
Abstract
Net primary productivity (NPP) plays a pivotal role in the global carbon balance but estimating the NPP of underwater habitats remains a challenging task. Seaweeds (marine macroalgae) form the largest and most productive underwater vegetated habitat on Earth. Yet, little is known about the distribution of their NPP at large spatial scales, despite more than 70 years of local-scale studies being scattered throughout the literature. We present a global dataset containing NPP records for 246 seaweed taxa at 429 individual sites distributed on all continents from the intertidal to 55 m depth. All records are standardized to annual aerial carbon production (g C m−2 yr−1) and are accompanied by detailed taxonomic and methodological information. The dataset presented here provides a basis for local, regional and global comparative studies of the NPP of underwater vegetation and is pivotal for achieving a better understanding of the role seaweeds play in the global coastal carbon cycle.
Measurement(s) | Seaweed net primary productivity |
Technology Type(s) | photorespirometry, biomass accumulation, eddy covariance |
Factor Type(s) | Annual per-area net primary productivity |
Sample Characteristic - Organism | Phaeophyceae • Rhodophyta • Chlorophyta |
Sample Characteristic - Environment | marine biome |
Sample Characteristic - Location | Global |
Similar content being viewed by others


Background & Summary
NPP is a major driver of ecological functioning and a key flux in the global carbon cycle1. The advent of remote sensing technologies has facilitated the measurement of terrestrial2,3,4, freshwater5,6, and oceanic7,8 NPP at unprecedented scales, with most global models of NPP available to date relying on space-based observations4,6. In contrast, the magnitude, patterns and determinants of spatial and temporal variation of primary productivity in the coastal ocean remains poorly understood9. This is particularly true for submerged vegetated habitats such as seaweed forests or seagrass beds, which are important contributors to coastal productivity globally10, but whose NPP cannot be measured accurately by satellite sensors as these perform poorly at shallow depths where submerged vegetation occurs (0–30 m)11. Rather, most observations rely on in situ measurements12. Existing measurements of coastal vegetation NPP vary however in methodology and are usually reported in different units, hindering our understanding of the role these habitats play in the carbon cycle and how it compares to other primary producers13. Additionally, the majority of measurements are conducted at local scales, which means compilation of multiple local-scale datasets is required to unravel larger spatiotemporal patterns12.
Seaweeds form the largest and most productive underwater vegetated habitat on Earth, drawing a flux of CO2 comparable to the Amazon rainforest every year14. The carbon assimilated through this production fuels local marine food webs15,16 and can constitute a trophic subsidy to areas with low primary production such as soft-bottom communities17. Recent studies also suggest that seaweed carbon makes important contributions to oceanic carbon export18, with some estimates identifying seaweeds as major contributors to oceanic carbon sequestration19. This has reopened the debate on their potential use as carbon dioxide removal and/or climate change mitigation tools20,21, although great uncertainties exist in the carbon fluxes they underpin19. Indeed, despite the fact that it has been more than 70 years since seaweeds were shown to be amongst Earth’s most productive organisms22,23,24, we still know little about how their NPP varies across taxa, space and time25. Previous attempts to collate seaweed NPP data at large spatial scales have been geographically restricted (e.g. refs. 13,26) or focused on specific taxa (e.g. refs. 27,28). These limitations have precluded a global understanding of the patterns and determinants of NPP across seaweed taxa, which is in urgent need to inform on the promising potential of seaweeds.
Here we describe the most comprehensive global dataset of marine macroalgae NPP gathered to date. Data was obtained from the primary literature or provided directly by authors and contains records from a total of 246 taxa from 429 sites in 72 different ecoregions. Measurements of seaweed NPP were collected at the taxa level and reflect per-area productivity rates across a range of depths and seaweed groups. Each record is accompanied by detailed descriptions of the methodology used and is classified into habitat groups depending on the growing substrate, vegetation height and dominant vegetation at the study site. The dataset can be used to answer a range of long-standing questions, from investigating productivity patterns across taxa, methods, locations, and habitats, to building the first global NPP products for shallow submerged vegetation29. Additionally, as nearly all records have geographic coordinates, NPP measurements can be linked to available environmental data layers (e.g. ref. 30).
Methods
Data compilation
An extensive search of published reports, PhD thesis, and the peer-reviewed literature was performed to capture studies dealing with the net primary productivity or biomass production of wild (i.e. not cultured) marine macroalgae. First, a formal search was performed in the Scopus database using the search terms “primary AND product* OR growth or npp AND (seaweed OR alga* OR kelp OR rocky AND reef OR turf OR temperate AND reef OR coral OR polar OR Arctic)”, which yielded 498 entries (April 2022). We then filtered the query by searching for relevant content in the title and abstract, yielding a total of 69 studies. Further searches were conducted in the China National Knowledge Infrastructure database (CNKI), J-STAGE repository (Japan), and Scientific Electronic Library Online (SciELO) to capture studies with English abstracts from underrepresented regions such as Asia and South America. Additional studies were included from existing reviews on the productivity of tropical31,32, temperate13,33 and polar seaweeds34 and from being cited in the scanned papers. Finally, we included a few more studies from MSc or PhD thesis, the authors’ unpublished data, and other published reports based on our knowledge of the research field.
Data selection and quality control
Given that our analysis was centered on patterns of annual areal carbon production by seaweeds, each of the potentially relevant studies was then evaluated against the following set of criteria to determine if they could be included in the final dataset. First, studies had to examine seaweed NPP or biomass accumulation on a per area basis. This criterion excluded studies examining biomass-specific productivity rates (e.g. refs. 26,35) unless those rates were applied to standing biomasses or covers in the field (e.g. ref. 36). Second, studies had to provide discrete estimates of NPP at the primary producer level (i.e. seaweed species or assemblage) with minimal interference of other photosynthetic or heterotrophic organisms. This criterion excluded studies examining net ecosystem primary production (NEP) and metabolism when the NPP of the seaweed component could not be accurately determined. Such studies usually relied on diel dissolved oxygen measurements in the water column (e.g. refs. 37,38), which often cannot resolve which organisms are responsible for primary production (but see ref. 39). An exception were oxygen measurements conducted directly above seaweed-dominated benthos (e.g. Aquatic Eddy Covariance method) with little heterotrophic respiration40,41. Third, studies had to capture seasonal variability in NPP across the year. This criterion excluded studies conducted at a single point in time, month or season, with the exception of studies concerning annual species where the growth or biomass accumulation was measured at the end of their life-cycle (i.e. the maximum period of growth). Fourth, quantification of productivity had to be performed in situ or outdoor mesocosms mimicking natural conditions. This criterion excluded laboratory-only experiments, aquaculture yields, model estimates (e.g. Ecopath models) and field studies in which the natural environmental conditions were experimentally modified (e.g. nutrient enrichment, acidification, sediment additions). Fifth, details of the specific sampling location and measuring method had to be provided. Sixth, studies had to provide new data not previously reported in other publications. This criterion excluded reviews, meta-analysis, as well as studies approximating NPP based on rates obtained elsewhere. After applying the criteria above, our final filtered dataset featured 1,084 records from 237 independent studies published between 1967 and 2022 and covered a range of seaweed vegetation types (Fig. 1a,b).

Temporal coverage of seaweed NPP measurements conducted at different habitat types and by tidal level (intertidal or subtidal), which are indicated in different colours. (a) Number of database records (i.e. a measurement of NPP per taxa, depth, site, year and method) depending on when the measurements were conducted. (b) Number of studies by date of publication (note not all data came from published studies).
Available data were extracted into an excel template from the suitable articles’ text, tables, figures (using the graph digitizing tool Webplot Digitizer42) or supplementary material. In our study, a record was considered to be the aerial net primary productivity of a taxon over the course of a year. If the data in a given study was not directly reported as annual rates, these where computed based on the monthly, bimonthly or seasonal means, with the corresponding standard deviation also being computed. The sampling effort (frequency of measurements throughout the year) was also recorded as it may have impacted the estimates’ accuracy. Data were entered into the template in the same units as the original source, but were also standardized to annual areal carbon production (i.e. g C m−2 y−1) to facilitate comparison. Values reported in fresh (16% or records) or dry (54% of records) weight (FW, DW, respectively) were converted to carbon units. Conversion factors provided in the studies were preferably used, but otherwise these were derived from the a single extensive database43 to minimize variability. Species or genus-specific factors were used in most cases, but family- and order-specific factors where occasionally used when these were not available for a given species. Metadata describing the depth; substrate; sampling year and season; taxonomy; study site and its geolocation; measuring method; and data extraction procedure were attached to each individual row. When a given value was not available, it was entered as “NA”. If a study reported NPP from multiple taxa, depths, sites, methods or time points, these were entered as separate case studies (separate rows). NPP of taxa within the same sample plot (e.g. multi-species Sargassum bed, kelp and understory algae) was also entered as separate records, but a specific column was created to denote that data would require summation of the rows to yield total areal productivity of that plot.
A site was defined as a single location where NPP was measured using the criteria above, with its geographic coordinates being added as metadata. If these were not directly provided in the article, we used the maps and/or description of the study locations to approximate their coordinates on Google Maps, noting also that these were approximations in the record’s metadata. NPP records across depths were considered to be within the same study site as long as measurements were within 30 m of each other. Each independent site was given a unique ID within each study.
As different sampling methods measure different aspects of photosynthesis and carbon assimilation13, we also recorded the method used to estimate each value of NPP. These were grouped into several subcategories, which mostly fell into two basic approaches: photorespirometry and biomass accumulation (Table 1). Photorespirometry-based methods measure direct carbon assimilation or oxygen evolution (which is later converted to carbon units based on the photosynthetic quotient), while biomass accumulation measures only the carbon destined to plant growth (mostly blade growth), and thus is expected to always yield lower estimates of NPP44. Biomass accumulation approximates well true NPP when carbon demand for growth matches carbon fixation45, but increasingly diverges when there is surplus of carbon derived from photosynthesis (e.g. in high light conditions) and carbon is directed towards other pathways (e.g. dissolved organic carbon exudation46). It is worth noting that both photorespirometry and biomass-accumulation-based methods are typically conducted at small spatial scales (1s m; plant or assemblage level), and therefore may not capture habitat-scale (10s m) NPP. Studies measuring oxygen and carbon fluxes directly over the water column or benthos (e.g. Aquatic Eddy Covariance) may provide better estimates of whole-ecosystem productivity, but these rarely resolve the taxon-specific contributions to productivity (but see ref. 37). An overview and discussion of the advantages disadvantages of each method is provided elsewhere (e.g. refs. 13,47,48).
Studies and taxa were also classified according to the habitat where measurements were performed using the information given within the published article (Table 2). Habitat categories were defined based on key structural parameters like vegetation height, the dominant vegetation (e.g. brown, red or green algae) as well as their position within the water column (benthic or pelagic). Within a study, taxa from different groups could be classed in the same habitat (e.g. canopy, epiphytes and understory algae all being part of a “marine forest”) unless they formed distinct patches within the habitat matrix (e.g. red algal bed patches interspersed with marine forests49,50), or the study examined different depth bands, sites or habitats. When incubations of different taxa were performed in isolation within a study, these were independently assigned a habitat category.
Data Records
The dataset, together with a reference list of all the studies included in it, is publicly accessible for download in the Figshare repository51.
Taxonomic coverage
The database contains NPP information for >240 species or taxonomic entities (e.g. crustose coralline algae, algal turf), from 49 families, 26 orders and all major seaweed groups and functional forms. The majority of species with NPP records are brown algae (55%; kingdom Phaeophyta) (Fig. 2a), with just over half the database being composed of records from the orders Laminariales and Fucales (558 records, Fig. 2b)

Taxonomic coverage of the database. Multiple denotes taxonomic groupings that involve species from different phyla (e.g. algal turfs).
Spatial and temporal coverage
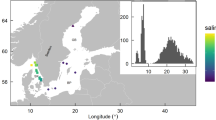
The dataset contains NPP data from 429 sampling sites (Fig. 3A) spanning from the high intertidal (3 m above mean sea level) down to 55 m (Fig. 3B). Sites span all major oceanographic realms and are distributed from the poles to the tropics, with most of the records concentrated in temperate latitudes 40–60° and concerning marine forests. The vast majority of studies measured NPP over 1-2 years. Only 2% of records report measurements conducted ≥3 years, and only three records report continuous NPP measurements >10 years. The temporal resolution of the measurements conducted within the sampling period varies from biweekly to annual measurements.

Location (A) and depth (B) of the study sites included in the database. Measurements conducted in the intertidal (i.e. above sea level are indicated in grey).
Data collection sources and methods
Records were mostly extracted from the published literature (94%), followed by PhD and MSc thesis (2.5%), unpublished personal data (2.5%) and a minor fraction from published reports in the grey literature. Most of the data were sourced from tables and text (73% of records), whilst the rest was extracted from graphs (20%) or from raw data (7%). The vast majority of NPP records in the database were obtained using biomass-accumulation-based methods (87%), followed by photorespirometry-based methods (12.9%), with only a minor fraction of records using both methods and Aquatic Eddy Covariance (n = 2). While biomass accumulation and photorespirometry measure different aspects of carbon assimilation, NPP patterns from both methods are largely consistent across latitude (Fig. 4). Biomass accumulation measurements are well distributed globally (Fig. 5a), while photorespirometry-based measurements are common in coral reefs (mostly on algal turfs), the open ocean (pelagic Sargassum spp. rafts), and a few other temperate locations (Fig. 5b). Despite its limitations48, biomass accumulation remains the most widely used methodology to estimate NPP, with photorespirometry-based studies showing a small decline over time (Fig. 6). This may be due to the rise of more novel methods resolving gas fluxes in marine ecosystems at higher temporal and spatial resolution (e.g. Aquatic Eddy Covariance). Adoption of these relatively novel methods in seaweed habitats is still not widespread however52, possibly due to the their relatively high cost and current inability to be deployed in wave-highly-exposed environments where many seaweeds thrive.

Latitudinal patterns of observed NPP depending on measuring methods. Dots indicate the average NPP of a study conducted within a given location.

Distribution of observations depending on the methodology applied to measure NPP. (a) Biomass-accumulation-based and (b) Photorespirometry-based methods.

Number of studies measuring seaweed NPP per decade grouped by broad methodology.
Technical Validation
The database was curated by the authors, with each of the records identifying who entered the data in the “Person_entering_data” column. We used templates to minimize spelling errors, inconsistencies, and incorrect values. Upon finalizing data entry, we conducted quality control by
-
i)
Checking taxonomic names. The validity of taxa names was checked using the taxon match tool of the World Register of Marine Species (WoRMS) in May 2021. The names were corrected and updated if taxonomies had changed since publication of the study.
-
ii)
Checking geographic coordinates. We projected the coordinates on a 1:10,000,000 shapefile of the world’s landmasses (EPSG:3857) checking they did not lay on land. When that was the case, we individually checked each value to ensure it was correct.
-
iii)
Checking for duplicates. Records with identical NPP values for the same species and GPS coordinates were double checked for accuracy.
-
iv)
Checking for outliers. Frequency histograms and quantile plots were generated to evaluate potential outliers. Records with very small (<1 g C m−2 y−1, i.e. 10% quartile) or large (<1,100 g C m−2 y−1, i.e. 95% quartile) NPP values were double checked for accuracy.
Usage Notes
Each of the records (rows) in our database provides the average annual aerial NPP and standard deviation (when reported) of a given taxon at a given site, depth, year and study and by a given measuring method. Given that records were collected across multiple individual studies conducted at different time points, for certain purposes, some records may not be directly comparable. Thus, each record is also accompanied by a series of metadata describing the taxonomic information, geographic coordinates, description of the measuring method used as well as vegetation and substrate type. The dataset variables’ (columns) definitions and descriptions can be found in Table 3. When the taxa measured includes species from multiple genera, families, orders or classes, this is indicated as “Mixed”.
Despite our efforts to obtain measurements across the globe, our dataset contains taxonomic, depth and geographical biases (Figs. 2, 3), with most records concerning brown algae from shallow depths (<10 m) and few records being available from South America, Africa, the Indian Ocean and Antarctica. We advise that researchers using the database should be aware of the influence these biases might have on their analyses.
Code availability
The code used to validate the dataset and make the figures in this manuscript is available at the Figshare repository51.
References
Field, C. B., Behrenfeld, M. J., Randerson, J. T. & Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science. 281, 237–240 (1998).
Knapp, A. K. & Smith, M. D. Variation among biomes in temporal dynamics of aboveground primary production. Science. 291, 481–484 (2001).
Gillman, L. N. et al. Latitude, productivity and species richness. Glob. Ecol. Biogeogr. 24, 107–117 (2015).
Anav, A. et al. Spatiotemporal patterns of terrestrial gross primary production: A review. Rev. Geophys. 53, 1–34 (2015).
Goldman, C. R., Jassby, A. & Powell, T. Interannual fluctuations in primary production: Meteorological forcing at two subalpine lakes. Limnol. Oceanogr. 34, 310–323 (1989).
Sayers, M. J., Fahnenstiel, G. L., Shuchman, R. A. & Bosse, K. R. A new method to estimate global freshwater phytoplankton carbon fixation using satellite remote sensing: initial results. Int. J. Remote Sens. 42, 3708–3730 (2021).
Behrenfeld, M. J. et al. Climate-driven trends in contemporary ocean productivity. Nature 444, 752–755 (2006).
Uitz, J., Claustre, H., Gentili, B. & Stramski, D. Phytoplankton class-specific primary production in the world’s oceans: Seasonal and interannual variability from satellite observations. Global Biogeochem. Cycles 24, GB3016 (2010).
Holt, J. et al. Modelling the global coastal ocean. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 367, 939–951 (2009).
Duarte, C. M., Losada, I. J., Hendriks, I. E., Mazarrasa, I. & Marbà, N. The role of coastal plant communities for climate change mitigation and adaptation. Nat. Clim. Chang. 3, 961–968 (2013).
Saba, V. S. et al. An evaluation of ocean color model estimates of marine primary productivity in coastal and pelagic regions across the globe. Biogeosciences 8, 489–503 (2011).
Duarte, C. M. et al. Seagrass community metabolism: Assessing the carbon sink capacity of seagrass meadows. Global Biogeochem. Cycles 24, 1–8 (2010).
Charpy-Roubaud, C. & Sournia, A. The comparative estimation of phytoplanktonic, microphytobenthic and macrophytobenthic primary production in the oceans. Mar. Microb. Food Webs 4, 31–57 (1990).
Duarte, C. M. et al. Global estimates of the extent and production of macroalgal forests. Global Ecology and Biogeography. 31(7), 1422–1439, https://doi.org/10.1111/geb.13515 (2022).
Duggins, D. O. & Estes, J. A. Magnification of secondary production by kelp detritus in coastal marine ecosystems. Science. 245, 170–173 (1989).
Dunton, K. H. & Schell, D. M. Dependence of consumers on macroalgal (Laminaria solidungula) carbon in an arctic kelp community: 13C evidence. Mar. Biol. 625, 615–625 (1987).
Krumhansl, K. A. & Scheibling, R. E. Production and fate of kelp detritus. Mar. Ecol. Prog. Ser. 467, 281–302 (2012).
Ortega, A. et al. Important contribution of macroalgae to oceanic carbon sequestration. Nat. Geosci. 12, 748–754 (2019).
Krause-Jensen, D. & Duarte, C. M. Substantial role of macroalgae in marine carbon sequestration. Nat. Geosci. 9, 737–742 (2016).
Bach, L. T. et al. Testing the climate intervention potential of ocean afforestation using the Great Atlantic Sargassum belt. Nat. Commun. 12, 2556 (2021).
Duarte, C. M., Wu, J., Xiao, X., Bruhn, A. & Krause-Jensen, D. Can Seaweed Farming Play a Role in Climate Change Mitigation and Adaptation? Front. Mar. Sci. 4 (2017).
Kanwisher, J. W. Photosynthesis and respiration in some seaweeds. in Some contemporary studies in marine science:: a collection of original scientific papers presented to Dr. S.M. Marshall, O.B.E., F.R.S. in recognition of her contribution with the late Dr. A.P. Orr to marine biological progress (eds. Barnes, H. & Marshall, S. M.) 407 (Allen & Unwin, 1966).
Blinks, L. R. Photosynthesis and productivity of littoral marine algae. J. Mar. Res. 14, 363–373 (1955).
Printz, H. Seasonal growth and production of dry matter in Ascophyllum nodosum. Avh. Utg. Av Det Nor. Videnskaps-akademi i Oslo. I. Mat. Klasse 4, 1–15 (1950).
Rassweiler, A., Reed, D. C., Harrer, S. L. & Nelson, J. C. Improved estimates of net primary production, growth and standing crop of Macrosystis pryifera in Southern California. Ecology 99, 2132 (2018).
Littler, M. M. & Arnold, K. E. Primary Productivity of Marine Macroalgal Functional-Form Groups From Southwestern North America. Journal of Phycology 18, 307–311 (1982).
Krause-Jensen, D. et al. Seasonal sea ice cover as principal driver of spatial and temporal variation in depth extension and annual production of kelp in Greenland. Glob. Chang. Biol. 18, 2981–2994 (2012).
Smale, D. A. et al. Environmental factors influencing primary productivity of the forest - forming kelp Laminaria hyperborea in the northeast Atlantic. Sci. Rep. 10, 12161 (2020).
Pessarrodona, A. et al. Global seaweed productivity. Science Advances https://doi.org/10.1126/sciadv.abn2465 (2022) (in press).
Assis, J. et al. Bio-ORACLE v2.0: Extending marine data layers for bioclimatic modelling. Glob. Ecol. Biogeogr. 27, 277–284 (2018).
Fulton, C. J. et al. Form and function of tropical macroalgal reefs in the Anthropocene. Funct. Ecol. 33, 989–999 (2019).
Tebbett, S. B. & Bellwood, D. R. Algal turf productivity on coral reefs: A meta-analysis. Mar. Environ. Res. 168, 105311 (2021).
Wernberg, T., Krumhansl, K., Filbee-Dexter, K. & Pedersen, M. F. Status and trends for the world’s kelp forests. in World Seas: An Environmental Evaluation: Ecological Issues and Environmental Impacts (ed. Sheppard, C.) 57–78, https://doi.org/10.1016/B978-0-12-805052-1.00003-6 (Academic Press, 2019).
Gómez, I. et al. Light and temperature demands of marine benthic microalgae and seaweeds in polar regions. Bot. Mar. 52, 593–608 (2009).
Kindig, A. C. & Littler, M. M. Growth and primary productivity of marine macrophytes exposed to domestic sewage effluents. Mar. Environ. Res. 3, 81–100 (1980).
Wanders, J. B. W. The role of benthic algae in the shallow reef of Curaçao (Netherlands Antilles) III: The significance of grazing. Aquat. Bot. 3, 357–390 (1977).
Hatcher, B. G. Reef primary productivity: a beggar’s banquet. Trends Ecol. Evol. 3, 106–111 (1988).
Odum, H. T. & Odum, E. P. Trophic Structure and Productivity of a Windward Coral Reef Community on Eniwetok Atoll. Ecol. Monogr. 25, 291–320 (1955).
Owen, D. P., Long, M. H., Fitt, W. K. & Hopkinson, B. M. Taxon-specific primary production rates on coral reefs in the Florida Keys. Limnol. Oceanogr. 1–14, https://doi.org/10.1002/lno.11627 (2020).
Attard, K. M. et al. Benthic oxygen exchange in a live coralline algal bed and an adjacent sandy habitat: An eddy covariance study. Mar. Ecol. Prog. Ser. 535, 99–115 (2015).
Attard, K. M. Seasonal metabolism and carbon export potential of a key coastal habitat: The perennial canopy-forming macroalga Fucus vesiculosus. Limnol. Oceanogr. 64, 149–164 (2019).
Rohatgi, A. WebPlotDigitizer. (2019).
Brey, T., Müller-Wiegmann, C., Zittier, Z. M. C. & Hagen, W. Body composition in aquatic organisms - A global data bank of relationships between mass, elemental composition and energy content. J. Sea Res. 64, 334–340 (2010).
Thom, R. M. Spatial and Temporal Patterns of Fucus distichus ssp. edentatus (de la Pyl.) Pow. (Phaeophyceae: Fucales) in Central Puget Sound. Bot. Mar. 26, 471–486 (1983).
Johnston, C. S., Jones, R. G. & Hunter, D. R. A seasonal carbon budget for a laminarian population in a Scottish sea-loch. Helgoländer wissenschaftliche Meeresuntersuchungen 30, 527–545 (1977).
Blain, C. O., Hansen, S. C. & Shears, N. T. Coastal darkening substantially limits the contribution of kelp to coastal carbon cycles. Glob. Chang. Biol. 1–17, https://doi.org/10.1111/gcb.15837 (2021).
Randall, J., Wotherspoon, S., Ross, J., Hermand, J. & Johnson, C. An in situ study of production from diel oxygen modelling, oxygen exchange, and electron transport rate in the kelp Ecklonia radiata. Mar. Ecol. Prog. Ser. 615, 51–65 (2019).
Rodgers, K. L., Rees, T. A. V. & Shears, N. T. A novel system for measuring in situ rates of photosynthesis and respiration of kelp. Mar. Ecol. Prog. Ser. 528, 101–115 (2015).
Sanderson, J. C. Subtidal Macroalgal Studies in East and South Eastern Tasmanian Coastal Waters. (University of Tasmania, 1990).
Miller, R. J., Reed, D. C. & Brzezinski, M. A. Community structure and productivity of subtidal turf and foliose algal assemblages. Mar. Ecol. Prog. Ser. 388, 1–11 (2009).
Pessarrodona, A. et al. A global dataset of seaweed net primary productivity, Figshare, https://doi.org/10.6084/m9.figshare.14882322 (2021).
Berg, P., Huettel, M., Glud, R. N., Reimers, C. E. & Attard, K. M. Aquatic Eddy Covariance: The Method and Its Contributions to Defining Oxygen and Carbon Fluxes in Marine Environments. Ann. Rev. Mar. Sci. 14, 431–455 (2022).
Lees, D. C., Houghton, J. P., Erickson, D. E., Driskell, W. B. & Boettcher, D. E. Ecological studies of intertidal and shallow subtidal habitats in lower Cook Inlet, Alaska. (1980).
Kelly, E. L. A. et al. A budget of algal production and consumption by herbivorous fish in an herbivore fisheries management area, Maui, Hawaii. Ecosphere 8, e01899 (2017).
Pedersen, M. F., Nejrup, L. B., Fredriksen, S., Christie, H. C. & Norderhaug, K. M. Effects of wave exposure on population structure, demography, biomass and productivity of the kelp Laminaria hyperborea. Mar. Ecol. Prog. Ser. 451, 45–60 (2012).
Kain, J. M. The biology of Laminaria hyperborea X. The effect of depth on some populations. J. Mar. Biol. Assoc. United Kingdom 57, 587–607 (1977).
Yatsuya, K., Nishigaki, T., Douke, A., Itani, M. & Wada, Y. Annual net productions of sargassacean species in coastal areas with different environmental characteristics in Kyoto Prefecture, the Sea of Japan. Nippon Suisan Gakkaishi 73, 880–890 (2007).
Carter, A. R. & Simons, R. H. Regrowth and Production Capacity of Gelidium pristoides (Gelidiales, Rhodophyta) under Various Harvesting Regimes at Port Alfred, South Africa. Bot. Mar. 30, 227–232 (1987).
Santelices, B., Vásquez, J., Ohme, U. & Fonck, E. Managing wild crops of Gracilaria in central Chile. in Eleventh International Seaweed Symposium (eds. Bird, C. J. & Ragan, M. A.) 77–89 (Springer Netherlands, 1984).
Pessarrodona, A., Foggo, A. & Smale, D. A. Can ecosystem functioning be maintained despite climate-driven shifts in species composition? Insights from novel marine forests. J. Ecol. 10, 91–104 (2018).
Dunton, K. H. An annual carbon budget for an arctic kelp community. in The Alaskan Beaufort Sea: ecosystems and environments. (eds. Barnes, P. W., Schell, D. & Reimnitz, E.) 311–326 (Academic press, 1984).
Klumpp, D. W. & McKinnon, A. D. Commmunity structure, biomass and productivity of epilithic algal communities on the Great Barrier Reef; dynamics at different spatial scales. Mar. Ecol. Prog. Ser. 86, 77–89 (1992).
Westphalen, G. & Cheshire, A. C. Quantum efficiency and photosynthetic production of a temperate turf algal community. Aust. J. Bot. 45, 343–349 (1997).
Morrissey, J. Primary productivity of coral reef benthic macroalgae. Proceedings of the 5th International Coral Reef Congress 77–82 (1985).
Howard, K. L. & Menzies, R. J. Distribution and Production of Sargassum in the Waters off the Carolina Coast. Bot. Mar. 12, 244–254 (1969).
Weigel, B. L. & Pfister, C. A. The dynamics and stoichiometry of dissolved organic carbon release by kelp. Ecology 102, 1–17 (2020).
Tait, L. W., South, P. M., Lilley, S. A., Thomsen, M. S. & Schiel, D. R. Assemblage and understory carbon production of native and invasive canopy-forming macroalgae. J. Exp. Mar. Bio. Ecol. 469, 10–17 (2015).
Rodgers, K. & Shears, N. Modelling kelp forest primary production using in situ photosynthesis, biomass and light measurements. Mar. Ecol. Prog. Ser. 553, 67–79 (2016).
Acknowledgements
All authors acknowledge funding from Euromarine. TW and KFD are grateful for support from the Australian Research Council (DP190100058 TW, KFD; DE190100692, KFD) and the Norwegian Blue Forest Network. PJM received support from NERC grant NE/S01169/1.
Author information
Authors and Affiliations
Contributions
A.P. conceived the idea and collected the data, curated the database, analysed the data, and drafted the manuscript. K.F.D., K.K., M.P.F., P.J.M. contributed to data compilation; All authors commented on the manuscript draft and approved its submission.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pessarrodona, A., Filbee-Dexter, K., Krumhansl, K.A. et al. A global dataset of seaweed net primary productivity. Sci Data 9, 484 (2022). https://doi.org/10.1038/s41597-022-01554-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41597-022-01554-5
This article is cited by
-
Deep-living and diverse Antarctic seaweeds as potentially important contributors to global carbon fixation
Communications Earth & Environment (2024)
-
Assessing photosynthetic uptake of total inorganic carbon in an Ecklonia cava dominated seaweed artificial reef: Population- and community-level metabolisms
Journal of Applied Phycology (2024)
-
Carbon capture by stipitate kelp forests in Peru: insights from population assessment of Lessonia trabeculata at 15°S
Journal of Applied Phycology (2024)
-
Benthic Incubation Chamber (BIC) for in-situ assessment of primary productivity in different canopy-forming communities
Marine Biology (2024)
-
A novel approach reveals underestimation of productivity in the globally important macroalga, Ascophyllum nodosum
Marine Biology (2022)
