
Abstract
Males and females vary in many characteristics that typically underlie how well a host is able to fight infection, such as body-size, immune capacity, or energy availability. Although well studied in the context of sexual signalling, there is now growing recognition that these differences can influence aspects of pathogen evolution as well. Here we consider how co-infection between multiple pathogen strains is shaped by male-female differences. In natural populations, infections by more than one pathogen strain or species are believed to be a widespread occurrence. Using the water flea, Daphnia magna, we exposed genetically identical males and females to replicated bacterial co-infections. We found that pathogen transmission and virulence were much higher in females. However, males did not simply lower average pathogen fitness, but rather the influence of co-infection was more varied and less defined than in females. We discuss how pathogens may have more fitness benefits to gain, and consequently to lose, when infecting one sex over the other.
Similar content being viewed by others

Introduction
Males and females of the same species can be strikingly different from each other. They may differ in many overt traits, such as size, morphology or colouration1; but these differences can also extend to the immune response, with one sex often being more severely affected by infections2,3. In birds and mammals, for example, males are often more susceptible to infection and develop higher parasite loads than females4,5, whereas the reverse pattern is often observed in invertebrates6,7,8. When combined with the diversity of sex ratios, sex biases in dispersal, and general behavioural difference between males and females1,9,10, there is ample opportunity for dimorphism between the sexes to influence many aspects of disease epidemiology and evolution3,8,11.
Here we consider how differences between males and females can influence a widespread feature of host-pathogen interactions – the influence of co-infections on the outcome of infection. Multiple infections are believed to be more common in nature than infections composed of only a single parasite species or genotype12,13. Both theory and empirical studies have shown that the overall virulence of a co-infection can increase, decrease, or remain intermediate to the intrinsic properties of each infecting pathogen14. Rarely, however, have these concepts been explored within the context of sexual dimorphism in host resistance and exploitative potential. Co-infection studies typically either explore infection outcomes in only one sex15,16, or have not explicitly accounted for males-female differences17.
We explored the interplay between host sex and co-infection using the well-studied crustacean Daphnia magna and its obligate bacterial pathogen Pasteuria ramosa 15,18,19. In Daphnia up to 50 percent of all individuals in a population can be infected with two or more pathogen species20, or eight or more genotypes of the one pathogen species21. Male Daphnia, however, are significantly smaller, more resistant, and die younger than females22; potentially offering a parasite a more difficult pool of resources to exploit than females. To study how the contrasting environments provided by males and females impacted on host and pathogen fitness, we manipulated pathogen competition in genetically identical male and female Daphnia (genotype HU-HO-2), using three P. ramosa genotypes (C1, C14, and C19) with known infection characteristics. In line with theory14, the reduction in host lifespan (relative to uninfected controls) and the production of pathogen transmission spores were used as estimates of co-infection outcomes. We hypothesised that any differences between single and multiple infections seen in females would be dampened in males as a result of the sexual dimorphism in physiological characteristics.
Results
Our results reveal that the production of transmission spores under single and co-infections depended on the sex of the host (Fig. 1). While all pathogen genotypes performed significantly better in females (overall sex effect P < 0.01, Table 1), the influence of the pathogen combinations was sex-specific (overall infection term, P < 0.05, Table 1). This significant interaction was driven by the co-infection combinations where the two pathogens differ in their capacity to produce spores (Fig. 1C and D, see Table 1; but not Fig. 1B). Corresponding to the pattern of pathogen growth, we found that the relative changes in virulence were concordant for each sex (Fig. 2). Overall the average reduction in host lifespan was strongly and negatively correlated with the average production of mature transmission spores (correlation [95% CI]: females −0.93 [−0.99, −0.45]; males −0.98 [−0.99, −0.78]). Although females suffered a much higher reduction in lifespan than males (overall sex effect: P < 0.01, Table 1), there was no evidence for differences between the sexes in how well different pathogen combinations induced virulence (overall interaction term, P > 0.05, Table 1).

Transmission spores at death for all co-infection treatments. Pathogen combinations in panel A are ordered by spore loads in infected females (left to right, lowest to highest). Panels B to D show the same results subset by the co-infection combinations, with lower-case letters indicating significant groupings via post-hoc t-tests and Benjamini & Hochberg adjusted p-values.

Relative reduction in host lifespan for all co-infection treatments. Pathogen combinations in panel A are ordered by spore loads in infected females (left to right, lowest to highest). Panels B to D show the same results subset by the specific co-infection combinations, with lower-case letters indicating significant groupings via post-hoc t-tests and Benjamini & Hochberg adjusted p-values.
We compared virulence under coinfection with that expected based on interference competition (intermediate values), competition for resources (higher values than most virulent) or spite (lower values than the least virulent). Post-hoc comparisons, as indicted in Figs 1 and 2, revealed that co-infection scenarios similar to resource competition or spite are least likely, as co-infection never significantly exceeded the most virulent pathogen, nor was lower than the least virulent. Instead, co-infection largely matched the average of the innate characteristics of each infecting genotype in isolation; albeit with some sex-specific differences. When the two pathogens differed in their virulence or spore production, co-infection in females was indistinguishable from the average of the two single infections, and different to that predicted by either the least or most virulent strain (see Table 2). In contrast, coinfection in males was more varied, with one co-infection (C1 + C19) not significantly different to the predicted intermediate values, and another (C14 + C19) more similar to the most virulent and least transmissive strain (C19).
Discussion
Males and females vary in many characteristics that combine to offer pathogens two environments that differ in their exploitative potential3. Here, males of Daphnia magna are smaller and more resistant to infection than their genetically identical female counterparts22, a pattern common to many other arthropods6,7. Our results show that these differences change how well a pathogen can exploit a host, with pathogen transmission (mature spore loads) and virulence (reduction in expected lifespan) being much higher in females. However, the presence of the significant interaction term for pathogen transmission indicates that males do not simply lower average pathogen fitness, but rather the transition from the worst performing pathogen, to the mixed co-infection, to the best performing pathogen was different in each sex.
Emerging from our results is that the best performing pathogens have the most to gain from single infections in females, but also the most to lose from co-infections in the same sex. High-growth pathogen strains (C1 or C14) outperformed their lesser performing competitor (C19) in single infections by 2.1 million spores on average in females, versus only 0.55 million spores in males. As the evolutionary advantage of a pathogen depends its success relative to other competitors, not its absolute transmission, this represents a nearly four-fold improvement in relative fitness. Conversely, the same pathogens also experienced the greatest reduction in transmission success when co-infections occurred in females. Total spore loads for a co-infection were always less than that of the high-growth pathogen strain in isolation; a substantial loss of potential fitness even if the other pathogen was outcompeted completely. We also cannot rule out that the lesser performing pathogen in isolation (C19) is a superior competitor in a co-infection, a fact that would further decrease the fitness of the pathogen ranked best in isolation. Overall, this loss of fitness for high growth pathogens (and potential gain for low growth pathogens) was three-fold higher in females (1.2 million spores) than males (0.41 million spores).
The distinction between males and females was more subtle when considering the virulence of co-infections. For females, virulence in co-infections closely matched those expected by interference competition or immune-mediated competition between competing strains14. Overall virulence of a co-infection was firmly intermediate to that of the two competing pathogens, ruling out alternate explanations that predict either an increase (competition for a finite pool of resources, or facilitation via immune depletion) or a decrease (generation of spite) in virulence14. Evidence for interference competition-like mechanisms in males was less well-supported, as certain co-infection combinations tracked the overall characteristics of the least virulent pathogen. Given the lack of an overall significant interaction term for virulence, however, it appears that co-infection in males does not fundamentally change the driver of overall virulence, but rather that the patterns of co-infection in males may be less defined or extreme than in females.
As with any form of sexual dimorphism, underlying the observed differences in pathogen virulence and transmission may be a range of morphological, physiological, or immune based processes. In this system, males are more resistant (in terms of infection success) and so presumably also better equipped to limit the growth of a pathogen. For male and female Daphnia, however, an obvious point of difference is the divergence in body size22, whereby males are approximately 25 percent smaller than females. This could also explain the results we observe; irrespective of any immune response. With the larger body size and greater capacity for gigantism (i.e. pathogen induced growth) in females, there is naturally more resources available for a pathogen to exploit (energy or space for replication, for example). Infections in males may just be more constrained by the host, limiting both the potential fitness gains, and even loses, that a pathogen can derive from infection in this sex.
Our findings divert somewhat from previous studies in this system which have typically assumed that virulence tracks the most virulent co-infecting strain15, but increasingly it is becoming clear that co-infections depend as much on the specific genotype of the pathogens involved, as well as that of the host, and their interaction17,23. We suggest that heterogeneity between the sexes is an additional factor that is likely to influence co-infection outcomes by reducing the intensity of co-infection scenarios when males are infected. Extending this work to include genetic difference in the degree of sexual dimorphism, which we expect given the strong clonal differences observed in Daphnia, would further integrate the study of host-pathogen co-evolution with the evolution of male-female differences (see ref.3).
Methods
The host and pathogen genotypes used in this study were chosen as completely compatible and displayed strong genetic interactions for the severity of infectious disease in earlier studies19,24. The host genotype (HU-HO-2) originates from Hungary, while the three pathogen genotypes were C1 originating from Moscow, Russia; C19, derived from North Germany; and, C14 from Finland, Tvärminne. Various combinations of these clones, in some form, have been used previously in studies of co-infection23,25.
Production of male and female Daphnia
To reliably induce the production of males and females from a single Daphnia genotype, we used a modified protocol based on the work of Olmstead and Leblanc26. Briefly, we exposed the parental generation to a juvenoid hormone, methyl farnesoate (Echelon Biosciences, product number S-0153), after their first and second clutches, and then collected the third and fourth clutches for use in this experiment. Individuals were placed in 20-mL of the standard Daphnia media (ADaM27, modified after28), but supplemented with methyl farnesoate at a concentration of 300 μg/L. This media was changed either every two days, or when an animal produced a clutch. Importantly, we found that the hormone had no detectable effect on the subsequent fitness of the offspring.
A short pulse of hormone has been used in previous studies to study male-female differences in genetically identical Daphnia 29. To confirm that the hormone had negligible impact on the life-history of exposed animals, we conducted a series of experimental trials (see Table 3) using female Daphnia raised from mothers that were either exposed to the hormone or raised normally in standard Daphnia media. We found that there was no significant difference between the animals collected from treated and untreated mothers in the trait means and variances of both general life-history traits (lifespan and clutch production), and the characteristics of infection (infection rates; spore loads, lifespan, and clutch production of infected animals). Using this approach removes the need to stress mothers (low-light, high-density, or starvation) to induce male production; a process which can also influence infection characteristics19,30.
Experimental infection trials
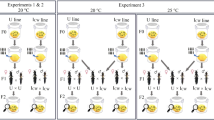
Animals were first raised individually in 60-mL jars containing 50 mL of artificial Daphnia medium following standard protocols18,19, and fed daily with algae (Chlorella vulgaris, up to 5 million cells animal−1 day−1). At four days old, individual males and females were exposed to 20,000 spores from a single P. ramosa genotype (C1, C14, and C19), or 20,000 spores containing an equal mix of two genotypes (C1 + C14, C1 + C19, or C14 + C19, 10,000 per genotype). As per standard approaches, this process was repeated the following day when the animals were five days old (40,000 spores in total) to ensure that an internal immune or physiological response, rather than moulting, contributed to differences in infection (see ref.31). In total, there were 12 treatment groups (two host sex groups × [3 single infections + 3 multiple infections]), with 36 animals initially assigned to each, plus 20 unexposed controls per sex (472 initial animals).
Survival was monitored daily and animals frozen on the day of death for the assessment of infection status and spore production using a Neubauer haemocytometer. As expected, overall infection rates were lower in males (mean ± SE: 0.58 ± 0.06; range: 0.35 to 0.70) than females (mean ± SE: 0.83 ± 0.04; range: 0.69 to 0.92). Analysis of variance revealed that infection rates indeed varied significantly with host sex (χ2 = 33.18, df = 1, P < 0.01) and the co-infection treatments (χ2 = 18.91, df = 5, P < 0.01), but were otherwise largely consistent (interaction term: χ2 = 6.13, df = 5, P = 0.29). Due to difference in the average lifespan between males and females (males: 33 days ± 1.9; females: 67 days ± 2.0), we subsequently focused on two traits of common currency: the production of transmission spores at host death; and, the reduction in lifespan as compared to the average of the unexposed controls.
Statistical analysis
All statistical analyses were performed in R (ver. 3.2.4; R Development Core Team, available at www.r-project.org). Initially there were 472 animals exposed to the co-infection treatments, arising from approximately 36 animals each assigned to 12 treatment groups (two host sex groups × [3 single infections + 3 multiple infections]) plus 20 male and 20 female unexposed controls. However, this sample size was reduced by the end of the experiment as: i) not all animals became infected; ii) they died before day 15 post-infection and so could not reliably have their infection status assessed; or iii) were removed from the study due to sampling errors (see Supplementary Information for more details). Before analyses, we checked for the assumptions of linear models (normality, homogeneity of variances). In the case of the spore loads at host death and the reduction in average lifespan, we used a white-adjusted analysis of variance to correct for unequal variances as implemented using the car package of R32.
Traits were first analysed using a full-factorial analysis of variance (Type III) with the infection treatment (various single or multiple infections), host sex (male or female), and their interaction as fixed effects. For each trait of interest, we began first with an overall test, using the entire dataset (both single and multiple infection together). If a significant effect was detected (host sex, infection treatment, or their interaction) we then explored each subset of pathogen combinations (e.g. C1 versus C14 versus C1 + C14) to see if specific single versus multiple infections combinations were contributing to the overall pattern. In this way, we avoid issues of multiple testing that would arise if we explored every related combination of single and multiple infections in isolation. Finally, one sample t-tests were used to compare the values for co-infection treatment groups against an expected value based on interference competition (intermediate values), competition for resources (higher values than most virulent) or spite (lower values than the least virulent).
Availability of materials and data
Datasets supporting the conclusions of this article are available via a figshare repository at https://dx.doi.org/10.4225/03/585732493b9ad.
Change history
28 September 2017
A correction to this article has been published and is linked from the HTML version of this paper. The error has been fixed in the paper.
References
Andersson, M. B. Sexual Selection. (Princeton University Press, 1994).
Zuk, M. The sicker sex. PLoS Path 5, e1000267 (2009).
Gipson, S. A. Y. & Hall, M. D. The evolution of sexual dimorphism and its potential impact on host-pathogen coevolution. Evolution 70, 959–968 (2016).
Poulin, R. Sexual inequalities in helminth infections: A cost of being a male? Am Nat 147, 287–295 (1996).
Schalk, G., Forbcs, M. R., Schalk, G. & Forbes, M. R. Male biases in parasitism of mammals: Effects of study type, host age, and parasite taxon. Oikos 78, 67–74 (1997).
Stoehr, A. M. & Kokko, H. Sexual dimorphism in immunocompetence: what does life-history theory predict? Behav Ecol 17, 751–756 (2006).
Sheridan, L. A. D., Poulin, R., Ward, D. F. & Zuk, M. Sex differences in parasitic infections among arthropod hosts: Is there a male bias? Oikos 88, 327–334 (2000).
Duneau, D. & Ebert, D. Host sexual dimorphism and parasite adaptation. Plos Biol 10, e1001271 (2012).
Cluttonbrock, T. H. & Iason, G. R. Sex-ratio variation in mammals. Q Rev Biol 61, 339–374 (1986).
Pusey, A. E. Sex-biased dispersal and inbreeding avoidance in birds and mammals. Trends Ecol Evol 2, 295–299 (1987).
Cousineau, S. V. & Alizon, S. Parasite evolution in response to sex-based host heterogeneity in resistance and tolerance. J Evol Biol 27, 2753–2766 (2014).
Read, A. F. The ecology of genetically diverse infections. Science 292, 1099–1102 (2001).
Petney, T. N. & Andrews, R. H. Multiparasite communities in animals and humans: frequency, structure and pathogenic significance. Int J Parasitol 28, 377–393 (1998).
Alizon, S., de Roode, J. C. & Michalakis, Y. Multiple infections and the evolution of virulence. Ecol Lett 16, 556–567 (2013).
Ben Ami, F., Mouton, L. & Ebert, D. The effects of multiple infections on the expression and evolution of virulence in a Daphnia-endoparasite system. Evolution 62, 1700–1711 (2008).
Taylor, L. H., Mackinnon, M. J. & Read, A. F. Virulence of mixed-clone and single-clone infections of the rodent malaria Plasmodium chabaudi. Evolution 52, 583–591 (1998).
Louhi, K.-R., Sundberg, L.-R., Jokela, J. & Karvonen, A. Interactions among bacterial strains and fluke genotypes shape virulence of co-infection. Proc R Soc B 282, 20152097–8 (2015).
Clerc, M., Ebert, D. & Hall, M. D. Expression of parasite genetic variation changes over the course of infection: implications of within-host dynamics for the evolution of virulence. Proc R Soc B 282, 20142820 (2015).
Hall, M. D. & Ebert, D. Disentangling the influence of parasite genotype, host genotype and maternal environment on different stages of bacterial infection in Daphnia magna. Proc R Soc B 279, 3176–3183 (2012).
Decaestecker, E., Declerck, S., De Meester, L. & Ebert, D. Ecological implications of parasites in natural Daphnia populations. Oecologia 144, 382–390 (2005).
Mouton, L., Nong, G., Preston, J. F. & Ebert, D. Variable-number tandem repeats as molecular markers for biotypes of Pasteuria ramosa in Daphnia spp. Appl Environ Microbiol 73, 3715–3718 (2007).
Duneau, D., Luijckx, P., Ruder, L. F. & Ebert, D. Sex-specific effects of a parasite evolving in a female-biased host population. BMC Biol 10, 104 (2012).
Ben Ami, F. & Routtu, J. The expression and evolution of virulence in multiple infections: the role of specificity, relative virulence and relative dose. BMC Evol Biol 13, 97–1 (2013).
Luijckx, P., Ben Ami, F., Mouton, L., Pasquier, Du, L. & Ebert, D. Cloning of the unculturable parasite Pasteuria ramosa and its Daphnia host reveals extreme genotype-genotype interactions. Ecol Lett 14, 125–131 (2010).
Izhar, R., Routtu, J. & Ben Ami, F. Host age modulates within-host parasite competition. Biol Lett 11, 20150131–20150131 (2015).
Olmstead, A. W. & Leblanc, G. A. Juvenoid hormone methyl farnesoate is a sex determinant in the crustacean Daphnia magna. J Exp Zool 293, 736–739 (2002).
Klüttgen, B., Dülmer, U., Engels, M. & Ratte, H. T. ADaM, an artificial freshwater for the culture of zooplankton. Water Research 28, 743–746 (1994).
Ebert, D., Zschokke-Rohringer, C. D. & Carius, H. J. Within-and between-population variation for resistance of Daphnia magna to the bacterial endoparasite Pasteuria ramosa. Proc R Soc B 265, 2127–2134 (1998).
Lohr, J. N., David, P. & Haag, C. R. Reduced lifespan and increased ageing driven by genetic drift in small populations. Evolution 68, 2494–2508 (2014).
Hall, M. D., Vettiger, A. & Ebert, D. Interactions between environmental stressors: the influence of salinity on host-parasite interactions between Daphnia magna and Pasteuria ramosa. Oecologia 171, 789–796 (2013).
Duneau, D. & Ebert, D. The role of moulting in parasite defence. Proc R Soc B 279, 3049–3054 (2012).
Fox, J. & Weisberg, S. An R Companion to Applied Regression. (SAGE Publications, 2011).
Acknowledgements
We thank V. Sundaramoorthi for assistance. This work was supported by an ARC Discovery Early Career Researcher Award to MDH.
Author information
Authors and Affiliations
Contributions
O.T., S.G., and M.H. conceived and designed the study. O.T. and S.G. carried out the lab work. O.T. and M.H. drafted the manuscript. All authors gave final approval for publication.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Change History: A correction to this article has been published and is linked from the HTML version of this paper. The error has been fixed in the paper.
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
An erratum to this article is available at https://doi.org/10.1038/s41598-017-12310-w.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Thompson, O., Gipson, S.A.Y. & Hall, M.D. The impact of host sex on the outcome of co-infection. Sci Rep 7, 910 (2017). https://doi.org/10.1038/s41598-017-00835-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-00835-z
This article is cited by
-
Features and associated factors of bacterial skin infections in hospitalized patients with pemphigus: a single-center retrospective study
Annals of Clinical Microbiology and Antimicrobials (2020)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
