
Abstract
The growth of telomerase-deficient cancers depends on the alternative lengthening of telomeres (ALT), a homology-directed telomere-maintenance pathway. ALT telomeres exhibit a unique chromatin environment and generally lack the nucleosome remodeler ATRX, pointing to an epigenetic basis for ALT. Recently, we identified a protective role for the ATRX-interacting macroH2A1.2 histone variant during homologous recombination and replication stress (RS). Consistent with an inherent susceptibility to RS, we show that human ALT telomeres are highly enriched for macroH2A1.2. However, in contrast to ATRX-proficient cells, ALT telomeres transiently lose macroH2A1.2 during acute RS to facilitate DNA double-strand break (DSB) formation, a process that is almost completely prevented by ectopic ATRX expression. Telomeric macroH2A1.2 is re-deposited in a DNA damage response (DDR)-dependent manner to promote homologous recombination-associated ALT pathways. Our findings thus identify the dynamic exchange of macroH2A1.2 on chromatin as an epigenetic link among ATRX loss, RS-induced DDR initiation and telomere maintenance via homologous recombination.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$189.00 per year
only $15.75 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others

Data availability
Raw data and image files are available upon request.
References
O’Sullivan, R. J. & Almouzni, G. Assembly of telomeric chromatin to create ALTernative endings. Trends Cell Biol. 24, 675–685 (2014).
Pickett, H. A. & Reddel, R. R. Molecular mechanisms of activity and derepression of alternative lengthening of telomeres. Nat. Struct. Mol. Biol. 22, 875–880 (2015).
Dilley, R. L. & Greenberg, R. A. ALTernative telomere maintenance and cancer. Trends Cancer 1, 145–156 (2015).
Heaphy, C. M. et al. Altered telomeres in tumors with ATRX and DAXX mutations. Science 333, 425 (2011).
Lovejoy, C. A. et al. Loss of ATRX, genome instability, and an altered DNA damage response are hallmarks of the alternative lengthening of telomeres pathway. PLoS Genet. 8, e1002772 (2012).
Clynes, D. et al. Suppression of the alternative lengthening of telomere pathway by the chromatin remodelling factor ATRX. Nat. Commun. 6, 7538 (2015).
Napier, C. E. et al. ATRX represses alternative lengthening of telomeres. Oncotarget 6, 16543–16558 (2015).
Nguyen, D. T. et al. The chromatin remodelling factor ATRX suppresses R-loops in transcribed telomeric repeats. EMBO Rep. 18, 914–928 (2017).
O’Sullivan, R. J. et al. Rapid induction of alternative lengthening of telomeres by depletion of the histone chaperone ASF1. Nat. Struct. Mol. Biol. 21, 167–174 (2014).
Leung, J. W. et al. Alpha thalassemia/mental retardation syndrome X-linked gene product ATRX is required for proper replication restart and cellular resistance to replication stress. J. Biol. Chem. 288, 6342–6350 (2013).
Goldberg, A. D. et al. Distinct factors control histone variant H3.3 localization at specific genomic regions. Cell 140, 678–691 (2010).
Ratnakumar, K. et al. ATRX-mediated chromatin association of histone variant macroH2A1 regulates alpha-globin expression. Genes Dev. 26, 433–438 (2012).
Wong, L. H. et al. ATRX interacts with H3.3 in maintaining telomere structural integrity in pluripotent embryonic stem cells. Genome Res. 20, 351–360 (2010).
Khurana, S. et al. A macrohistone variant links dynamic chromatin compaction to BRCA1-dependent genome maintenance. Cell Rep. 8, 1049–1062 (2014).
Kustatscher, G., Hothorn, M., Pugieux, C., Scheffzek, K. & Ladurner, A. G. Splicing regulates NAD metabolite binding to histone macroH2A. Nat. Struct. Mol. Biol. 12, 624–625 (2005).
Kim, J. et al. Replication stress shapes a protective chromatin environment across fragile genomic regions. Mol. Cell 69, 36–47 (2018).
Bouwman, P. et al. 53BP1 loss rescues BRCA1 deficiency and is associated with triple-negative and BRCA-mutated breast cancers. Nat. Struct. Mol. Biol. 17, 688–695 (2010).
Bunting, S. F. et al. 53BP1 inhibits homologous recombination in Brca1-deficient cells by blocking resection of DNA breaks. Cell 141, 243–254 (2010).
Xu, Y. et al. 53BP1 and BRCA1 control pathway choice for stalled replication restart. eLife 6, e30523 (2017).
Dilley, R. L. et al. Break-induced telomere synthesis underlies alternative telomere maintenance. Nature 539, 54–58 (2016).
Porro, A., Feuerhahn, S., Reichenbach, P. & Lingner, J. Molecular dissection of telomeric repeat-containing RNA biogenesis unveils the presence of distinct and multiple regulatory pathways. Mol. Cell. Biol. 30, 4808–4817 (2010).
Henson, J. D. et al. DNA C-circles are specific and quantifiable markers of alternative-lengthening-of-telomeres activity. Nat. Biotechnol. 27, 1181–1185 (2009).
Poole, L. A. et al. SMARCAL1 maintains telomere integrity during DNA replication. Proc. Natl Acad. Sci. USA 112, 14864–14869 (2015).
Bailey, S. M., Brenneman, M. A. & Goodwin, E. H. Frequent recombination in telomeric DNA may extend the proliferative life of telomerase-negative cells. Nucleic Acids Res. 32, 3743–3751 (2004).
Nabetani, A., Yokoyama, O. & Ishikawa, F. Localization of hRad9, hHus1, hRad1, and hRad17 and caffeine-sensitive DNA replication at the alternative lengthening of telomeres-associated promyelocytic leukemia body. J. Biol. Chem. 279, 25849–25857 (2004).
Cho, N. W., Dilley, R. L., Lampson, M. A. & Greenberg, R. A. Interchromosomal homology searches drive directional ALT telomere movement and synapsis. Cell 159, 108–121 (2014).
Pan, X. et al. FANCM, BRCA1, and BLM cooperatively resolve the replication stress at the ALT telomeres. Proc. Natl Acad. Sci. USA 114, E5940–E5949 (2017).
Ramamoorthy, M. & Smith, S. Loss of ATRX suppresses resolution of telomere cohesion to control recombination in ALT cancer cells. Cancer Cell 28, 357–369 (2015).
Conomos, D., Reddel, R. R. & Pickett, H. A. NuRD-ZNF827 recruitment to telomeres creates a molecular scaffold for homologous recombination. Nat. Struct. Mol. Biol. 21, 760–770 (2014).
Juhasz, S., Elbakry, A., Mathes, A. & Lobrich, M. ATRX promotes DNA repair synthesis and sister chromatid exchange during homologous recombination. Mol. Cell 71, 11–24 (2018).
Clynes, D. et al. ATRX dysfunction induces replication defects in primary mouse cells. PLoS ONE 9, e92915 (2014).
Huh, M. S. et al. Compromised genomic integrity impedes muscle growth after Atrx inactivation. J. Clin. Invest. 122, 4412–4423 (2012).
Watson, L. A. et al. Atrx deficiency induces telomere dysfunction, endocrine defects, and reduced life span. J. Clin. Invest. 123, 2049–2063 (2013).
Ishov, A. M., Vladimirova, O. V. & Maul, G. G. Heterochromatin and ND10 are cell-cycle regulated and phosphorylation-dependent alternate nuclear sites of the transcription repressor Daxx and SWI/SNF protein ATRX. J. Cell. Sci. 117, 3807–3820 (2004).
Berube, N. G., Smeenk, C. A. & Picketts, D. J. Cell cycle-dependent phosphorylation of the ATRX protein correlates with changes in nuclear matrix and chromatin association. Hum. Mol. Genet. 9, 539–547 (2000).
Minocherhomji, S. et al. Replication stress activates DNA repair synthesis in mitosis. Nature 528, 286–290 (2015).
Buschbeck, M. & Hake, S. B. Variants of core histones and their roles in cell fate decisions, development and cancer. Nat. Rev. Mol. Cell Biol. 18, 299–314 (2017).
Kim, J. K., Oberdoerffer, P. & Khurana, S. The histone variant macroH2A1 is a splicing-modulated caretaker of genome integrity and tumor growth. Mol. Cell. Oncol. 5, e1441629 (2018).
Wan, B. et al. SLX4 assembles a telomere maintenance toolkit by bridging multiple endonucleases with telomeres. Cell Rep. 4, 861–869 (2013).
Henson, J. D. et al. The C-Circle Assay for alternative-lengthening-of-telomeres activity. Methods 114, 74–84 (2017).
Ruthenburg, A. J. et al. Recognition of a mononucleosomal histone modification pattern by BPTF via multivalent interactions. Cell 145, 692–706 (2011).
Acknowledgements
We thank J. Cooper for critical reading of the manuscript, T. Karpova for imaging support and R.A. Greenberg for reagents. R.J.G. acknowledges support from the Medical Research Council, grant no. MC_UU_12025/unit program MC_UU_12009/3. This work was supported by the National Institutes of Health (NIH) training grant no.T32 GM007491–38 (P.D.R.) and grant no. R01 CA155232 (M.J.G.), and the Intramural Research Programs of the NIH National Cancer Institute, Center for Cancer Research and the National Institute on Aging.
Author information
Authors and Affiliations
Contributions
J.K., Y.L. and P.O. conceived the project. J.K. performed ChIP, western blot and cell growth analyses. C.S. performed dot plots, Q-FISH, T-SCE and TIF assay, with help from K.W. and P.J.C. A.D.T. performed immunofluorescence analyses, P.D.R. performed in vitro affinity purification. R.J.G. contributed U2OS-ATRX cells. P.O. wrote the manuscript with help from Y.L., M.J.G. and J.K.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Integrated supplementary information
Supplementary Figure 1 MacroH2A1.2 function in telomere maintenance.
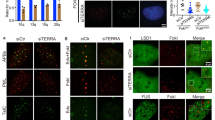
(a) Dot blot for telomere DNA content in the indicated ChIP samples or input DNA from U2OS cells. (b) Dot blot for Alu repeat DNA content in indicated ChIP samples or input DNA from U2OS cells analyzed in Fig. 1b. Equal amounts of DNA were loaded for all samples. A quantification relative to input is shown in Fig. 1b. Triplicates reflect three independent IP experiments. (c) Western blot analysis of macroH2A1.2 knockdown efficiency in the indicated cell lines expressing a control siRNA (si-ctrl), two independent siRNAs against macroH2A1.2 (si-1.2–1 and si-1.2–2) or an siRNA against the macroH2A1.1 splice variant (si-1.1). Lysates were harvested 72 h post transfection and analyzed by immunoblotting with the indicated antibodies. (d – f) Telomere Q-FISH in the indicated cell lines expressing either si-ctrl, si-1.2–1 or si-1.2–2. Samples were collected 72 h post transfection and analyzed as described in Fig. 1c. *** p < 0.001, on the basis of one-way ANOVA. (g) C-circle analysis in GM847 cells expressing a control shRNA (sh-RFP), or two distinct shRNAs against macroH2A1.2 (sh-1.2–1, sh-1.2–2). Values are expressed as mean and s.e.m. (n = 3), * p < 0.01, ** p < 0.01 on the basis of Student’s two-tailed t-test.
Supplementary Figure 2 MacroH2A1.2 promotes homology-directed DSB repair at telomeres.
(a) T-SCE frequency in U2OS cells expressing the indicated siRNAs. T-SCE was measured and analyzed as in Fig. 2a, this experiment represents an independent replicate of Fig. 2a. (b) BRCA1 accumulation at TRF1-FokI induced DSBs in U2OS cells 50 h after transfection with a control siRNA (si-ctrl, n = 251 cells) or si-macroH2A1.2 (si-1.2–1, n = 180 cells). Representative IF images are shown; scale bar, 10 μm. The percentage of BRCA1+ foci was determined based on a greater than two-fold increase in BRCA1 intensity at TRF1-FokI foci relative to total nuclear intensity, two-fold cutoff value is based on expression of catalytic dead TRFI-FokI-D450A (see Supplementary Fig. 2f). ** denotes p < 0.001 by Mann-Whitney U test. One of six independent experiments is shown. (c) Fraction of γ-H2AX+ TRF1-FokI foci as defined in Supplementary Fig. 2b, f in the presence of the indicated siRNAs, values are expressed as mean and s.e.m. (n = 6). ** p < 0.01, based on Student’s paired, two-tailed t-test. (d) BRCA1 accumulation at γ-H2AX+ TRF1-FokI foci quantified as in (b). (e) Fraction of BRCA1+ γ-H2AX+ TRF1-FokI foci as defined in (d). Values are expressed as mean and s.e.m., * p < 0.01 based on Student’s paired, two-tailed t-test. (f) BRCA1 and γ-H2AX accumulation at TRF1-FokI-induced DSBs in U2OS cells expressing TRF1-FokI WT or the D450A catalytic mutant. (g) BRCA1 accumulation at TRF1-FokI induced DSBs in U2OS cells overexpressing eGFP (n = 53 cells) or macroH2A1.2 (n = 69 cells) analyzed as in (b), one of three representative experiments is shown, see Fig. 2j for mean and s.e.m. Overexpression was confirmed by Western blot analysis. (h) BRCA1 accumulation at TRF1-FokI foci as in (b) in cells expressing si-ctrl or an siRNA against macroH2A1.1 (si-1.1). The fraction of BRCA1+ TRF1-FokI foci is shown, values are expressed as mean and s.e.m. (n = 3). (i) U2OS cell-based HR reporter assay, where GFP+ cells represent cells having undergone HR. HR efficiency was measured by fluorescence activated cell scanning (FACS) 72 h post transfection with the indicated siRNAs. The assay was performed as described previously (Cell Rep. 8, 1049–1062, 2014).
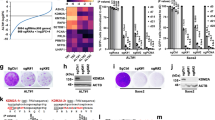
Supplementary Figure 3 Replication stress and ATRX modulate macroH2A1.2 levels at telomeres.
(a) H2B ChIP at three distinct subtelomeric loci in the presence (dark colors) or absence of HU treatment (light colors) in U2OS cells carrying a Dox-inducible ATRX transgene. Cells were either treated with vehicle (control) or Dox (ATRX) before HU treatment. Enrichment relative to input is shown, values are expressed as mean and s.d. (n = 3). (b) MacroH2A1.2 ChIP in U2OS cells at the indicated loci in the absence (gray bars) or presence of Aph (black bars). NFS; non-fragile region. Enrichment relative to input is shown, values are expressed as mean and s.d. (n = 3). (c) MacroH2A1.2 ChIP at a control locus (NFS-1) or subtelomeric loci on chromosomes 10 and 15 in the indicated cell lines cells treated with (+) or without HU (–). Enrichment relative to untreated samples is shown for each locus. Values are expressed as mean and s.d. (n = 3). (d) Dot blot for telomeric DNA content in the indicated ChIP samples or input DNA from ALT-positive SaOS-2 cells analyzed in (c). (e) Dot blot analysis for telomeric DNA content in the indicated ChIP samples or input DNA from ALT-negative K562 cells in the presence or absence of ATRX knockdown treated with DMSO (–) or HU (+). A quantification of telomeric C-probe signal (TelC) in macroH2A1.2 IP relative to input is shown, values are expressed as mean and s.e.m. (n = 3). * < p < 0.05 based on Student’s two-tailed t-test. (f) Western blot analysis of the indicated proteins in K562 cells expressing a control shRNA or an shRNA against ATRX. (g) Dot blot analysis for telomeric DNA content in the indicated ChIP samples or input DNA from Fig. 3e. ATMi/ATRi: ATM/ATR inhibitor.
Supplementary Figure 4 MacroH2A1.2 loss impairs ALT tumor cell growth.
(a) ChIP for macroH2A1.2 at the indicated loci in K562 cells in the presence (WT) or absence of macroH2A1.2 (1.2 CRISPR-KO). Values are normalized to input and expressed as mean and s.d. (n = 3). (b, c) Population doubling (PD) analyses in ALT-positive U2OS cells (b) or SaOS-2 cells (c) infected with either sh-RFP or one of two independent macroH2A1.2 sh-RNAs (sh-1.2–1 or sh-1.2–2), followed by serial passaging. Cumulative PDs were calculated using the formula “3.32 (log Xe – log Xb) + S” in which Xb is the inoculum cell number, Xe the cell harvest number at the end of the incubation time, and S is the starting PD. PDs were then plotted for the indicated days. Values are expressed as mean and s.d. (n = 3). A Western blot analysis of macroH2A1.2 knockdown efficiency in SaOS-2 cells is shown (d). (e, f) PD analyses for ALT-negative K562 cells (e) or 293T cells (f), performed as described above.
Supplementary information
Supplementary Figures, Supplementary Tables and Supplementary Dataset
Supplementary Figures 1–4, Supplementary Tables 1–2 and Supplementary Dataset 1
Rights and permissions
About this article

Cite this article
Kim, J., Sun, C., Tran, A.D. et al. The macroH2A1.2 histone variant links ATRX loss to alternative telomere lengthening. Nat Struct Mol Biol 26, 213–219 (2019). https://doi.org/10.1038/s41594-019-0192-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41594-019-0192-3
This article is cited by
-
Alternative lengthening of telomeres (ALT) cells viability is dependent on C-rich telomeric RNAs
Nature Communications (2023)
-
Histone dynamics during DNA replication stress
Journal of Biomedical Science (2021)
-
Imaging the response to DNA damage in heterochromatin domains reveals core principles of heterochromatin maintenance
Nature Communications (2021)
-
The roles of histone variants in fine-tuning chromatin organization and function
Nature Reviews Molecular Cell Biology (2020)
-
Histone H2A variants alpha1-extension helix directs RNF168-mediated ubiquitination
Nature Communications (2020)
