
Abstract
We used a high-resolution oxygen isotope (δ18Ocoral), carbon isotope (δ13Ccoral) and Sr/Ca ratios measured in the skeleton of a reef-building coral, Porites sp., to reveal seasonal-scale upwelling events and their interannual variability in the Gulf of Oman. Our δ13Ccoral record shows sharp negative excursions in the summer, which correlate with known upwelling events. Using δ13Ccoral anomalies as a proxy for upwelling, we found 17 summer upwelling events occurred in the last 26 years. These anomalous negative excursions of δ13Ccoral result from upwelled water depleted in 13C (dissolved inorganic carbon) and decreased water-column transparency. We reconstructed biweekly SSTs from coral Sr/Ca ratios and the oxygen isotopic composition of seawater (δ18OSW) by subtracting the reconstructed Sr/Ca-SST from δ18Ocoral. Significant δ18OSW anomalies occur during major upwelling events. Our results suggest δ13Ccoral anomalies can be used as a proxy for seasonal upwelling intensity in the Gulf of Oman, which, driven by the Indian/Arabian Summer Monsoon, is subject to interannual variability.
Similar content being viewed by others

Introduction
The Gulf of Oman is located on the northeastern coast of the Arabian Peninsula and both the Arabian Sea and the Gulf of Oman are located in arid environments. The climate is dominated by the seasonal reversal of the Indian/Arabian Monsoon, which in turn governs the surface wind field of the Indian Ocean north of 10°S. The intensity and direction of the monsoon winds vary seasonally. During the southwest (SW) Monsoon develops during the boreal summer (from June to mid-September) and is characterized by strong airflow across the Arabian Sea that feeds moisture and rainfall to the Indian subcontinent.
The Indian/Arabian Summer Monsoon causes coastal upwelling bringing cooler temperatures, nitrified and saline water to the sea surface along the southern coast of the Arabian Peninsula. Upwelled water flows northward and affects the oceanic stratification of the Gulf of Oman through gyres and eddy systems that sweep into the Oman Sea1. The Northern Arabian Sea is therefore one of the most productive areas in the world2. The SW Monsoon is also the major climatic factor affecting the near-shore environment and areas of coral growth in Oman during the summer months3.
The high nutrient content of this water induces phytoplankton blooms. Satellite-based ocean color measurements show the temporal and spatial variability of the surface chlorophyll-a distribution along the coast of the Southern Arabian Peninsula4. In the Gulf of Oman, upwelling does not necessarily occur every summer5,6,7. In addition, observational records that allow us to understand the dynamics of upwelling events in the Gulf of Oman are scarce. Satellite based sea surface temperature (SST) in the Gulf of Oman did not reflect low SST excursions in summer measured by CTDs. (Fig. S1). Long-term and in situ records of primary production, salinity and temperature are necessary in order to understand upwelling events7. In this study, we used paleo-climatic reconstructions from coral geochemical records to provide a history of summer monsoon-driven upwelling variability in the Gulf of Oman.
The geochemical proxies in coral skeletal carbonate provide a long-term history of environmental variation, with high time resolution (2 weeks to a month)8, 9. Coral skeletal oxygen isotopes (δ18Ocoral) reflect SST and oxygen isotopes in seawater (δ18OSW)10, 11. Coral skeletal Sr/Ca ratios are used as SST proxies12. Sea-surface salinity (SSS) is derived from δ18OSW, which is generated by subtracting the temperature component (obtained from coral Sr/Ca) from δ18Ocoral 12. Stable isotopes of carbon in coral skeletons (δ13Ccoral) are influenced by kinetic isotopic fractionation, vital effects (photosynthesis and respiration) and the carbon isotopic composition of dissolved inorganic carbon in seawater (δ13CDIC-SW)13,14,15,16. Coral skeletons are precipitated in isotopic disequilibrium with ambient seawater as a result of kinetic and vital effects. The kinetic effect selectively depletes 12C and 16O in coral skeletons and is particularly important when coral growth rates are very low (<4 mm per year)13, 14. Photosynthetic activities of zooxanthellae affect δ13Ccoral by changing the carbon isotopes in the internal dissolved inorganic carbon pool of the coral17 . A 50% weakening of solar radiation induces a decrease of approximately 0.5‰VPDB in δ13Ccoral 18. The amount of solar radiation received by the coral varies depending on incoming solar radiation, cloud cover and water transparency17, 19, 20. Upwelling can reduce water transparency and change the sea-surface δ13CDIC-SW. Therefore, upwelling events should be registered by the coral via a decrease in δ13Ccoral. We used a coral record from the Gulf of Oman to reconstruct the timing and frequency of upwelling events using high-resolution records of Sr/Ca ratios, δ18OSW and δ13Ccoral based on a 26-year-old coral core.
Results and Discussion
We determined δ18Ocoral, δ13Ccoral and Sr/Ca ratios from 664 samples. Each powdered sample was split for paired stable isotope and Sr/Ca analysis. Sr/Ca ratios and δ18Ocoral showed 26 distinct annual cycles (Fig. 1a and b). The average of the Sr/Ca ratios was 9.28 (mmol × mol−1), with values ranging from 8.98 to 9.56 (mmol × mol−1). The δ18Ocoral averaged −4.33 (‰VPDB) and ranged from −4.92 to −3.41 (‰VPDB). We calculated the regression line between satellite SST and Oman coral Sr/Ca ratios using seasonal maxima and minima to avoid potential biases due to intra-seasonal age model uncertainties, as follows:

Oman coral proxy records and extension rate. (a) Coral skeletal δ18OVPDB record, (b) Coral skeletal Sr/Ca ratio record. Grey arrows indicate the years of non-increasing Sr/Ca ratios in summer. (c) δ18OSW-anomaly, (d) Coral skeletal δ13CVPDB record, (e) Extension rate calculated from distances between the anchor points in winter of each year. (f) Oman coral skeletal δ13CVPDB anomaly. The timing of anomalous negative excursions of AN-δ13C in the summer are shown as black arrows. (g) In situ data showing low-SST and high chlorophyll-a (square symbols).
We established a regression line between satellite SST and δ18Ocoral, assuming that δ18Ocoral reflect only SST variations, with the same δ18Ocoral samples with Sr/Ca ratios, as follows:
The correlation coefficient between δ18Ocoral and Sr/Ca ratios was 0.77 (P < 0.01). δ18Osw were calculated by subtracting the temperature component (estimated from coral Sr/Ca ratios) from δ18Ocoral, following the method proposed by Nurhati et al.21. The slope of the δ18Ocoral -SST regression is −0.104 ± 0.005‰VPDB/°C, which is too high to be consistent with published estimates12, 13, 22. This suggests a significant contribution of δ18OSW to δ18Ocoral. We therefore used the published regression slope of −0.18 ± 0.03 (‰/°C)12 to convert δ18Ocoral to SST, and our slope of −0.044 mmol × mol−1/°C for SST estimation. The δ18OSW anomalies were calculated by applying a band-pass filter to remove the periodicity components longer than 2 years and subtracting the seasonal cycle. Relative changes of δ18OSW are on the order of ± 0.424‰VSMOW (2σ). Anomalies above or below this threshold were marked as significant δ18OSW anomalies (Fig. 1c). The uncertainty of calculated δ18OSW is ± 0.113‰VSMOW (following Nurhati et al.21).
The average δ13Ccoral was −1.62 (‰VPDB) and ranged from −3.28 to +0.29 (‰VPDB). The δ13Ccoral also showed clear seasonal variation (Fig. 1d) and distinct short-term negative anomalies (Fig. 1d). The δ13Ccoral analysis was performed to avoid contamination from organic matter. We measured each CO2 gas sample 6 times using a dual inlet system loaded on a MAT251. Analytical precision of the δ13Ccoral (standard deviations) were below 0.05‰. Growth rate disturbances and anomalous-colored annual band were not observed on X-ray photographs and coral cores. Therefore, the variations of δ13Ccoral were assumed to reflect environmental changes rather than the coral growth disturbances23.
The interpretations of δ13Ccoral has been debated about what the δ13Ccoral values are reflecting10, 13,14,15,16, 24, 25. The main factors influence that can influence δ13Ccoral include: (1) kinetic effect and vital effect, (2) solar radiation, (3) water-column transparency, (4) variation of δ13CDIC-SW and (5) autotroph/heterotroph ratios.
Kinetic effects have been recognized as simultaneous 18O and 13C enrichment in coral skeletons with low extension rates13. Strong kinetic effects mask vital effects13. In our core, δ13Ccoral values showed a weak negative correlation with the δ18Ocoral record (r = −0.317, n = 634, P < 0.001: Fig. S2a). Summer δ13Ccoral did not correlate significantly with δ18Ocoral (r = 0.140, n = 181, P > 0.05: Fig. S2a). Winter δ13Ccoral had no significant correlation with winter δ18Ocoral (r = 0.04, P > 0.05, n = 159: Fig. S2b). The extension rates show that the Oman coral grew very quickly, on average 25.1 mm/year with a range between 19 to 31.5 mm. These values were considerably higher than the critical value estimated for kinetic isotopic fractionation effects (4 mm/year) (Fig. 1f)14. Therefore, the coral growth history and the lack of correlation between δ13Ccoral and δ18Ocoral suggest that the kinetic isotopic effect did not significantly affect this coral record.
Previous studies reported δ13Ccoral on seasonal and inter-annual variations are attributable to solar radiation17, 26. To investigate the processes driving these δ13Ccoral fluctuations, we compared δ13Ccoral with satellite-based outgoing longwave radiation (OLR) (Fig. S3a) which reflect cloud cover. For a comparison of δ13Ccoral with monthly-resolved OLR data, biweekly resolved δ13Ccoral data were resampled at a monthly resolution using the software AnalySeries (version 2.0.8)27. The δ13Ccoral were compared with OLR, and we calculated the correlation coefficients between these time series. δ13Ccoral without anomalous δ13Ccoral excursions positively correlated with OLR at a significant level (r = 0.411, P < 0.01, n = 302: Fig. S3a and S3b). A significant correlation appeared between the mean seasonal cycle of δ13Ccoral and OLR averaged over the past 26 years (r = 0.702, P = 0.01, n = 12: Fig. S3c and d). The positive correlations between δ13Ccoral and OLR (Fig. S2b and S2d) suggest that δ13Ccoral captured the variation of photosynthetic activity caused by the seasonal solar radiation cycle. At inter-annual resolution, the 15 month-moving average profile of δ13Ccoral positively correlate with that of OLR (r = 0.347, P < 0.01, n = 303: Fig. S4a and S4b). The duration of low OLR and coeval δ13Ccoral decreased from 1992 to 1993. We propose that insolation and OLR had decreased in globally as a result of up-stirred volcanic aerosol from the eruption of Mount Pinatubo, the Philippines in June 199128. Low δ13Ccoral from 1992 to 1993 would be influenced by decreasing insolation which resulted from the volcanic eruption of Mount Pinatubo.
We calculated the δ13Ccoral anomaly (δ13Canomaly) by removing the 15 month-moving average (31 bi-weekly data point) after subtracting the averaged seasonal cycle of δ13Ccoral. The threshold for δ13Ccoral anomalous excursions was determined as a standard deviation of 1σ: ± 0.343‰VPDB. In summer, the anomalous negative excursions of the δ13Canomaly occurred 17 times in summer, while 1 anomalous negative excursion occurred in the spring of 1993 (Fig. 1f). Anomalous positive δ13Canomaly excursions were also observed prior to summer negative δ13Canomaly excursions. The δ13Canomaly had no significant correlation with OLR anomaly calculated by same procedure (r = 0.05, P > 0.3 Fig. S4c and S4d), suggesting that anomalous negative excursions of δ13Canomaly in the summer (AN-δ13C) would not be generated from OLR variations.
We examined the timing of the AN-δ13C with the compiled evidence of each past upwelling event documented from in situ and satellite observations (Fig. 1f and g). Abrupt SST decreasing events in summer were revealed in 1987–198929 and 200030 from satellite SST data, in 19926, 19944, 200130, 200230 (Fig. S5) and 199031 based on in situ SST data, and 2010 based on our vertical seawater temperature profile (Fig. S6). The vertical profile of seawater temperature deduced by temperature sensors attached to the diving gear of local volunteer divers in 2010, also suggest that the thermocline was closer to the surface during summer upwelling events (Fig. S6). In addition, Al-Azri et al.1 had measured chlorophyll-a concentrations, nutrients, phytoplankton density and SST in Fahal Island (23.67°N, 58.5°E) and Bandar Al Khayran (23.51°N, 58.72°E: near to our coral sample site). From July to September 2004, upwelling was observed as increasing chlorophyll-a concentrations and phytoplankton density as well as decreasing SST1. In August 2005, SST decreased for 1 month, while other parameters did not change1. Al-Azri et al., 2012 reported that in situ chlorophyll-a and satellite based SST suggested that upwelling also occurred in July, 20087. The satellite observations (SeaWiFS and MODIS at 24°N, 58°E from Asia-Pacific Data Research Center32) from 1997 to 2013 suggested that chlorophyll-a concentrations in the Gulf of Oman increased in August 2000, September 2004 and August 2008 (Fig. S7). In other upwelling years, chlorophyll-a concentrations in satellite data were not available to compare with AN-δ13C due to the lack of satellite data in summer. Based on these in situ and satellite datasets, past upwelling events occurred in 198729, 198829, 198929, 199031, 19926, 19946, 2000–200230, 20041, 20087 and 2010 (Fig. 1g). The AN-δ13C corresponds with these past upwelling events.
The possible controlling factors of the AN-δ13C with upwelling events are: (1) decreasing water-column transparency33, (2) variations of δ13CDIC-SW 16, 34, and (3) change to heterotroph feeding10. It is known that increasing chlorophyll-a concentrations correspond with upwelling events1, 7 inducing phytoplankton blooms, thereby decreasing water-column transparency and depleting 13Ccoral with low photosynthetic activities of zooxanthellae33. Moreover, lower δ13CDIC-SW supply from greater depths decreases δ13CDIC-SW at the sea surface24, 25, 34. Upwelling events may produce an AN-δ13C due to sudden decreases in water-column transparency and δ13CDIC-SW. Heterotrophic feeding would also be the controlling factor of negative δ13C coral with upwelling events. A study35 reported that corals feeding 13C-depleted zooplankton decreased their δ13Ccoral. The coral records from the Gulf of Aqaba, Red Sea suggested that increasing heterotrophy with upwelling decreased δ13Ccoral for an approximately half a year10. Afterwards, δ13Ccoral could be increased by the preferential uptake of 12C by phytoplankton at the sea surface16. In the western Indonesian coast, it was reported that δ13Ccoral increased by approximately 2.2‰VPDB after large phytoplankton blooms due to upwelling16.
We propose the following mechanism to explain the short-term negative peaks in the δ13Ccoral: 1. Upwelling events bring deep, cold and nutrient-rich water with low δ13CDIC-SW to the surface in summer. Upwelling events cause unusually high nutrient conditions in the Gulf of Oman. Photosynthesis activities in zooxanthella would be emphasized in eutrophic conditions and temporarily increased δ13Ccoral. 2. Lower δ13CDIC-SW from the deep sea decreases δ13Ccoral. 3. Phytoplankton blooms arise from a nutrient supply to the sea surface. 4. Phytoplankton primarily depletes 12CO2-SW. Active phytoplankton photosynthesis increases 13CO2-SW. 5. δ13Ccoral increases with the restoration of δ13CDIC-SW.
We compared the AN-δ13C minima with the upwelling periods (number of the days) in summer (Figs 2a, S5 and S6). In situ daily to weekly SST data in 1992, 1994, 2001, 2002 and 2010 revealed that SST during upwelling events was as same as winter SST (23.5 °C), and daily fluctuations of SST in upwelling periods ranged within 3 °C. Therefore, the numbers of the days for upwelling periods were defined as the duration of SST lower than 26.5 °C in summer. δ13Canomaly values of no upwelling years (0 days) was estimated from in situ δ13CDIC-SW in Arabian Sea (+1.325‰VPDB at 0–10 m depth in non-upwelling seasons36) and the value of δ13C in isotopic equilibrium between coral carbonate and seawater37. The AN-δ13C minima were correlated to the upwelling periods as below.

(a) Cross-plot of in situ low-SST periods and AN-δ13C minima. The dotted line indicates the regression between in situ low-SST periods and AN-δ13C minima (filled circles) and estimated δ13Canomaly value for no upwelling periods (r = −0.937, P < 0.05, n = 6). The δ13Canomaly value for no upwelling (open circle) was estimated from in situ δ13CDIC-SW from the Arabian Sea36 and the value of δ13C in isotopic equilibrium between coral carbonate and seawater37 (b) Estimated upwelling periods from AN-δ13C minima. Black bars indicate the years which were used for the regression between in situ low-SST periods and δ13Canomaly minima.
Upwelling periods (days) = −87.16 ± 16.40 × AN-δ13C minima (‰VPDB) − 4.92 ± 9.45 (r = −0.937, P < 0.05; Fig. 2a).
Then, past upwelling periods in the year with no in situ SST data were reconstructed from each AN-δ13C using this equation (Fig. 2b). The estimated uncertainty for reconstructed upwelling-periods was 12.66 days (1σ) including the analytical precisions of δ13Ccoral, the intercept and the slope of this equation. In 1987, 2006, 2008, 2009, each upwelling period was extremely long, over 120 days (Fig. 2b). In those years, coral extension rates decreased to 23 mm/year (Fig. 1e). The long upwelling events would therefore have a negative effect on coral extension rate due to eutrophic conditions and decreased water-column transparency.
We compared the reconstructed upwelling events from AN-δ13C (Fig. 2b) with Sr/Ca ratios and δ18OSW-anomaly (Fig. 1b and c). Sr/Ca ratios showed 1-month increasing (cooling) in summer except in 1994, 2001, 2002, 2006, and 2009, however, these did not correspond to reconstructed upwelling events. In non-AN-δ13C (upwelling) years (1989, 1991, 1997–1998, 2003, 2007, 2011–2012), the δ18OSW-anomaly was low in summer. Upwelling events in the Gulf of Oman are driven by the SW Monsoon, which causes strong seasonal winds parallel to the coast of Southern Oman in the Arabian Sea, while the associated Ekman transport creates strong upwelling along the coastal margins, bringing cold, nutrient-rich water to the surface1, 4. This upwelled water has indirect impacts on corals and reef areas farther north through gyres and eddy systems that sweep into the Oman Sea1, 4. In addition, upwelling may be influenced by vertical seawater density, depending on SST and SSS38. The δ18OSW-anomaly record suggested that deep seawater did not reach the sea surface as low-density water masses might form a cap on the sea surface in the Gulf of Oman.
Observations suggest that the primary productivity of the Gulf of Oman is subject to inter-annual variability1, but long-term observational records are lacking. Our new δ13Ccoral record captured past upwelling events and their periods in the Gulf of Oman for 26 years. Thus, coral skeletal archives fill an important gap in the observational record and have great potential for increasing our understanding of the upwelling mechanisms in the Gulf of Oman. Moreover, it is possible to reconstruct past SST, SSS and upwelling frequency/intensity during the Holocene (from 0 to 10 ka) by applying the same methods to fossil corals from the Arabian Peninsula.
Methods
Coral sampling
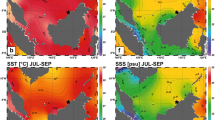
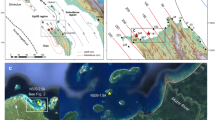
On February 23, 2013, we drilled a Porites sp. coral colony in the Gulf of Oman (23°30′ N, 58°45′ E: Fig. 3a and b). This Porites colony was living at a 2 m water depth in a small bay (Bandar Khayran) south of Muscat. There was no dry-riverbed (locally name: wadi) nearby; thus, we excluded the influence of occasional plumes of freshwater from coastal runoff at the site. In total, the coral core was 71 cm long. On the sampling date, we measured in situ SST and SSS at 24.3 °C and 38.2 PSU (practical salinity unit). Meteorological records from the weather station at Seeb Airport (23.60°N, 58.30°E) showed low precipitation rates, with less than 14.0 mm/month (the monthly average precipitation climatology for the past 23 years was 0.28–14.0 mm/month; GHCN-Monthly ver. 2). For coral proxy calibration, we used Advanced Very High Resolution Radiometer (AVHRR) satellite SST data, SODA satellite SSS data (http://iri.columbia.edu: Fig. 3c) and OLR data (https://climexp.knmi.nl/: Fig. 3c)39,40,41. Salinity records decrease in summer suggesting a possible occurrence of upwelling events.

(a),(b) Map of the sampling site in the Gulf of Oman. The figures were generated using Generic Mapping Tools (GMT ver. 4.5.12)44. (c) Climatological data estimated from biweekly SST from 1987 to 2013 (data from AVHRR39), monthly SSS from 1987 to 2008 (data from SODA40) and OLR41 during past 26 years (data from https://climexp.knmi.nl/). Error bars indicate climatology deviation (1σ).
Subsampling
The coral core was sliced into 5-mm-thick slabs. We took X-radiographs of the coral slabs to identify the coral growth axis (Fig. 4). We prepared ledges of 1.5 mm in thickness along the maximum growth axis and obtained coral powder at a resolution of 0.5 mm for geochemical analysis.

X-radiograph of coral core OMN130221 (Porites sp.). The white line indicates the measurement lines. Several overlapping measurement lines were sampled to ensuring reproducibility.
Oxygen and carbon isotope measurements
The coral powder was weighed, and 100 μg (±20 μg) were taken for oxygen and carbon stable isotope analysis. The sample powder was reacted with 100% H3PO4 at 70 °C in an automated carbonate preparation device (Kiel II). The δ13Ccoral and δ18Ocoral were analyzed with a Finnigan MAT251 stable isotope ratio mass spectrometer system installed at Hokkaido University. Analytical errors for δ13Ccoral and δ18Ocoral were determined to be 0.08 and 0.07‰, respectively, based on replicate measurements of the NBS-19 standard (1σ, n = 40).
Trace element measurements
We measured Sr/Ca ratios with a SPECTRO CIROS CCD SOP inductively coupled plasma optical emission spectrophotometer installed at Kiel University following a combination of methods described by Schrag42 and de Villiers et al.43. Approximately 250 μg of coral powder was dissolved in 4 mL of HNO3. The sample solution for the measurement of trace elements was prepared via serial dilution with 2% HNO3 for a Ca concentration of ca. 8 ppm. Analytical precision of the Sr/Ca determinations was 0.07% RSD or 0.01 mmol × mol−1 (1σ).
Data analysis
We used the coral Sr/Ca ratios to develop an age model for all proxies. Minima and maxima of the coral Sr/Ca ratios were chosen as anchor points and tied to the maxima and minima of SST, respectively. To obtain a time series with equidistant time steps, we resampled the proxy data at a biweekly resolution using the AnalySeries software, version 2.0.827. Annual extension rates were estimated from the distance (in mm) between the winter anchor points in each sclerochronological year.
References
Al-Azri, A. R., Piontkovski, S. A., Al-Hashmi, K. A., Goes, J. I. & Do Gomes, H. R. Chlorophyll a as a measure of seasonal coupling between phytoplankton and the monsoon periods in the Gulf of Oman. Aquat. Ecol. 44, 449–461 (2010).
Qasim, S. Z. Oceanography of the northern Arabian Sea. Deep Sea Res. Part A. Oceanogr. Res. Pap. 29, 1041–1068 (1982).
Burt, J. A. et al. Oman’s coral reefs: A unique ecosystem challenged by natural and man-related stresses and in need of conservation. Mar. Pollut. Bull. 105, 498–506 (2016).
Wiggert, J. D., Hood, R. R., Banse, K. & Kindle, J. C. Monsoon-driven biogeochemical processes in the Arabian Sea. Prog. Oceanogr. 65, 176–213 (2005).
Elliott, A. J. & Savidge, G. Some features of the upwelling off Oman. J. Mar. Res. 48, 319–333 (1990).
Coles, S. L. Reef corals occurring in a highly fluctuating temperature environment at Fahal Island, Gulf of Oman (Indian Ocean). Coral Reefs 16, 269–272 (1997).
Al-Azri, A. R. et al. Mesoscale and Nutrient Conditions Associated with the Massive 2008 Cochlodinium polykrikoides Bloom in the Sea of Oman/Arabian Gulf. Estuaries and Coasts 37, 325–338 (2013).
Cobb, K. M., Charles, C. D., Cheng, H. & Edwards, R. L. El Niño/Southern Oscillation and tropical Pacific climate during the last millennium. Nature 424, 271–276 (2003).
Watanabe, T. et al. Permanent El Niño during the Pliocene warm period not supported by coral evidence. Nature 471, 209–211 (2011).
Felis, T., Pätzold, J., Loya, Y. & Wefer, G. Vertical water mass mixing and plankton blooms recorded in skeletal stable carbon isotopes of a Red Sea coral. J. Geophys. Res. 103, 30731 (1998).
Tudhope, A. W., Lea, D. W., Shimmield, G. B., Chilcott, C. P. & Head, S. Monsoon Climate and Arabian Sea Coastal Upwelling Recorded in Massive Corals from Southern Oman. Palaios 11, 347 (1996).
Gagan, M. K. et al. Temperature and Surface-Ocean Water Balance of the Mid-Holocene Tropical Western. Pacific. Science 279, 1014–1018 (1998).
McConnaughey, T. 13C and 18O isotopic disequilibrium in biological carbonates: I. Patterns. Geochim. Cosmochim. Acta 53, 151–162 (1988).
McConnaughey, T. 13C and 18O isotopic disequilibrium in biological carbonates: II. In vitro simulation of kinetic isotope effects. Geochim. Cosmochim. Acta 53, 163–171 (1989).
Swart, P. K., Leder, J. J., Szmant, A. M. & Dodge, R. E. The origin of variations in the isotopic record of scleractinian corals: II. Carbon. Geochim. Cosmochim. Acta 60, 2871–2885 (1996).
Abram, N. J., Gagan, M. K., McCulloch, M. T., Chappell, J. & Hantoro, W. S. Coral reef death during the 1997 Indian Ocean Dipole linked to Indonesian wildfires. Science 301, 952–955 (2003).
Fairbanks, R. G. & Dodge, R. E. Annual periodicity of the 18O/16O and 13C/12C ratios in the coral Montastrea annularis. Geochim. Cosmochim. Acta 43, 1009–1020 (1979).
Grottoli, A. Effect of light and brine shrimp on skeletal δ13C in the Hawaiian coral Porites compressa: a tank experiment. Geochim. Cosmochim. Acta 66, 1955–1967 (2002).
Pätzold, J. Growth rhythms recorded in stable isotopes and density bands in the reef coral Porites lobata (Cebu, Philippines). Coral Reefs 3, 87–90 (1984).
Wellington, G. M. & Dunbar, R. B. Stable isotopic signature of El Niño Southern Oscillation events in eastern tropical pacific reef corals. Coral Reefs 14, 5–25 (1995).
Nurhati, I. S., Cobb, K. M. & Di Lorenzo, E. Decadal-scale SST and salinity variations in the central tropical pacific: Signatures of natural and anthropogenic climate change. J. Clim. 24, 3294–3308 (2011).
Juillet-Leclerc, A. & Schmidt, G. A calibration of the oxygen isotope paleothermometer of coral aragonite from porites. Geophys. Res. Lett. 28, 4135–4138 (2001).
Hartmann, A. C., Carilli, J. E., Norris, R. D., Charles, C. D. & Deheyn, D. D. Stable isotopic records of bleaching and endolithic algae blooms in the skeleton of the boulder forming coral Montastraea faveolata. Coral Reefs 29, 1079–1089 (2010).
Nozaki, Y., Rye, D. M., Turekian, K. K. & Dodge, R. E. A 200 year record of carbon-13 and carbon-14 variations in a Bermuda coral. Geophys. Res. Lett. 5, 825–828 (1978).
Al-Rousan, S. & Felis, T. Long-term variability in the stable carbon isotopic composition of Porites corals at the northern Gulf of Aqaba, Red Sea. Palaeogeogr. Palaeoclimatol. Palaeoecol 381–382, 1–14 (2013).
Klein, R., Pätzold, J., Wefer, G. & Loya, Y. Seasonal variations in the stable isotopic composition and the skeletal density pattern of the coral Porites lobata (Gulf of Eilat, Red Sea). Mar. Biol. 112, 259–263 (1992).
Paillard, D., Labeyrie, L. & Yiou, P. Macintosh Program performs time-series analysis. Eos, Trans. Am. Geophys. Union 77, 379 (1996).
Trenberth, K. E. & Dai, A. Effects of Mount Pinatubo volcanic eruption on the hydrological cycle as an analog of geoengineering. Geophys. Res. Lett. 34, 1–5 (2007).
Glynn, P. W. Monsoonal upwelling and episodic Acanthaster predation as probable controls of coral reef distribution and community structure in Oman, Indian Ocean. Atoll Research Bulletin 379, 379 (1993).
Claereboudt, M. R. Reef corals and coral reefs of the Gulf of Oman. Historical Association of Oman, 322 (2006).
Quinn, N. J. & Johnson, D. W. Cold water upwellings cover Gulf of Oman coral reefs. Coral Reefs-Journal of the International Society for Reef Studies 15.4, 214 (1996).
Asia-Pacific Data-Research Center. Available at: http://apdrc.soest.hawaii.edu (Accessed: March 2017).
Yamazaki, A. et al. Reconstructing palaeoenvironments of temperate regions based on high latitude corals at Tatsukushi Bay in Japan. J. Japanese Coral Reef Soc 11, 91–107 (2009).
Kroopnick, P. M. The distribution of 13C of ΣCO2 in the world oceans. Deep Sea Res. Part A. Oceanogr. Res. Pap. 32, 57–84 (1985).
Grottoli, A. G. & Wellington, G. M. Effect of light and zooplankton on skeletal δ13C values in the eastern Pacific corals Pavona clavus and Pavona gigantea. Coral Reefs 18, 29–41 (1999).
Peeters, F. The effect of upwelling on the distribution and stable isotope composition of Globigerina bulloides and Globigerinoides ruber (planktic foraminifera) in modern surface waters of the NW Arabian Sea. Glob. Planet. Change 34, 269–291 (2002).
Romanek, C. S., Grossman, E. L. & Morse, J. W. Carbon isotopic fractionation in synthetic aragonite and calcite: Effects of temperature and precipitation rate. Geochim. Cosmochim. Acta 56, 419–430 (1992).
Kumar, S. P. & Prasad, T. G. Formation and spreading of Arabian Sea high-salinity water mass. J. Geophys. Res. 104, 1455 (1999).
Reynolds, R. W. et al. Daily high-resolution-blended analyses for sea surface temperature. J. Clim. 20, 5473–5496 (2007).
Giese, B. S. & Carton, J. A. A Reanalysis of Ocean Climate Using Simple Ocean Data Assimilation (SODA). Mon. Weather Rev. 136, 2999–3017 (2008).
Liebmann, B. & Smith, C. A. Description of a complete (interpolated) outgoing longwave radiation dataset. Bull. Am. Meteorol. Soc. 77, 1275–1277 (1996).
Schrag, D. P. Rapid analysis of high-precision Sr/Ca ratios in corals and other marine carbonates. Paleoceanography 14, 97–102 (1999).
de Villiers, S. An intensity ratio calibration method for the accurate determination of Mg/Ca and Sr/Ca of marine carbonates by ICP-AES. Geochemistry Geophys. Geosystems 3 (2002).
Wessel, P. & Smith, W. H. F. Free Software Helps Map and Display Data. Eos Trans. AGU. 72(41), 441–446 (1991).
Acknowledgements
We acknowledge C. A. Grove, H. Takayanagi and K. Ohmori for their help with coral core-drilling and fieldwork at the Sultanate of Oman. K. Bremer supported ICP-OES analysis. CREES members at Hokkaido University provided assistance with slicing the coral core. This work was supported by JSPS KAKENHI Grant Number JP25257207.
Author information
Authors and Affiliations
Contributions
T.W. and M.P. designed the project. T.W., A.Y., M.P. M.R.C. collected samples. T.K.W., A.Y. and D.G.-S. analyzed the samples. T.K.W. wrote the manuscript. All authors helped with the interpretation of the data and writing the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Watanabe, T.K., Watanabe, T., Yamazaki, A. et al. Past summer upwelling events in the Gulf of Oman derived from a coral geochemical record. Sci Rep 7, 4568 (2017). https://doi.org/10.1038/s41598-017-04865-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-04865-5
This article is cited by
-
Loss of a globally unique kelp forest from Oman
Scientific Reports (2022)
-
Giant clam (Tridacna) distribution in the Gulf of Oman in relation to past and future climate
Scientific Reports (2022)
-
Oman coral δ18O seawater record suggests that Western Indian Ocean upwelling uncouples from the Indian Ocean Dipole during the global-warming hiatus
Scientific Reports (2019)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
