimental work and wrote the main manuscript text. D.Q.F. prepared Figures 1 and 3. C.M. prepared Figure 2. Both authors reviewed the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Murphy, C., Fuller, D.Q. Seed coat thinning during horsegram (Macrotyloma uniflorum) domestication documented through synchrotron tomography of archaeological seeds. Sci Rep 7, 5369 (2017). https://doi.org/10.1038/s41598-017-05244-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-05244-w
This article is cited by
-
Applications of Microct Imaging to Archaeobotanical Research
Journal of Archaeological Method and Theory (2024)
-
Synchrotron tomography applications in agriculture and food sciences research: a review
Plant Methods (2022)
-
Monsoon forced evolution of savanna and the spread of agro-pastoralism in peninsular India
Scientific Reports (2021)
-
A simple and efficient Agrobacterium-mediated in planta transformation protocol for horse gram (Macrotyloma uniflorum Lam. Verdc.)
Journal of Genetic Engineering and Biotechnology (2020)
-
A correlation analysis of Light Microscopy and X-ray MicroCT imaging methods applied to archaeological plant remains’ morphological attributes visualization
Scientific Reports (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
