
Abstract
The mitochondrial membrane-bound AAA protein Bcs1 translocate substrates across the mitochondrial inner membrane without previous unfolding. One substrate of Bcs1 is the iron–sulfur protein (ISP), a subunit of the respiratory Complex III. How Bcs1 translocates ISP across the membrane is unknown. Here we report structures of mouse Bcs1 in two different conformations, representing three nucleotide states. The apo and ADP-bound structures reveal a homo-heptamer and show a large putative substrate-binding cavity accessible to the matrix space. ATP binding drives a contraction of the cavity by concerted motion of the ATPase domains, which could push substrate across the membrane. Our findings shed light on the potential mechanism of translocating folded proteins across a membrane, offer insights into the assembly process of Complex III and allow mapping of human disease-associated mutations onto the Bcs1 structure.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others

Data availability
Atomic coordinates and structure factors have been deposited in the Protein Data Bank (PDB) with accession codes PDB 6U1Y (ΔNmBcs1-AMP-PNP) and PDB 6UKO (FLmBcs1-ADP). Cryo-EM structures and atomic models have been deposited in the Electron Microscopy Data Bank and PDB with accession codes EMDB 20808, PDB 6UKP (Apo mBcs1) and EMDB 20811, PDB 6UKS (ATPγS-bound mBcs1). Source data for Extended Data Fig. 1 are provided with the paper.
References
Twomey, E. C. et al. Substrate processing by the Cdc48 ATPase complex is initiated by ubiquitin unfolding. Science 365, eaax1033 (2019).
Cooney, I. et al. Structure of the Cdc48 segregase in the act of unfolding an authentic substrate. Science 365, 502–505 (2019).
Park, E. & Rapoport, T. A. Mechanisms of Sec61/SecY-mediated protein translocation across membranes. Annu. Rev. Biophys. 41, 21–40 (2012).
Letts, J. A. & Sazanov, L. A. Clarifying the supercomplex: the higher-order organization of the mitochondrial electron transport chain. Nat. Struct. Mol. Biol. 24, 800–808 (2017).
DiMauro, S. & Schon, E. A. Mitochondrial respiratory-chain diseases. N. Engl. J. Med. 348, 2656–2668 (2003).
Esser, L., Yu, C. A. & Xia, D. Structural basis of resistance to anti-cytochrome bc(1) complex inhibitors: implication for drug improvement. Curr. Pharm. Des. 20, 704–724 (2014).
Xia, D. et al. Crystal structure of the cytochrome bc1 complex from bovine heart mitochondria. Science 277, 60–66 (1997).
Hunte, C., Koepke, J., Lange, C., Rossmanith, T. & Michel, H. Structure at 2.3 A resolution of the cytochrome bc(1) complex from the yeast Saccharomyces cerevisiae co-crystallized with an antibody Fv fragment. Structure 8, 669–684 (2000).
Pfanner, N., Warscheid, B. & Wiedemann, N. Mitochondrial proteins: from biogenesis to functional networks. Nat. Rev. Mol. Cell Biol. 20, 267–284 (2019).
Cui, T. Z., Smith, P. M., Fox, J. L., Khalimonchuk, O. & Winge, D. R. Late-stage maturation of the Rieske Fe/S protein: Mzm1 stabilizes Rip1 but does not facilitate its translocation by the AAA ATPase Bcs1. Mol. Cell. Biol. 32, 4400–4409 (2012).
Wagener, N. & Neupert, W. Bcs1, a AAA protein of the mitochondria with a role in the biogenesis of the respiratory chain. J. Struct. Biol. 179, 121–125 (2012).
Esser, L. et al. Surface-modulated motion switch: capture and release of iron-sulfur protein in the cytochrome bc1 complex. Proc. Natl Acad. Sci. USA 103, 13045–13050 (2006).
Lill, R. Function and biogenesis of iron-sulphur proteins. Nature 460, 831–838 (2009).
Graham, L. A., Brandt, U., Sargent, J. S. & Trumpower, B. L. Mutational analysis of assembly and function of the iron-sulfur protein of the cytochrome bc1 complex in Saccharomyces cerevisiae. J. Bioenerg. Biomembr. 25, 245–257 (1993).
Brandt, U., Yu, L., Yu, C. A. & Trumpower, B. L. The mitochondrial targeting presequence of the Rieske iron-sulfur protein is processed in a single step after insertion into the cytochrome bc1 complex in mammals and retained as a subunit in the complex. J. Biol. Chem. 268, 8387–8390 (1993).
Cruciat, C. M., Hell, K., Folsch, H., Neupert, W. & Stuart, R. A. Bcs1p, an AAA-family member, is a chaperone for the assembly of the cytochrome bc(1) complex. EMBO J. 18, 5226–5233 (1999).
Nobrega, F. G., Nobrega, M. P. & Tzagoloff, A. BCS1, a novel gene required for the expression of functional Rieske iron-sulfur protein in Saccharomyces cerevisiae. EMBO J. 11, 3821–3829 (1992).
Ogura, T. & Wilkinson, A. J. AAA+ superfamily ATPases: common structure—diverse function. Genes Cells 6, 575–597 (2001).
Folsch, H., Guiard, B., Neupert, W. & Stuart, R. A. Internal targeting signal of the BCS1 protein: a novel mechanism of import into mitochondria. EMBO J. 15, 479–487 (1996).
Petruzzella, V. et al. Identification and characterization of human cDNAs specific to BCS1, PET112, SCO1, COX15, and COX11, five genes involved in the formation and function of the mitochondrial respiratory chain. Genomics 54, 494–504 (1998).
De Meirleir, L. et al. Clinical and diagnostic characteristics of complex III deficiency due to mutations in the BCS1L gene. Am J. Med. Genet. A. 121A, 126–131 (2003).
Fernandez-Vizarra, E. et al. Impaired complex III assembly associated with BCS1L gene mutations in isolated mitochondrial encephalopathy. Hum. Mol. Genet. 16, 1241–1252 (2007).
Holm, L. & Sander, C. Protein structure comparison by alignment of distance matrices. J. Mol. Biol. 233, 123–138 (1993).
Nouet, C., Truan, G., Mathieu, L. & Dujardin, G. Functional analysis of yeast bcs1 mutants highlights the role of Bcs1p-specific amino acids in the AAA domain. J. Mol. Biol. 388, 252–261 (2009).
Wagener, N., Ackermann, M., Funes, S. & Neupert, W. A pathway of protein translocation in mitochondria mediated by the AAA-ATPase Bcs1. Mol. Cell 44, 191–202 (2011).
Busch, A. & Waksman, G. Chaperone-usher pathways: diversity and pilus assembly mechanism. Philos. Trans. R. Soc. Lond. B. 367, 1112–1122 (2012).
Palmer, T. & Berks, B. C. The twin-arginine translocation (Tat) protein export pathway. Nat. Rev. Microbiol. 10, 483–496 (2012).
Kurisu, G., Zhang, H., Smith, J. L. & Cramer, W. A. Structure of the cytochrome b6f complex of oxygenic photosynthesis: tuning the cavity. Science 302, 1009–1014 (2003).
Frickey, T. & Lupas, A. N. Phylogenetic analysis of AAA proteins. J. Struct. Biol. 146, 2–10 (2004).
Gates, S. N. et al. Ratchet-like polypeptide translocation mechanism of the AAA+ disaggregase Hsp104. Science 357, 273–279 (2017).
Gao, Y. et al. Structures and operating principles of the replisome. Science 363, eaav7003 (2019).
Dong, Y. et al. Cryo-EM structures and dynamics of substrate-engaged human 26S proteasome. Nature 565, 49–55 (2019).
de la Pena, A. H., Goodall, E. A., Gates, S. N., Lander, G. C. & Martin, A. Substrate-engaged 26S proteasome structures reveal mechanisms for ATP-hydrolysis-driven translocation. Science 362, eaav0725 (2018).
Han, H., Monroe, N., Sundquist, W. I., Shen, P. S. & Hill, C. P. The AAA ATPase Vps4 binds ESCRT-III substrates through a repeating array of dipeptide-binding pockets. eLife 6, e31324 (2017).
Lo, Y. H. et al. Cryo-EM structure of the essential ribosome assembly AAA-ATPase Rix7. Nat. Commun. 10, 513 (2019).
Ramos-Arroyo, M. A. et al. Clinical and biochemical spectrum of mitochondrial complex III deficiency caused by mutations in the BCS1L gene. Clin. Genet. 75, 585–587 (2009).
Hinson, J. T. et al. Missense mutations in the BCS1L gene as a cause of the Bjornstad syndrome. N. Engl. J. Med. 356, 809–819 (2007).
Baker, R. A., Priestley, J. R. C., Wilstermann, A. M., Reese, K. J. & Mark, P. R. Clinical spectrum of BCS1L Mitopathies and their underlying structural relationships. Am. J. Med. Genet. A 179, 373–380 (2019).
Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallogr. D. 60, 2126–2132 (2004).
Smith, P. K. et al. Measurement of protein using bicinchoninic acid. Anal. Biochem. 150, 76–85 (1985).
Lanzetta, P. A., Alvarez, L. J., Reinach, P. S. & Candia, O. A. An improved assay for nanomole amounts of inorganic phosphate. Anal. Biochem. 100, 95–97 (1979).
Hess, H. H. & Derr, J. E. Assay of inorganic and organic phosphorus in the 0.1-5 nanomole range. Anal. Biochem. 63, 607–613 (1975).
Otwinowski, Z. & Minor, W. Processing of X-ray diffraction data collected in oscillation mode. Methods Enzymol. 276, 307–326 (1997).
Vagin, A. & Teplyakov, A. Molecular replacement with MOLREP. Acta Crystallogr. D. 66, 22–25 (2010).
Murshudov, G. N., Vagin, A. A. & Dodson, E. J. Refinement of macromolecular structures by the maximum-likelihood method. Acta Crystallogr. D. 53, 240–255 (1997).
Collaborative Computational Project, N. The CCP4 suite: programs for protein crystallography. Acta Crystallogr. D. 50, 760–763 (1994).
Adams, P. D. et al. PHENIX: building new software for automated crystallographic structure determination. Acta Crystallogr. D. 58, 1948–1954 (2002).
Meyerson, J. R. et al. Self-assembled monolayers improve protein distribution on holey carbon cryo-EM supports. Sci. Rep. 4, 7084 (2014).
Zheng, S. Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nat. Methods 14, 331–332 (2017).
Zhang, K. Gctf: real-time CTF determination and correction. J. Struct. Biol. 193, 1–12 (2016).
Zivanov, J. et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife 7, e42166 (2018).
Vilas, J. L. et al. MonoRes: automatic and accurate estimation of local resolution for electron microscopy maps. Structure 26, 337–344 e4 (2018).
Ramirez-Aportela, E. et al. Automatic local resolution-based sharpening of cryo-EM maps. Bioinformatics https://doi.org/10.1093/bioinformatics/btz671 (2019).
Pettersen, E. F. et al. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612 (2004).
Blazquez, A. et al. Infantile mitochondrial encephalomyopathy with unusual phenotype caused by a novel BCS1L mutation in an isolated complex III-deficient patient. Neuromuscul. Disord. 19, 143–146 (2009).
Visapaa, I. et al. GRACILE syndrome, a lethal metabolic disorder with iron overload, is caused by a point mutation in BCS1L. Am. J. Hum. Genet. 71, 863–876 (2002).
de Lonlay, P. et al. A mutant mitochondrial respiratory chain assembly protein causes complex III deficiency in patients with tubulopathy, encephalopathy and liver failure. Nat. Genet. 29, 57–60 (2001).
Leveen, P. et al. The GRACILE mutation introduced into Bcs1l causes postnatal complex III deficiency: a viable mouse model for mitochondrial hepatopathy. Hepatology 53, 437–447 (2011).
Acknowledgements
This research was supported by the intramural Research Program of the Center for Cancer Research, National Cancer Institute, National Institutes of Health. M.J.B. is supported by US National Institute of Environmental Health Sciences (grant no. ZIC ES103326 to M.J.B). We thank the staff members of the SER-CAT and GM/CA beamlines at the Advanced Photon Source, Argonne National Laboratory for beamline support. All DNA sequencing services was conducted at the Center for Cancer Research Genomics Core, National Cancer Institute and computation for the EM image reconstruction was carried out using the Biowulf Linux cluster (biowulf.nih.gov) at the National Institutes of Health, Bethesda. We thank G. Leiman for editorial assistance.
Author information
Authors and Affiliations
Contributions
W.K.T., M.J.B. and D.X. initiated the project. D.X. obtained funding. W.K.T. designed and conducted all the experiments. W.K.T., L.E. and D.X. analyzed the X-ray diffraction data and determined the structure. M.J.B., T.F. and N.V. acquired the EM images. W.K.T., M.J.B. and A.L.H. processed the EM images and reconstructed the maps. W.K.T. built the model into the EM maps. W.K.T. and D.X. interpreted the data and wrote the manuscript with input from all authors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Anke Sparmann was the primary editor on this article and managed its editorial process and peer review in collaboration with the rest of the editorial team.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Biochemical characterizations of recombinant mBcs1.
a, Size exclusion chromatographic profile of mBcs1 on Superdex 200. Inset: SDS-PAGE showing the purity of mBcs1 along the purification steps. b, One representative Michaelis Menten plot out of three experiments of mBcs1 ATPase activity to ATP concentration (n=3 technical replicates). c, Nucleotide-dependent conformational change of mBcs1 in the presence of different nucleotides as observed by Blue-Native PAGE.
Extended Data Fig. 2 Rotation function plots showing the presence of a proper 7-fold axis.
a, Full-length mBcs1 and b, ΔNmBcs1 crystals.
Extended Data Fig. 3 Data processing workflow for mBcs1 in ATPγS-bound form.
Particles of mBcs1 in the presence of ATPγS appear to form a mix of single and dual heptamer. Initial processing showed the two heptamers in the dual heptamer do not interact uniformly, hence reducing the resolutions. Therefore, only the class that contains single heptamers was selected for subsequent refinement. To improve resolution, the AAA-domain was masked for final refinement.
Extended Data Fig. 4 Single-particle cryo-EM analysis of mBcs1 in ATPγS-bound form.
a, Representative motion-corrected micrograph of an ATPγS-bound mBcs1 sample. b, Gold-standard Fourier shell correlation (FSC) curves of the final density map of the ATPγS-bound mBcs1. The reported resolution for this structure was based on the FSC=0.143 criterion. c, Local resolution map of ATPγS-bound mBcs1 structure at 4σ contour level. The color code is given by the bottom color strip. d, Angular distribution of particles used in the final 3D reconstruction of ATPγS-bound mBcs1. The height of the cylinder is proportional to the number of particles for that view. e, Map-to-model FSC.
Extended Data Fig. 5 Structural comparison of mBcs1 domains in different nucleotide states and example densities of bound ligands.
The ATPγS-bound mBcs1 structure is shown in colored ribbons (subunit A in cyan; subunit B in green) and subunits of the apo mBcs1 structure are shown in gray ribbons. a, Superposition of the Bcs1-specific domain of subunit A in the apo state (cyan) with the ATPγS-bound structure of the same domain (gray). The distance change in residue I49 of subunits B is indicated. b, Superposition of the structure of the AAA domain of subunit A in the apo state (cyan) with of the same structure in the ATPγS-bound state (gray)The distance change in residue R218 of subunits B is indicated. c, Conformational difference in a single mBcs1 subunit in the two nucleotide states. The distance change in residue K145 from the tip of the helix H2 is indicated. d, EM density map of ATPγS binding site (5.0σ level, gray mesh). mBcs1 is represented in blue cartoon. Walker-A K236 and ATPγS are shown as stick models with carbon, oxygen, nitrogen, phosphorous and sulphur atoms in blue, red, dark blue, orange and yellow, respectively. e, Difference Fourier map showing the bound AMP-PNP in the nucleotide-binding site (3.0σ level, gray mesh) in crystal structure of ΔNmBcs1. The structure of ΔNmBcs1 is represented by gray cartoon. Green sphere is Mg2+. Walker-A K236 and ATPγS are shown as stick models with carbon, oxygen, nitrogen and phosphorous atoms in gray, red, dark blue and orange, respectively. f, Difference Fourier map showing the bound ADP in the nucleotide-binding site (3.0σ level, cyan mesh) in crystal structure of full-length mBcs1. Residues E282, D283 and ADP are shown as stick models with carbon, oxygen, nitrogen and phosphorous atoms in gray, red, dark blue and magenta, respectively.
Extended Data Fig. 6 Data processing workflow for mBcs1 in apo form.
Additional details are provided in the Methods section.
Extended Data Fig. 7 Single-particle cryo-EM analysis of mBcs1 in apo form.
a, Representative motion-corrected micrograph of apo-mBcs1 sample. b, Gold-standard Fourier shell correlation (FSC) curves of the final density map for the apo mBcs1 structure. The reported resolution for this structure was based on the FSC=0.143 criterion. c, Local resolution map for the apo mBcs1 structure at 2.5σ level. The color code is given by the bottom color strip. d, Angular distribution of particles used in the final 3D reconstruction of apo mBcs1. The height of the cylinder is proportional to the number of particles for that view. e, Map-to-model FSC.
Extended Data Fig. 8 Superposition of structural domains of mBcs1.
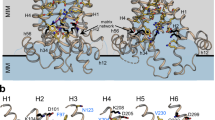
a, Superposition of the AAA domain from apo (gray) and ATPγS-bound (magenta) structures. b, Superposition of the Bcs1-specific domain from apo (gray) and ATPγS-bound (magenta) structures. c, Interface between two neighboring AAA domains of mBcs1 undergoes a sliding movement upon ATP binding. The structure in the vicinity of ATP binding site for the apo structure (both subunit A and B) is shown as cartoon in gray. The Arg-finger residue R343 of subunit A of the apo mBcs1 is shown as a stick model. The subunit A of the ATPγS-bound mBcs1 is shown in cyan and its neighboring subunit B is shown in green. The residue R343 of subunit A of the ATPγS-bound structure is shown as a stick model. The two structures are superimposed based on subunit B. The distances from CA atoms of R343 of apo or ATPγS-bound Bcs1 to the γ-phosphate of ATP are given. d, Interface between two neighboring AAA domains of bacterial NtrC1 in ADP- or ADP•BeF3-bound forms. Distances from CA atoms of Arg-finger residue R299 to bound ADP•BeF3 are given. e, Interface between two neighboring AAA domains of mammalian AAA protein p97 D1 domain in ADP- or ATPγS-bound forms. Distances from CA atoms of Arg-finger residue R299 to γ-phosphate of ATPγS are given.
Extended Data Fig. 9 Electrostatic surface potential of substrate and the putative substrate-binding cavity of mBcs1 in different states.
a, Electrostatic potential surface for the ISP subunit. b, Electrostatic potential surface of the apo mBcs1. The front portion of the surface was cut away to reveal the interior surface potential of the putative substrate-binding cavity. c, Electrostatic potential surface of the mBcs1 bound with ATPγS. The front portion of the surface was cut away to reveal the interior surface potential of the putative substrate-binding cavity.
Extended Data Fig. 10 Pathogenic mutations of BCS1L.
a, Pathogenic mutations of BCS1L and their locations in the Bcs1 structure. b–d, Mapping of the mutations on ATPγS-bound and apo mBcs1 in two orthogonal orientations: top view (top) and side view (bottom). TM, Bcs1-specific and AAA regions are in black, blue and magenta ribbons, respectively. Mutations are showed in spheres. Mutations found in (b), Björnstad syndrome; (c) GRACILE syndrome and (d) Complex III deficiency.
Supplementary information
Supplementary Video 1
Morphing of conformational transition from apo to ATPγS-bound state of mBcs1 in a concerted motion. Side view of mBcs1
Supplementary Video 2
Morphing of conformational transition from apo to ATPγS-bound state of mBcs1 in a concerted motion. View from the matrix side
Source data
Source Data Extended Data Fig. 1
Statistical source data
Rights and permissions
About this article

Cite this article
Tang, W.K., Borgnia, M.J., Hsu, A.L. et al. Structures of AAA protein translocase Bcs1 suggest translocation mechanism of a folded protein. Nat Struct Mol Biol 27, 202–209 (2020). https://doi.org/10.1038/s41594-020-0373-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41594-020-0373-0
This article is cited by
-
A concerted ATPase cycle of the protein transporter AAA-ATPase Bcs1
Nature Communications (2023)
-
ncRNAs-mediated high expression of TIMM8A correlates with poor prognosis and act as an oncogene in breast cancer
Cancer Cell International (2022)
-
Cryo-EM structure of transmembrane AAA+ protease FtsH in the ADP state
Communications Biology (2022)
-
The assembly, regulation and function of the mitochondrial respiratory chain
Nature Reviews Molecular Cell Biology (2022)
-
The Pseudomonas aeruginosa substrate-binding protein Ttg2D functions as a general glycerophospholipid transporter across the periplasm
Communications Biology (2021)
