
Abstract
The 2016 version of the FosSahul database compiled non-human vertebrate megafauna fossil ages from Sahul published up to 2013 in a standardized format. Its purpose was to create a publicly available, centralized, and comprehensive database for palaeoecological investigations of the continent. Such databases require regular updates and improvements to reflect recent scientific findings. Here we present an updated FosSahul (2.0) containing 11,871 dated non-human vertebrate fossil records from the Late Quaternary published up to 2018. Furthermore, we have extended the information captured in the database to include methodological details and have developed an algorithm to automate the quality-rating process. The algorithm makes the quality-rating more transparent and easier to reproduce, facilitating future database extensions and dissemination. FosSahul has already enabled several palaeoecological analyses, and its updated version will continue to provide a centralized organisation of Sahul’s fossil records. As an example of an application of the database, we present the temporal pattern in megafauna genus richness inferred from available data in relation to palaeoclimate indices over the past 180,000 years.
Similar content being viewed by others
Introduction
The study of fossils provides essential insights into past ecological processes1, and the origins and evolution of species through time2,3. With the advancement of dating methods, the number of reliably dated fossil records — i.e., those with quantified confidence intervals encompassing the true age of the specimen4 — is steadily increasing. However, to apply the information of these records in comprehensive analyses, they first need to be compiled and organized in a standardized and detailed framework. The development of the FosSahul database5,6 unified and quality-controlled fossil specimens across the Sahul region (i.e., mainland Australia, Tasmania, New Guinea, joined at times of lower sea levels), and has opened many opportunities for new, broad-scale analyses of ecological processes7. For example, since its compilation, the database has provided a robust source of data to investigate extinction drivers at both continental and regional scales in Australia8,9,10, and to identify new areas for potential fossil discovery11.
To maintain FosSahul’s utility, given the growing wealth of new fossil data being published each year, regular updates and maintenance are essential to ensure this database incorporates the latest fossil discoveries and remains relevant. Furthermore, FosSahul includes its own quality-rating scheme based on a two-stage set of objective criteria4. Because dating methods will continue to improve and new research will provide more reliable fossil ages of both existing and newly discovered fossils, it is important to have a fully transparent and easily reproducible quality-rating method.
In this paper, we provide the latest updated version of the FosSahul database (FosSahul 2.0) and present an automated algorithm that runs the two-stage quality assessment4 on dated fossil records in the database. We describe this new feature and demonstrate the versatility and applicability of FosSahul by using the resulting high-quality ages to calculate a megafauna (>44 kg adult mass12) vertebrate richness indicator13 for the last 180,000 years across Australia, and visually link this to palaeoclimate metrics (i.e., mean annual temperature and precipitation anomalies and velocity, see Methods) based on climate hindcasts from the LOVECLIM Earth-system model14.
Results
Database update
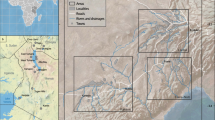
Compared to its originally published version, the updated FosSahul (2.0) database15 contains ~ 28% more dated fossil records (11,871 in total, see Table 1) compiled from 160 published sources (previously 144). Of these records, 27% are fossils of megafauna. The fossil records stem from 605 deposits (an increase of 67%) and encompass 1% more species and genera than FosSahul 1.0. Of all records, 14% (1,626) received a quality rating of A* or A. These 1,626 records were located on the Australian mainland, Tasmania and Kangaroo Island, with a concentration in south-eastern Australia, in the coastal regions of Western Australia and along the Great Australian Bight (Fig. 1). There are no reliable records (B and below) for central and northern Australia and the rest of Sahul, and all reliable records older than 120,000 years are concentrated in three locations (Leeuwin–Naturaliste Region in Western Australia, Naracoorte in South Australia, and Cuddie Springs in New South Wales) (Fig. 1). Ages rated B or C can still be useful, for example, in analyses only requiring information on the presence or absence of species11, and so we retained these in the database.

Distribution of dated non-human vertebrate fossil records in the Sahul region for different time periods of the Late Quaternary (up to 897,000 years ago) rated as reliable (A* or A; blue circles) and unreliable (B or C; red triangles).
Vertebrate richness and number of records in relation to climate
The corrected sampled-in-bin diversity index13 applied to high-quality megafauna ages of FosSahul 2.0 shows an increase (from 1.5 to 10; Fig. 3, top panel) in diversity of megafauna specimens until ~ 90,000 years ago, followed by a decrease until ~ 30,000 years ago when mean annual temperature slightly increased (+1 °C) after a drop of ~4 °C between 80,000 and 60,000 years ago. This variation in temperature seems to be part of a climate cycle that occurred also between 200,000 and 80,000 years ago. Such a climatic cycle is present in the mean annual precipitation outputs showing a succession of increase/decrease in precipitation approximatively every 20,000 years. Both temperature and precipitation velocity had low variation (i.e., <40 and 8 m year−1, respectively) across Sahul for the last 200,000 years. Although only ranging from 100,000 to 30,000 years ago, the diversity index of the southeast Australian subsample follows the same trend as that from the complete dataset, indicating that the richness estimates are probably still indicative of real trends despite the spatial bias potentially caused by the uneven distribution of fossil records in the full dataset.
Discussion
The updated and extended FosSahul database (FosSahul 2.0) now contains a total of 11,871 dated non-human vertebrate fossil records from the Quaternary, originating from 160 sources published up to 2018. Compared to FosSahul 1.0, the updated version contains fewer records with a high-quality rating. The main reason for this is that the previous version of the database was rated ‘manually’ without the standardized consistency of the now-implemented algorithm. Therefore, although the authors relied on expert consultation for the quality ratings provided in FosSahul 1.0, not all of the information used for each record was consistently recorded in the first version of the database. This was a weakness of FosSahul 1.0, and the main reason for our extension of the database to include additional, detailed information, and the development of the R code to do the rating objectively. The new database is now accompanied by an algorithm (Fig. 2) that automates the previously developed quality-rating procedure for fossil ages4.

Flowchart presenting the quality-rating method implemented by the automated algorithm. Panel (a) shows the first step of the rating procedure using the example of an uranium-thorium series (U-Th) age for teeth, panel (b) shows the second step. Based on the pre-quality rating received in the first step (a), ages are given a final rating depending on whether they are direct or indirect ages and (only for indirect ages) their association with the dated material. Panel (b) has been adapted from Rodríguez-Rey et al.4.

Genus-level corrected, sampled-in-bin diversity index calculated from FosSahul 2.0 high-quality ages (i.e., scored A* and A) for megafauna specimens (from Sahul = green; from south-eastern Australia = orange), number of megafauna records (based on the full dataset = light grey; high-quality ages only = dark grey; high-quality ages for south-eastern Australia only = thin black bars), mean annual temperature (°C) and precipitation anomaly (mm day−1) relative to the present day, temperature velocity (m year−1), and precipitation velocity (m year−1) across time (in thousands of years before present). Both the ‘corrected, sampled-in-bin diversity index’ and the ‘number of records’ are calculated using 10,000-year time increments, with the oldest records dated to 180,000 years before present. Climate variable plots show the median value (solid line), and the 25th and 75th percentiles (light shading) calculated across Sahul. Yellow shading represents putative arrival window (including uncertainties) of humans in Sahul, see Bradshaw et al.41 for discussion.
While the algorithm is tailored to FosSahul, it could easily be adapted to automate similar ratings on other databases. Although this algorithm has been designed to be user-friendly and to make the quality-rating process easier and less subjective, the user still has to make decisions in the process that require a sound understanding of the specific dating techniques and the quality-rating procedure itself. We therefore recommend that experts should be involved to avoid drawing misinformed conclusions regarding the quality of any age estimate. For future versions of FosSahul, the algorithm will be highly useful to non-specialists to rate the reliability of ages for new records that will be added to the database as new sources are published. Furthermore, it will allow a rapid recalculation of ratings as geochronologists reassess which methods and quality-assurance indicators are appropriate. This will facilitate the process of maintaining FosSahul as a database that reflects current scientific findings. The fact that fossil records are not evenly distributed across the continent underlines the need for new fossil discoveries, as well as improved dating methods and re-dating of existing fossils.
Climate change and/or the arrival of humans have been linked to an increase in megafauna extinctions in Sahul8,10,16,17,18,19,20,21, but there are many more elements that determine vertebrate richness across such broad spatial scales22,23,24. Therefore, it is essential to consider multiple potential drivers simultaneously, as well as the spatial patterns of biodiversity turnover when making conclusions about palaeoecological trends through time10.
In terms of climate, previous studies have highlighted that palaeoclimate trends in Australia are dominated by temporal variation in precipitation and/or aridity rather than temperature25, with important implications for extinctions — indeed, a decline in water levels at Lake Eyre and Lake Frome between 50,000 and 40,000 years before present were possibly related to local megafauna extinctions26, and Dortch et al.27 invoked drought as a cause of extinctions for Macropus spp. in south-eastern Australia. Spatial or temporal variation in precipitation has also been used to predict patterns in plant e.g.28,29 and mammal e.g.30,31 species richness, also across a wide range of latitudes32. Furthermore, water variables are strongly associated with vertebrate richness in regions with predominantly warm climates32. Climate stability has previously been associated with high biodiversity, given that environmental stability tends to minimize extinction and facilitate speciation through specialization and divergent selection33,34. However, given that the data in FosSahul only cover the last ~ 180,000 years at a continental scale, it is understandable that gross megafauna diversity patterns might not be explained by variation in climate stability over time8,35. Furthermore, here we only show patterns in vertebrate records at the broadest of scales, and climate conditions at finer scales potentially explain additional drivers of extinction patterns10 (Fig. 1). Although the current version of FosSahul does not contain records for investigating a finer-scale variation in diversity (at least not before 80,000 years ago), this is nevertheless an example of the type of analyses for which databases such as FosSahul can be used, particularly as more high-quality ages become available.
Overall, our study showcases the utility of the FosSahul database and highlights the importance of comprehensive databases for palaeoecological investigation. We emphasize that FosSahul is open to improvement and updates, and we encourage users to provide feedback on the database and the quality-rating algorithm.
Methods
Fossil data records
The original FosSahul database5 compiled Pleistocene to Holocene ages for non-human vertebrate fossil records from the Sahul region published in the scientific literature until 2013. When the database was assembled, Rodríguez-Rey et al.4 established a set of criteria to provide objective quality-rating criteria for fossil ages. We searched the literature for new fossil ages published from 2013 onwards and have updated the existing FosSahul website with these new records. Following the methods described in Rodríguez-Rey et al.5, we focused our literature search on megafauna records only because the collation of these was the original purpose of FosSahul. We only included non-megafauna vertebrate fossils when published alongside megafauna fossil dates. While these non-megafauna records are still useful, this part of the database was compiled opportunistically and is therefore not a complete representation of the available data. It is possible that a future version of FosSahul will include all fauna data; however, we have no such imminent plans.
Automated quality-rating algorithm
We developed an algorithm to automate the two-step quality rating (Fig. 2b) described in Rodríguez-Rey et al.4 and Rodríguez-Rey et al.5. While Rodríguez-Rey et al.5 established the quality-rating system and compiled the initial FosSahul database, quality-rating had to be done manually5. To reduce errors and to make the dating process more objective, reproducible and transparent, we extended the database to include the necessary fields providing more detail on dating techniques (Online-only Table 3) to run the rating automatically based on the quality criteria described in Rodríguez-Rey et al.4 (Table 2, Fig. 2).
In the first step, the algorithm rates the quality of the dating procedure based on the dated material (e.g., bone, charcoal, quartz grains) and the technique (e.g., 14C radiocarbon dating, amino acid racemization, luminescence dating, electron spin resonance, uranium-thorium series), including pre-treatments (if applicable). During this step, ages are assigned a pre-quality category (m*, m, B or C). For ages rated B or C, this rating is final. For ages that receive a pre-quality rating of m* or m, the second step of the algorithm rates these ages based on the association of the dated material with the remains of the target species4,5. For direct ages where the dated material is from the target species, the association is clear and the final rating is A* or A. For indirect ages, the association is assessed based on their specific depositional context (certain association = A; uncertain association = B; no or unknown association = C), with A being the highest possible rating for indirect ages (Table 2).
Climate variables
We used a climate reconstruction based on the three-dimensional Earth-system model LOVECLIM14,36. The model includes representations of the atmosphere, ocean, sea ice, ice sheets, the carbon cycle, land surface, as well as a vegetation model that simulates the dynamics of two terrestrial plant functional types — trees and grasses. We simulated 1000-year average climates over the past 180,000 years using LOVECLIM and downscaled the output resolution to 1×1° using bilinear interpolation. For each temporal snapshot and grid cell, we extracted annual mean precipitation, and mean annual temperature. We then calculated the anomaly of each variable relative to present day.
Index of vertebrate diversity in the database and climate-change metrics
Using only the high-quality fossil ages (A* and A), we calibrated the radiocarbon ages using the SHCal13 curve37 in OxCal 4.338 and calculated a corrected sampled-in-bin diversity index (to genus level at the Australian continental scale) using the R package divDyn13 as a temporal measure of taxonomic richness represented in the database. This metric uses the richness per bin, but includes a measure of sampling completeness based on consecutive time bins as a correction for missing species that should have been present39. However, given the constraints of fossil preservation and patchy distribution of palaeontological excavation sites, the calculated richness index is likely still biased by uneven sampling effort. We used 10,000-year time increments with the oldest records dated to 180,000 years before present. Although the database contains fossil records for other areas of the Sahul region, none of those received a high-quality rating. Our analysis therefore only focuses on the Australian continent. To limit the spatial bias caused by the uneven distribution of fossil records here, we also analysed a subset of the data from south-eastern Australia (longitude < 130 E, latitude > 24 S) that has a more even distribution of records.
Climate anomaly (of a given climate variable relative to the present day) is one of the most common metrics used to describe exposure to climate change, but climate velocity is likely more biologically relevant than climate anomalies because velocity accounts for regional changes in climate and the ability of topographic heterogeneity to buffer biota against these changes40. Climate velocity describes the rate and direction that an organism would need to migrate to maintain an isocline of temperature and precipitation variables40. This proxy is often considered synonymous with the rate of climate displacement for a species, such that the higher the velocity, the less probable a species will keep pace with its shifting climatic niche and thus survive. We calculated vectors of climate velocity (m year−1) for mean annual temperature (°C) and mean annual precipitation (mm day−1) following established methods40 from the palaeoclimate reconstruction provided by LOVECLIM over the last 180,000 years. We divided the rate of climate change through time (°C year−1) by the spatial gradient in climate at that location (°C m−1).
Data availability
Data are available on figshare15 (https://doi.org/10.6084/m9.figshare.8796944) and on the Global Ecology Flinders GitHub repository (https://github.com/GlobalEcologyFlinders/FosSahul).
Code availability
Code is available on figshare15 (https://doi.org/10.6084/m9.figshare.8796944) and on the Global Ecology Flinders GitHub repository (https://github.com/GlobalEcologyFlinders/FosSahul).
References
van Dam, J. A. Scanning the fossil record: stratophenomics and the generation of primary evolutionary-ecological data. Evol. Ecol. 26, 449–463, https://doi.org/10.1007/s10682-011-9511-9 (2012).
Pagel, M. Inferring the historical patterns of biological evolution. Nature 401, 877–884, https://doi.org/10.1038/44766 (1999).
Kidwell, S. M. & Holland, S. M. The quality of the fossil record: implications for evolutionary analyses. Annu. Rev. Ecol. Syst. 33, 561–588, https://doi.org/10.1146/annurev.ecolsys.33.030602.152151 (2002).
Rodríguez-Rey, M. et al. Criteria for assessing the quality of Middle Pleistocene to Holocene vertebrate fossil ages. Quat. Geochronol. 30, 69–79, https://doi.org/10.1016/j.quageo.2015.08.002 (2015).
Rodríguez-Rey, M. et al. A comprehensive database of quality-rated fossil ages for Sahul’s Quaternary vertebrates. Sci. Data 3, 160053, https://doi.org/10.1038/sdata.2016.53 (2016).
Rodríguez-Rey, M. et al. FosSahul database. ÆKOS Data Portal, https://doi.org/10.4227/05/56F077B3054E9 (2016).
Saltré, F. et al. Uncertainties in dating constrain model choice for inferring extinction time from fossil records. Quat. Sci. Rev. 112, 128–137, https://doi.org/10.1016/j.quascirev.2015.01.022 (2015).
Saltré, F. et al. Climate change not to blame for late Quaternary megafauna extinctions in Australia. Nat. Commun. 7, 10511, https://doi.org/10.1038/ncomms10511 (2016).
White, L. C., Saltré, F., Bradshaw, C. J. A. & Austin, J. J. High-quality fossil dates support a synchronous, Late Holocene extinction of devils and thylacines in mainland Australia. Biol. Lett. 14, https://doi.org/10.1098/rsbl.2017.0642 (2018).
Saltré, F. et al. Climate-human interaction associated with southeast Australian megafauna-extinction patterns. Nat. Commun. https://doi.org/10.1038/s41467-019-13277-0 (2019).
Block, S. et al. Where to dig for fossils: combining climate-envelope, taphonomy and discovery models. PloS ONE 11, e0151090, https://doi.org/10.1371/journal.pone.0151090 (2016).
Barnosky, A. D., Koch, P. L., Feranec, R. S., Wing, S. L. & Shabel, A. B. Assessing the causes of Late Pleistocene extinctions on the continents. Science 306, 70–75, https://doi.org/10.1126/science.1101476 (2004).
Kocsis, Á. T., Reddin, C. J., Alroy, J. & Kiessling, W. The R package divDyn for quantifying diversity dynamics using fossil sampling data. Methods. Ecol. Evol 10, 735–743, https://doi.org/10.1111/2041-210x.13161 (2019).
Goosse, H. et al. Description of the Earth system model of intermediate complexity LOVECLIM version 1.2. Geosci. Model Dev 3, 603–633, https://doi.org/10.5194/gmd-3-603-2010 (2010).
Peters, K. J. et al. FosSahul 2.0 database and R code. figshare, https://doi.org/10.6084/m9.figshare.8796944 (2019).
Johnson, C. N. et al. What caused extinction of the Pleistocene megafauna of Sahul? Proc. R. Soc. B 283, https://doi.org/10.1098/rspb.2015.2399 (2016).
Sandom, C., Faurby, S., Sandel, B. & Svenning, J.-C. Global late Quaternary megafauna extinctions linked to humans, not climate change. Proc. R. Soc. B 281, https://doi.org/10.1098/rspb.2013.3254 (2014).
Cooper, A. et al. Abrupt warming events drove Late Pleistocene Holarctic megafaunal turnover. Science 349, 602–606, https://doi.org/10.1126/science.aac4315 (2015).
Wroe, S. et al. Climate change frames debate over the extinction of megafauna in Sahul (Pleistocene Australia-New Guinea). Proc. Natl. Acad. Sci. U.S.A. 110, 8777–8781, https://doi.org/10.1073/pnas.1302698110 (2013).
Bird, M. I. et al. Humans, megafauna and environmental change in tropical Australia. J. Quat. Sci 28, 439–452, https://doi.org/10.1002/jqs.2639 (2013).
Allen, J. & O’Connell, J. Both half right: updating the evidence for dating first human arrivals in Sahul. Aust. Archaeol 79, 86–108, https://doi.org/10.1080/03122417.2014.11682025 (2014).
Jetz, W. & Fine, P. V. A. Global gradients in vertebrate diversity predicted by historical area-productivity dynamics and contemporary environment. PLoS Biol 10, e1001292, https://doi.org/10.1371/journal.pbio.1001292 (2012).
Fitterer, J. L., Nelson, T. A., Coops, N. C., Wulder, M. A. & Mahony, N. A. Exploring the ecological processes driving geographical patterns of breeding bird richness in British Columbia, Canada. Ecol. Appl. 23, 888–903, https://doi.org/10.1890/12-1225.1 (2013).
Stein, A., Gerstner, K. & Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol. Lett. 17, 866–880, https://doi.org/10.1111/ele.12277 (2014).
Kemp, C. W., Tibby, J., Arnold, L. J. & Barr, C. Australian hydroclimate during Marine Isotope Stage 3: a synthesis and review. Quat. Sci. Rev. 204, 94–104, https://doi.org/10.1016/j.quascirev.2018.11.016 (2019).
Cohen, T. J. et al. Hydrological transformation coincided with megafaunal extinction in central Australia. Geology 43, 195–198, https://doi.org/10.1130/G36346.1 (2015).
Dortch, J. et al. The timing and cause of megafauna mass deaths at Lancefield Swamp, south-eastern Australia. Quat. Sci. Rev. 145, 161–182, https://doi.org/10.1016/j.quascirev.2016.05.042 (2016).
Rakotoarinivo, M. et al. Palaeo-precipitation is a major determinant of palm species richness patterns across Madagascar: a tropical biodiversity hotspot. Proc. R. Soc. B 280, 20123048, https://doi.org/10.1098/rspb.2012.3048 (2013).
Tilman, D. & El Haddi, A. Drought and biodiversity in grasslands. Oecologia 89, 257–264, https://doi.org/10.1007/BF00317226 (1992).
Kay, R. F. & Madden, R. H. Mammals and rainfall: paleoecology of the middle Miocene at La Venta (Colombia, South America). J. Hum. Evol. 32, 161–199, https://doi.org/10.1006/jhev.1996.0104 (1997).
McCain, C. M. Could temperature and water availability drive elevational species richness patterns? A global case study for bats. Glob. Ecol. Biogeogr. 16, 1–13, https://doi.org/10.1111/j.1466-8238.2006.00263.x Ci (2007).
Hawkins, B. A. et al. Energy, water, and broad-scale geographic patterns of species richness. Ecology 84, 3105–3117, https://doi.org/10.1890/03-8006 (2003).
Dynesius, M. & Jansson, R. Evolutionary consequences of changes in species’ geographical distributions driven by Milankovitch climate oscillations. Proc. Natl. Acad. Sci. U.S.A. 97, 9115–9120, https://doi.org/10.1073/pnas.97.16.9115 (2000).
Fine, P. V. A. Ecological and evolutionary drivers of geographic variation in species diversity. Annu. Rev. Ecol. Evol. Syst. 46, 369–392, https://doi.org/10.1146/annurev-ecolsys-112414-054102 (2015).
Kaal, J., Schellekens, J., Nierop, K. G. J., Cortizas, A. M. & Muller, J. Contribution of organic matter molecular proxies to interpretation of the last 55 ka of the Lynch’s Crater record (NE Australia). Palaeogeogr. Palaeoclimatol. Palaeoecol 414, 20–31, https://doi.org/10.1016/j.palaeo.2014.07.040 (2014).
Timmermann, A. & Friedrich, T. Late Pleistocene climate drivers of early human migration. Nature 538, 92–95, https://doi.org/10.1038/nature19365 (2016).
Hogg, A. G. et al. SHCal13 southern hemisphere calibration, 0–50,000 years cal BP. Radiocarbon 55, 1889–1903, https://doi.org/10.2458/azu_js_rc.55.16783 (2013).
Bronk Ramsey, C. Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360, https://doi.org/10.1017/S0033822200033865 (2009).
Alroy, J. et al. Phanerozoic trends in the global diversity of marine invertebrates. Science 321, 97–100, https://doi.org/10.1126/science.1156963 (2008).
Loarie, S. R. et al. The velocity of climate change. Nature 462, 1052–1055, https://doi.org/10.1038/nature08649 (2009).
Bradshaw, C. J. A. et al. Minimum founding populations for the first peopling of Sahul. Nat. Ecol. Evol 3, 1057–1063, https://doi.org/10.1038/s41559-019-0902-6 (2019).
Acknowledgements
This research was done by the Australian Research Council Centre of Excellence for Australian Biodiversity and Heritage (project number CE170100015). ZJ and SU are the recipients of Australian Research Council Future Fellowships (project numbers FT150100138 and FT120100656) and RW the recipient of DECRA Fellowship DE150100070. We are grateful to S. Block and E. Bunney for their help with developing the algorithm.
Author information
Authors and Affiliations
Contributions
K.J.P., F.S. and C.J.A.B. conceptualized the paper, collected and analysed the data and wrote the manuscript. T.F., Z.J., R.W., M.M.D. and S.U. assisted with analyses and data collection. All authors edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Online-only Table
Online-only Table
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Peters, K.J., Saltré, F., Friedrich, T. et al. FosSahul 2.0, an updated database for the Late Quaternary fossil records of Sahul. Sci Data 6, 272 (2019). https://doi.org/10.1038/s41597-019-0267-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41597-019-0267-3
This article is cited by
-
Addendum: FosSahul 2.0, an updated database for the Late Quaternary fossil records of Sahul
Scientific Data (2021)
-
Extinction of eastern Sahul megafauna coincides with sustained environmental deterioration
Nature Communications (2020)
