
Abstract
The dataset presented here was collected by the GenTree project (EU-Horizon 2020), which aims to improve the use of forest genetic resources across Europe by better understanding how trees adapt to their local environment. This dataset of individual tree-core characteristics including ring-width series and whole-core wood density was collected for seven ecologically and economically important European tree species: silver birch (Betula pendula), European beech (Fagus sylvatica), Norway spruce (Picea abies), European black poplar (Populus nigra), maritime pine (Pinus pinaster), Scots pine (Pinus sylvestris), and sessile oak (Quercus petraea). Tree-ring width measurements were obtained from 3600 trees in 142 populations and whole-core wood density was measured for 3098 trees in 125 populations. This dataset covers most of the geographical and climatic range occupied by the selected species. The potential use of it will be highly valuable for assessing ecological and evolutionary responses to environmental conditions as well as for model development and parameterization, to predict adaptability under climate change scenarios.
Measurement(s) | growth ring • wood density |
Technology Type(s) | measuring table • calculation |
Factor Type(s) | tree species |
Sample Characteristic - Organism | Betula pendula • Fagus sylvatica • Picea abies • Populus nigra • Pinus pinaster • Pinus sylvestris • Quercus petraea |
Sample Characteristic - Location | Europe |
Machine-accessible metadata file describing the reported data: https://doi.org/10.6084/m9.figshare.11294993
Similar content being viewed by others


Background & Summary
Tree rings are an important archive of individual life history variation with a large range of applications across both natural and social sciences. In their annual growth rings, trees chronologically record the effect of any factor – ranging from local to global scale – that directly or indirectly affects radial growth processes1. Typical factors influencing annual tree growth are for example growing season temperatures or soil water availability in cold and arid regions, respectively. Large datasets from tree-ring series sampled across wide geographical ranges have been used in a broad range of studies, e.g. to infer climate variability2,3,4, reconstruct variation in streamflow5, investigate processes affecting forest dynamics6,7, identify the origin of wood used in ancient buildings8, and date historical tools and instruments9,10.
The most important archive of tree-ring data worldwide is the International Tree-Ring Data Bank (ITRDB11). It has more than 4250 centrally-held datasets of 226 tree species from all continents, except Antarctica12. However, most of these tree-ring datasets were obtained following classical dendrochronological protocols, which usually aim to maximize the climatic signals recorded in the ring-width series by sampling climatically stressed and old populations1. Such a sampling design is convenient for climate reconstructions but can lead to bias in terms of climate sensitivity when using these datasets to elucidate ecological and evolutionary processes13,14. This is particularly relevant given that classic site selection criteria predominantly targeted extreme micro-site conditions (e.g., ridge or treeline locations), and selectively excluded measurements with weak common growth signal.
The increasing use of dendrochronological techniques in transdisciplinary studies15 is driving demand for improved ecological representativeness in the global tree-ring archives, in particular by adding new datasets from non-stressed populations, which help better representing the full environmental niche of the species14. One example is the recent combination of evolutionary biology and dendrochronology to assess signs of local adaptation in trees by linking phenotypes inferred from tree rings to genomic and environmental information16,17,18. The advantage that dendrochronology provides in this context is that the outcome of a wide variety of growth-related traits acting over the lifespan of an individual can be inferred from a single sample (wood core). Such kind of integrated phenotypes can then be investigated together with other datasets, although the association analyses should also take in consideration that some external processes such as disturbances or forest dynamics can affect tree growth and limit the potential genetic and climatic information encoded in the tree-ring series. Following this approach, the European project GenTree (http://www.gentree-h2020.eu) aims to provide the first comprehensive pan-European assessment of phenotypic and genomic variation within and among environmentally contrasted populations across multiple tree species. To this end, the GenTree consortium has collected a dataset of tree-core characteristics from 142 sites located across the geographical range of seven ecologically and economically important European tree species. Measurements include the widths of all annual rings dated to the exact calendar year of formation, and whole-core wood density (measured for 125 sites), as well as complementary information at tree level such as tree height and diameter at stem breast height (DBH).
Here, we present this pan-European dataset of tree-ring width series and other fitness-related traits that cover wide geographical ranges and contrasting habitats of the studied species. Despite the limitation given by an underrepresented selection of individuals at each site (only 25), the potential of this dataset goes far beyond the GenTree project goals and will also be of value for assessing and/or modelling forest properties under climate change scenarios.
Methods
Site selection
Sampling covered seven of the most ecologically and economically important tree species in Europe: silver birch (Betula pendula Roth), European beech (Fagus sylvatica L.), Norway spruce (Picea abies (L.) Karst), European black poplar (Populus nigra L.), maritime pine (Pinus pinaster Aiton), Scots pine (Pinus sylvestris L.), and sessile oak (Quercus petraea (Matt.) Liebl.). Sites were selected using the following criteria: i) natural populations with no clear signs of natural or anthropogenic disturbances, ii) within or near to a EUFORGEN Gene Conservation Unit (http://portal.eufgis.org/search/), iii) no infrastructure at close proximity (houses, roads, electric cables, larges pipes), iv) no extreme slope, and v) reasonably accessible. Each sampled tree was georeferenced using hand-held global positioning systems. The centroid of all tree individuals was used to estimate the geographical position of each site. Elevation was extracted for each site using the global multi-resolution terrain elevation data 2010 (GMTED2010)19.
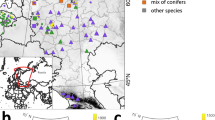
Sites were distributed across most of the geographical range of the species in Europe (Fig. 1). Climate-space diagrams were used to assess the relative climatic positions of each study site based on mean annual temperature and annual precipitation (Fig. 2). The geographical coordinates of tree occurrence in Europe were obtained from a reference publication20, and the corresponding climate data for the period 1979–2013 was extracted from CHELSA21 to plot full climate-space for each species, and overlaid the study sites on this distribution. The resulting plots show that the study sites are located in heterogeneous environmental conditions across a broad span of the climatic spaces occupied by the study species, covering contrasting habitats (Fig. 2).

Natural distributions of the seven selected tree species and the geographical location of each study site from which tree-ring width measurements were obtained (142 sites in total). Distribution maps were obtained from EUFORGEN (www.euforgen.org).

Climate-space diagrams for each species based on annual mean temperature and annual precipitation. Grey points represent species occurrences from across their total climate-space and red points show the climatic position of the selected study sites from which tree-ring measurements were obtained.
Sampling and laboratory protocols
Sampling took place from 2016 to 2018. At each study site, 25 dominant or co-dominant trees were selected. All trees were >25 m apart from each other to minimize the risk of sampling closely related individuals. Trees with visual symptoms of decay, infections, scars or abnormally low vigor were avoided. Each sampled tree was permanently labelled and one to three increment cores (depending on owner permission) were extracted at breast height (1.3 m) and perpendicular to the slope direction to avoid sampling reaction wood. Two cores were taken from one side of the stem and a third core from the opposite side. DBH was measured with a tape and height was estimated from ground to top of the tree using a clinometer.
The best core per tree, i.e. the core that was closest to the pith, without breakage or other obvious defects, was selected to conduct ring width measurements. Cores were air dried, mounted on wooden support beams, and then sanded with progressively finer sanding paper until wood cells were clearly visible under a binocular microscope. For silver birch and European black poplar, cores were surfaced along their cross-section using a core-microtome22 to obtain a clean cut plane surface facilitating the recognition of ring boundaries. Ring widths were measured to an accuracy of 0.01 mm using a binocular microscope connected to a LINTAB measuring device (Rinntech, Heidelberg, Germany). The exact year of formation was assigned to every annual ring through the cross-dating process1,23, by first visually cross-dating the tree-ring width series and then statistically verifying dating quality using the software CooRecorder (Cybis Elektronik & Data AB, Saltsjöbaden, Sweden). Missing rings, i.e. those that were absent within a series, were also actively detected and inserted into the series during the cross-dating process. Tree age at the coring height of 1.3 m was calculated as the length of the cross-dated tree-ring width series plus the estimated number of absent rings in the wood core towards the pith. The latter was estimated by fitting a template of concentric circles with known radii to the curve of the innermost rings and transforming the missing radius length into the number of absent rings. A summary of these parameters per population and species is reported in Online-only Table 1.
Whole-core wood density was determined on the second-best core (when available) and, in case the core was broken in several pieces, it was measured using the longest section. Wood volume was determined by the water-displacement method: the sample was immersed in a water-filled tray, which was placed on a balance. Weight of the displaced water was then converted to sample volume. Sample weight was measured on samples that had been dried in an oven at 102 °C for >2 hours (time required to obtain stable weights as reported by previous tests). Finally, wood density was obtained by dividing the sample weight by the sample volume.
The third core was kept as a reserve for any additional analyses. All original cores are stored at the Dendrosciences wood sample archives of the Swiss Federal Research Institute WSL in Birmensdorf (Switzerland) and can be made accessible upon request.
Data records
The dataset is composed of three comma-separated files, and one metadata file, which are freely accessible at Figshare repository24. The first file (site.csv) contains site descriptions including site identifier, geographical coordinates, elevation, and the contact details of the site coordinator. The second file (tree.csv) provides information at tree level, namely geographical coordinates, length of the ring width series, distance to the pith, estimated tree age, stem DBH, tree height, an assessment of dating confidence, and wood core density. The third file (trw_long_format.csv) contains annually-resolved tree ring width measurements of all trees included in the study (3600 trees in total). Missing values in the first two files (site.csv, and tree.csv) are denoted by NA. Missing ring measurements, defined as those actively detected during the cross-dating process, are denoted by 0 in the trw.csv file. The metadata file contains all definitions and unit for each variable. Additionally, rwl files of all sites containing the individual tree-ring width series (exact information than the trw_long_format.csv) are also included.
Both metadata and data files can also be accessed on the GnpIS information system at the following Gnpis Repository25. There, data will be updated as new data on additional GenTree species (Abies alba Mill., Pinus cembra L., Pinus halepensis Mill., Pinus nigra Arn., and Taxus baccata L.) are provided by partners.
Technical validation
Multiple steps were taken to ensure the technical quality of the measurements. The correct dating and quality of cross-dating was checked statistically with the software COFECHA26. It correlates each individual ring width series with the overall mean site series (after removing the series being tested). This analysis identifies mismatches and mistakes in the ring width measurements. The mean intercorrelation between raw individual series from the same site (Rbt.raw) was calculated and showed good within-site agreement (Online-only Table 1, Fig. S1). The expressed population signal (EPS), a measure of how well the mean series represents the common variability of the entire population if it were infinitely replicated, was also used to check the data quality. Low values of EPS usually indicate that the mean site series is influenced by individual processes rather than a consistent common signal. EPS values were calculated on the high-frequency domain (year-to-year variability) of the measured series. To do so, the low-frequency variability (decadal) was removed from the raw tree-ring series by applying a 32-year spline to each individual series. EPS was calculated in two different ways. To have an overview of the common variability shared by all the trees of a given site, EPS was calculated taking in consideration the common time period. In this case, most of the site chronologies (81%) presented an EPS above 0.85 (accepted threshold for signal strength in dendrochronological studies27) and only 19% showed an EPS lower than 0.85 (Online-only Table 1, Fig. S1). Due to the heterogenous age of the trees included in each site, we also calculated the EPS of the last 25 years but aiming at optimizing the maximum pairwise overlap. Similar percentages of sites presenting EPS above and below 0.85 were obtained (80% and 20%, respectively), but some of the sites that previously showed extremely low EPS values improved their EPS to reasonable values when assessing the maximum pairwise overlap. In general, low EPS values can be caused by a variety of factors such as short series length (not only old trees were selected), or low suitability for dendrochronological studies of some tree species such as silver birch and European black poplar. The sampling design did not specifically aim at selecting climatically limited populations, and consequently, the common signal of some sites might not be as strong as usually expected in dendrochronological studies, resulting in low EPS values. For this reason, and to complement the statistical assessment, the cross-dating confidence level of each dated ring series was classified (A = high confidence, B = possible doubts, C = very questionable). “A” letter was assigned to tree cores that were easily cross-dated and were well correlated with the rest of samples, “B” letter was assigned to cores with intermediate agreement with the rest of samples and/or showing small cracks in the wood, and “C” letter was assigned to tree cores that presented relatively low agreement with the rest of samples. The mean intercorrelation between detrended individual series from the same site was also calculated (Rbt.det, Online-only Table 1), which corroborated the generally good agreement among series in the high-frequency domains.
The average wood density per species was compared to those from a reference dataset28,29 (Fig. 3). As in this dataset, Norway spruce, black poplar, Scots pine, and maritime pine displayed lower mean density values than the ones obtained for silver birch, European beech and sessile oak (Online-only Table 1, Figs. 3 and S2).

Mean density and mean series length for each tree species and site. Vertical dashed black lines indicated the reference mean wood density values.
Change history
16 February 2021
A Correction to this paper has been published: https://doi.org/10.1038/s41597-020-0447-1
02 April 2020
A Correction to this paper has been published: https://doi.org/10.1038/s41597-020-0447-1
References
Fritts, H. Tree rings and climate. (Elsevier, 1976).
Büntgen, U. et al. 2500 years of European climate variablity and human susceptibility. Science 331, 578–582 (2011).
Alfaro-Sánchez, R. et al. Climatic and volcanic forcing of tropical belt northern boundary over the past 800 years. Nat. Geosci. 11 933–938 (2018).
Trouet, V., Babst, F. & Meko, M. Recent enhanced high-summer North Atlantic Jet variability emerges from three-century context. Nat. Commun. 9, 1–9 (2018).
Harley, G. L. et al. Suwannee River flow variability 1550–2005 CE reconstructed from a multispecies tree-ring network. J. Hydrol. 544, 438–451 (2017).
Cailleret, M. et al. A synthesis of radial growth patterns preceding tree mortality. Glob. Chang. Biol 23, 1675–1690 (2017).
Cailleret, M. et al. Early-warning signals of individual tree mortality based on annual radial growth. Front. Plant Sci. 9, 1–14 (2019).
Domínguez-Delmás, M., van Daalen, S., Alejano-Monge, R. & Wazny, T. Timber resources, transport and woodworking techniques in post-medieval Andalusia (Spain): Insights from dendroarchaeological research on historic roof structures. J. Archaeol. Sci. 95, 64–75 (2018).
Grissino-Mayer, H. D., Sheppard, P. R. & Cleaveland, M. K. A dendroarchaeological re-examination of the ‘Messiah’ violin and other instruments attributed to Antonio Stradivari. J. Archaeol. Sci. 31, 167–174 (2004).
Tegel, W., Muigg, B. & Büntgen, U. The wood of Merovingian weaponry. J. Archaeol. Sci. 65, 148–153 (2016).
Grissino-Mayer, H. D. & Fritts, H. C. The International Tree-Ring Data Bank: An enhanced global database serving the global scientific community. Holocene 7, 235–238 (1997).
Zhao, S. et al. The International Tree-Ring Data Bank (ITRDB) revisited: Data availability and global ecological representativity. J. Biogeogr. 46, 355–368 (2018).
Klesse, S. et al. Sampling bias overestimates climate change impacts on forest growth in the southwestern United States. Nat. Commun. 9, 5336 (2018).
Babst, F. et al. When tree rings go global: Challenges and opportunities for retro- and prospective insight. Quat. Sci. Rev. 197, 1–20 (2018).
Büntgen, U. Re-thinking the boundaries of dendrochronology. Dendrochronologia 53, 1–4 (2019).
Housset, J. M. et al. Tree rings provide a new class of phenotypes for genetic associations that foster insights into adaptation of conifers to climate change. New Phytol. 218, 630–645 (2018).
Heer, K. et al. Linking dendroecology and association genetics in natural populations: Stress responses archived in tree rings associate with SNP genotypes in silver fir (Abies alba Mill.). Mol. Ecol. 27, 1–11, https://doi.org/10.1111/mec.14538 (2018).
Trujillo-Moya, C. et al. Drought sensitivity of Norway spruce at the species’ warmest fringe: Quantitative and molecular analysis reveals high genetic variation among and within provenances. G3 8, 1225–1245 (2018).
Danielson, J. J. & Gesch, D. B. Global multi-resolution terrain elevation data 2010 (GMTED2010). US Department of the Interior and US Geological Survey, 2011–1073 (2011).
Mauri, A., Strona, G. & San-Miguel-Ayanz, J. EU-Forest, a high-resolution tree occurrence dataset for Europe. Sci. Data 4, 1–8 (2017).
Karger, D. N. et al. Climatologies at high resolution for the earth’s land surface areas. Sci. Data 4, 1–20 (2017).
Gärtner, H. & Nievergelt, D. The core-microtome: A new tool for surface preparation on cores and time series analysis of varying cell parameters. Dendrochronologia 28, 85–92 (2010).
Stokes, M. A. & Smiley, T. L. An Introduction to Tree-Ring Dating. (The University of Arizona Press, 1966).
Martínez-Sancho E. et al. The GenTree Dendroecological Collection, tree-ring and wood density data from seven tree species across Europe. figshare. https://doi.org/10.6084/m9.figshare.c.4561037 (2019).
Steinbach, D. et al. GnpIS: an information system to integrate genetic and genomic data from plants and fungi. Database 2013, bat058, https://doi.org/10.1093/database/bat058 (2013).
Holmes, R. L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 43, 68–78 (1983).
Wigley, T. M., Briffa, K. R. & Jones, P. D. On the average value of correlated time series, with applications in dendroclimatology and hydrometereology. J. Clim. Appl. Metereology 23, 201–213 (1984).
Chave, J. et al. Towards a worldwide wood economics spectrum. Ecol. Lett 12, 351–366 (2009).
Zanne, A. E. et al. Global wood density database. Dryad., https://doi.org/10.5061/dryad.234 (2009).
Acknowledgements
This publication is part of a project that has received funding from the European Union’s Horizon 2020 research and innovation programme under grant agreement No. 676876 (GenTree). This work was also supported by the Swiss Secretariat for Education, Research and Innovation (SERI) under contract No. 6.0032. We want to thank to Olivier Ambrosio, Francisco Auñón, Virgilijus Baliuckas, Eduardo Ballesteros, Giorgia Beffa, Sabine Brodbeck, William Brunetto, Jurata Buchovska, Fernando del Caño, Andreas Fera, Francis Gauvrit, Jean-Marc Gion, Rene Graf, Berit Gregorsson, Markus Hartmann, Andreas Helmersson, Enja Hollenbach, Jan Philipp Kappner, Johannes Lambertz, David Lopez-Quiroga, Jérémy Marchon, David Matter, Benjamin Meier, Helge Meischner, Pekka Närhi, Daniel Nievergelt, Juri Nievergelt, Anne Eskild Nilsen, Hans Nyeggen, Geir Østreng, Gilbert Pichet, Sebastian Richter, Christoph Rieckmann, Marcus Stefsky, Sergio San Segundo, Ivan Scotti, Jørn Henrik Sønstebø, Arne Steffenrem, Jussi Tiainen, Anne Verstege and Mikael Westerlund for their support during the field campaigns. We are also grateful to all the forest owners and national administrations for providing sampling permissions.
Author information
Authors and Affiliations
Contributions
F.V., L.O., S.C. and B.F. prepared the sampling design. All the authors contributed to the field sampling. L.S. coordinated the core inventory with support of S.M. and performed the tree-ring width measurements, including the cross-dating. E.M.-S. checked data quality with help from F.K., wrote the manuscript and prepared the relative data files. B.C., C.G. and P.F. performed the wood density measurements. B.D. checked the quality of site coordinates. C.R., F.G., B.D., L.O., K.H., G.v.A., E.M.-S. and P.F. coordinated all the activities. All authors commented on earlier versions and approved the current version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Online-only Table
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
The Creative Commons Public Domain Dedication waiver http://creativecommons.org/publicdomain/zero/1.0/ applies to the metadata files associated with this article.
About this article

Cite this article
Martínez-Sancho, E., Slámová, L., Morganti, S. et al. The GenTree Dendroecological Collection, tree-ring and wood density data from seven tree species across Europe. Sci Data 7, 1 (2020). https://doi.org/10.1038/s41597-019-0340-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41597-019-0340-y
This article is cited by
-
Gut microbiome-metabolome interactions predict host condition
Microbiome (2024)
-
Comprehensive molecular interaction map of TGFβ induced epithelial to mesenchymal transition in breast cancer
npj Systems Biology and Applications (2024)
-
Effects of experimental canopy openness on wood-inhabiting fungal fruiting diversity across succession
Scientific Reports (2024)
-
Spatial partitioning of terrestrial precipitation reveals varying dataset agreement across different environments
Communications Earth & Environment (2024)
-
Plastiphily is linked to generic virulence traits of important human pathogenic fungi
Communications Earth & Environment (2024)
