
Abstract
Breastfeeding profoundly shapes the infant gut microbiota, which is critical for early life immune development, and the gut microbiota can impact host physiology in various ways, such as through the production of metabolites. However, few breastmilk-dependent microbial metabolites mediating host–microbiota interactions are currently known. Here, we demonstrate that breastmilk-promoted Bifidobacterium species convert aromatic amino acids (tryptophan, phenylalanine and tyrosine) into their respective aromatic lactic acids (indolelactic acid, phenyllactic acid and 4-hydroxyphenyllactic acid) via a previously unrecognized aromatic lactate dehydrogenase (ALDH). The ability of Bifidobacterium species to convert aromatic amino acids to their lactic acid derivatives was confirmed using monocolonized mice. Longitudinal profiling of the faecal microbiota composition and metabolome of Danish infants (n = 25), from birth until 6 months of age, showed that faecal concentrations of aromatic lactic acids are correlated positively with the abundance of human milk oligosaccharide-degrading Bifidobacterium species containing the ALDH, including Bifidobacterium longum, B. breve and B. bifidum. We further demonstrate that faecal concentrations of Bifidobacterium-derived indolelactic acid are associated with the capacity of these samples to activate in vitro the aryl hydrocarbon receptor (AhR), a receptor important for controlling intestinal homoeostasis and immune responses. Finally, we show that indolelactic acid modulates ex vivo immune responses of human CD4+ T cells and monocytes in a dose-dependent manner by acting as an agonist of both the AhR and hydroxycarboxylic acid receptor 3 (HCA3). Our findings reveal that breastmilk-promoted Bifidobacterium species produce aromatic lactic acids in the gut of infants and suggest that these microbial metabolites may impact immune function in early life.
Similar content being viewed by others

Main
Human breastmilk is a well-adapted nutritional supply for the infant1. Breastfeeding provides the infant with important short-term protection against infections and may also provide long-term metabolic and immunological benefits1,2,3. These benefits may partly be mediated through the gut microbiota, since breastfeeding is the strongest determinant of gut microbiota composition and function during infancy4,5,6. Human breastmilk contains human milk oligosaccharides (HMOs), which are complex, highly abundant sugars serving as substrates for specific microbes including certain species of Bifidobacterium7. This co-evolution between bifidobacteria and the host, mediated by HMOs, to a large extent directs the colonization of the gut in early life, which has critical impact on the immune system8. Depletion of specific microbes, including Bifidobacterium, in early life has been associated with increased risk of allergy and asthma development in childhood9,10 and has been suggested to compromise immune function and lead to increased susceptibility to infectious disease11,12. Despite Bifidobacterium dominating the gut of breastfed infants and being widely acknowledged as beneficial, mechanistic insights into the contribution of these bacteria and their metabolites to immune function and development are limited and have mainly focused on short-chain fatty acids12,13. Recent studies show that microbial aromatic amino acid metabolites including tryptophan-derived indoles14 via activation of the aryl hydrocarbon receptor (AhR)15,16 can fortify the intestinal barrier17,18, protect against pathogenic infections15,19 and influence host metabolism17,20,21, making this group of microbial metabolites of particular interest in the context of early life.
Here, we show that breastmilk-promoted Bifidobacterium species, via a previously unrecognized aromatic lactate dehydrogenase (ALDH), produce aromatic lactic acids including indolelactic acid (ILA) in substantial amounts in the infant gut and that faecal concentrations of this metabolite are correlated with the capacity of infant faeces to activate AhR. We furthermore demonstrate that ILA via AhR and hydroxycarboxylic acid receptor 3 (HCA3)-dependent pathways impact immune functions ex vivo, suggesting that breastmilk-promoted Bifidobacterium via production of aromatic lactic acids impact the immune system in early life.
Results
Bifidobacterium associate with aromatic lactic acids in the infant gut
To explore interactions between breastfeeding status, gut microbial composition and metabolism of aromatic amino acids in early life, we used 16S ribosomal RNA amplicon sequencing to infer gut microbiota composition and a targeted ultraperformance liquid chromatography–mass spectrometry (UPLC–MS) metabolomics approach to quantify 19 aromatic amino acids and derivatives thereof (Supplementary Tables 1 and 2) in faecal samples from 59 healthy Danish infants from the SKOT I cohort22. The SKOT I infants included were born full term, 9.1 ± 0.3 (mean ± s.d.) months of age at sampling and 40.7% were still partially breastfed (Supplementary Data 1a,b). After stratification of the 9-month-old infants on the basis of breastfeeding status (partially breastfed versus weaned), principal coordinates analysis (PCoA) of weighted UniFrac distances showed a significant separation across the first PC-axis (r2 = 0.093, P < 0.001, Adonis test; Fig. 1a), which mirrored an increasing gradient in relative abundance of Bifidobacterium (r2 = 0.397, P < 0.001, Adonis test; Fig. 1b). Other metadata (age, gender, mode of delivery, current formula intake and age of introduction to solid foods) did not explain gut microbiota variation to the same degree as breastfeeding status (r2 < 0.05, P > 0.03, Adonis tests; Supplementary Data 1c) and no bacterial genera differed significantly according to these parameters (FDR-corrected P > 0.1, Mann–Whitney U-tests; Supplementary Data 1d). Principal component analysis (PCA) of faecal aromatic amino acid metabolite concentrations (Supplementary Data 1e) also suggested a minor separation by breastfeeding status, which was largely driven by three aromatic lactic acids, 4-hydroxyphenyllactic acid (4-OH-PLA), phenyllactic acid (PLA) and indolelactic acid (ILA) (Fig. 1c). Correlation analysis revealed that Bifidobacterium, but no other bacterial genera, were significantly associated with faecal concentrations of all three aromatic lactic acids (4-OH-PLA, PLA and ILA), in addition to indolealdehyde (IAld) (Fig. 1d and Supplementary Data 1f). The Bifidobacterium species (Extended Data Fig. 1a and Supplementary Data 1g) enriched in the breastfed infants, B. longum, B. bifidum and B. breve, were positively associated with the faecal concentrations of aromatic lactic acids (4-OH-PLA, PLA and ILA) and IAld (cluster 1 in Fig. 1e) but negatively associated with the faecal concentrations of aromatic propionic acids, aromatic amino acids and, to a lesser degree, with aromatic acetic acids (cluster 2 in Fig. 1e). In contrast, postweaning type Bifidobacterium species, including B. adolescentis, B. animalis/pseudolongum and B. catenulatum group23,24, were not significantly associated with aromatic lactic acids nor breastfeeding status (Fig. 1e). These associations were in agreement with the observation that the concentrations of the three aromatic lactic acids were higher in the faeces of breastfed than weaned infants (Extended Data Fig. 1b). Furthermore, the abundances of the three aromatic lactic acids in infant urine (Supplementary Figs. 1–3) showed similar positive associations with relative abundances of breastmilk-promoted Bifidobacterium species (Extended Data Fig. 1c). In addition, faecal and urinary levels of ILA were positively correlated (ρ = 0.68, P < 0.0001), showing that faecal levels of this metabolite are reflected systemically. Consistently, urine abundance of ILA, but not of PLA and 4-OH-PLA, were significantly higher in breastfed compared to weaned infants (Extended Data Fig. 1b). Together, this suggests that specific Bifidobacterium species produce aromatic lactic acids in the infant gut (Fig. 1f).

a,b, PCoA plots of weighted UniFrac distances based on OTUs from faecal samples of 9-month-old infants participating in the SKOT cohort (n = 59). a, Samples are coloured according to breastfeeding status, with ellipses indicating 80% confidence interval (CI) of datapoints for partially breastfed (red, n = 24) and weaned (blue, n = 35) infants. b, Samples are coloured according to relative abundance of the genus Bifidobacterium. c, PCA plot of concentrations (nmol g–1 faeces) of aromatic amino acids and their derivatives in SKOT faecal samples, coloured according to breastfeeding status, with ellipses indicating 80% CI of datapoints for breastfed (red, n = 24) and weaned (blue, n = 35) infants. Loadings (correlations between variables and the principal components) are shown with arrows, with annotations of the aromatic amino acids ILA, 4-OH-PLA and PLA shown. d, Heatmap illustrating Spearman’s rank correlation coefficients (two-sided tests) between the relative abundance of Bifidobacterium and concentrations of aromatic amino acids and their derivatives in SKOT faecal samples (n = 59). e, Heatmap illustrating hierarchical clustering (dendrogram on the right side) of Spearman’s rank correlation coefficients (two-sided tests) between the relative abundance of the different Bifidobacterium species and selected microbial-derived aromatic amino acid catabolites in SKOT faecal samples (n = 59). Box and whiskers plots are showing relative abundance (line, median; box, interquartile range (IQR); whiskers, minimum to maximum) of the Bifidobacterium species, stratified according to breastfeeding status, with statistical significance evaluated by two-sided Mann–Whitney U-test. f, The pathway of aromatic amino acid catabolism by gut microbes (modified from Smith and Macfarlane110, Smith and Macfarlane111 and Zelante et al.15). For all panels, asterisks indicate statistical significance: *P < 0.05, **P < 0.01, ***P < 0.001 and ****P < 0.0001.
Bifidobacterium species produce aromatic lactic acids in vitro
To confirm the ability of Bifidobacterium species detected in infants to produce aromatic lactic acids, Bifidobacterium type strains were grown anaerobically in a medium containing all three aromatic amino acids with either glucose or HMOs as sole carbohydrate sources. Analyses of culture supernatants revealed that ILA, PLA and 4-OH-PLA were produced mainly by B. bifidum, B. breve, B. longum ssp. longum, B. longum ssp. infantis and B. scardovii (Fig. 2a), in accordance with the associations observed in the 9-month-old infants (Fig. 1e). Other Bifidobacterium species, namely B. adolescentis, B. animalis ssp. lactis, B. animalis ssp. animalis, B. dentium, B. catenulatum, B. pseudocatenulatum and B. pseudolongum ssp. pseudolongum produced only low amounts of these metabolites (Fig. 2a). The ability of Bifidobacterium species to produce high levels of the aromatic lactic acids was generally convergent with the ability to use HMOs as a carbohydrate source (Fig. 2a), suggesting a link between breastmilk-promoted bifidobacteria and production of aromatic lactic acids. None of the downstream products of the aromatic lactic acids (Fig. 1f) was detected in any of the culture supernatants.

a, In vitro production of ILA, PLA and 4-OH-PLA by Bifidobacterium species type strains in modified MRS medium (MRSc) with 2% (w/v) glucose or a mix of HMOs as sole carbohydrate source. For the type strains of B. adolescentis, B. animalis ssp. animalis, B. animalis ssp. lactis, B. dentium and B. catenulatum, no or very poor growth (OD600nm < 0.4) was observed with HMOs as the carbohydrate source. Means of three biological replicates are shown. b, Neighbour-joining phylogenetic tree of all genes in the Bifidobacterium species type strains annotated as LDH-encoding genes (ldh). The four clusters are designated types 1–4. c, Production of ILA, PLA and 4-OH-PLA by E. coli LMG194 cells transformed with an inducible vector lacking (empty vector) or containing the type 4 ldh (Type4_ldh+) from B. longum ssp. infantis DSM 20088 in LB medium 5 h post-induction of gene expression by addition of l-arabinose and supplementation with the aromatic pyruvic acids (indolepyruvic acid, phenylpyruvic acid and 4-hydroxyphenylpyruvic acid). Bars show mean ± s.d. of three biological replicates. d, Growth curves of B. longum ssp. longum 105-A (WT), its isogenic insertional type 4 ldh mutant (Type4_ldh mutant) and the type 4 ldh mutant strain complemented with the type 4 ldh gene (complemented). Curves show mean ± s.d. of three biological replicates and doubling times reported as mean ± s.d.. e, Production of ILA, PLA, 4-OH-PLA and lactic acid by wild type, Type4_ldh mutant and the complemented strain in early stationary phase cultures as indicated in d. Bars show mean ± s.d. of three biological replicates. Statistical significance was evaluated by one-way ANOVA. ND, not detected.
Identification of a responsible ALDH
Since it has been reported that a lactate dehydrogenase (LDH) in Lactobacillus species can convert phenylpyruvic acid to PLA25, we hypothesized that a corresponding enzyme would be present in Bifidobacterium species. Alignment and phylogenetic analysis of all genes annotated as ldh in the Bifidobacterium type strains included in this study, revealed four clusters (Fig. 2b). Whereas all Bifidobacterium genomes contain the ldh (here designated as type 1 ldh) responsible for conversion of pyruvic acid to lactic acid in the bifidobacterial fructose 6-phosphate shunt26,27, some species have an extra ldh, here designated as type 2, type 3 and type 4, respectively. In agreement with the in vitro fermentations (Fig. 2a), all prominent aromatic lactic acid-producing Bifidobacterium species contain the type 4 ldh, suggesting that this could encode a previously unrecognized ALDH. A further analysis of all available whole-genome sequenced Bifidobacterium strains showed that the type 4 ldh is universally present in B. longum, B. bifidum, B. breve and B. scardovii strains (Supplementary Table 3). Interestingly, genomic analysis of the Bifidobacterium type strains revealed that the type 4 ldh gene is part of a genetic element containing an amino acid transaminase gene (suspected to be responsible for converting the aromatic amino acids into aromatic pyruvic acids) and a haloacid dehalogenase gene (of unknown importance) (Supplementary Fig. 4), which has been indicated to constitute an operon in B. breve28. Cloning of the type 4 ldh gene from the type strain of B. longum ssp. infantis (DSM20088) into a vector transformed into Escherichia coli revealed that the expression of the type 4 ldh gene indeed resulted in the appearance of PLA, 4-OH-PLA and ILA in the culture supernatant (Fig. 2c). To verify the type 4 ldh-dependent production of aromatic lactic acids in Bifidobacterium species, we generated a type 4 ldh insertional mutant strain by homologous recombination in B. longum ssp. longum 105-A (Supplementary Fig. 5), a genetically tractable strain29,30 containing the type 4 ldh (Supplementary Fig. 6a). The type 4 LDH amino acid sequence of the 105-A strain had >98% identity to the homologues in type strains of B. longum ssp. longum and B. longum ssp. infantis and >91% identity to B. bifidum, B. breve and B. scardovii (Supplementary Fig. 6b) but no non-bifidobacterial homologues were found by BLAST analysis (amino acid sequence identity cutoff 60%). Cultivation of the wild type (WT), the type 4 ldh mutant strain and a complemented type 4 ldh mutant strain in a medium containing the three aromatic amino acids confirmed that type 4 ldh disruption did not impair growth in a rich medium (Fig. 2d). ILA, PLA and 4-OH-PLA accumulated in the supernatant of the WT and of the complemented type 4 ldh mutant strains but not in the type 4 ldh mutant (Fig. 2e). Importantly, the type 4 ldh mutant was not significantly compromised in its ability to convert pyruvic acid to lactic acid (Fig. 2e), supporting the distinct role of type 4 ldh in converting aromatic pyruvic acids. Further, to demonstrate in vivo production of the indicated aromatic lactic acids, we monocolonized germ-free mice with either the WT or the type 4 ldh mutant strain and found a 20–60-fold increase in their concentrations in WT versus type 4 ldh mutant monocolonized mice (Extended Data Fig. 2). Purification and characterization of the recombinant type 4 LDH enzyme revealed that it had a mass of 33.9 kDa (Supplementary Fig. 7a), while the native molecular mass was estimated to be 71.9 kDa by size exclusion chromatography, indicating dimer formation in solution (Supplementary Fig. 7b). Lack of added metal ions or addition of ethylenediaminetetraacetic acid (EDTA) did not reduce enzymatic activity, the optimal pH was 8.0–8.5 and the enzyme was most stable at 37 °C (Supplementary Fig. 7c–e). Heterotrophic effects were neither observed for fructose 1,6-bisphosphate (an allosteric effector for type 1 LDH) nor for several intermediates for aromatic amino acid synthesis26,27 (Supplementary Fig. 8). However, we found that phosphate served as a positive effector, suggesting that type 4 LDH is an intracellular enzyme (Supplementary Fig. 9a,b). Assay performed at the different phosphate concentrations revealed the type 4 LDH is a K-type allosteric enzyme (Supplementary Fig. 9b). The catalytic rate (kcat) was moderate to high for the aromatic pyruvic acid substrates but very low for pyruvic acid (Fig. 3), in accordance with the non-impaired lactic acid production observed for the type 4 ldh mutant (Fig. 2e). Production of ILA, PLA and 4-OH-PLA from the respective aromatic pyruvic acid substrates was verified by high-performance liquid chromatography (HPLC) (Supplementary Fig. 9c). The enzyme showed highest affinity (lowest K0.5) for indolepyruvic acid but highest catalytic rate for 4-hydroxyphenylpyruvic acid in the presence of 100 mM phosphate (Fig. 3). However, the catalytic efficiency (kcat/K0.5) was highest for indolepyruvic acid (194 s−1 mM−1), followed by 4-hydroxyphenylpyruvic acid (16 s−1 mM−1) and phenylpyruvic acid (11 s−1 mM−1), suggesting preference for indolepyruvic acid. The observed Hill coefficient (nH = 1.0–1.4) for all substrates indicate weak positive cooperativity under the conditions tested. Collectively, these results show that the type 4 ldh gene (from now on denoted aldh) encodes an ALDH responsible for the production of ILA, PLA and 4-OH-PLA in Bifidobacterium species associated with breastfeeding.

a–d, Substrate saturation curves of the type 4 LDH obtained for indolepyruvic acid (a), phenylpyruvic acid (b), 4-hydroxyphenylpyruvic acid (c) and pyruvic acid (d). Kinetic parameters, which were calculated by curve-fitting two independent experimental data to the Hill equation, are shown in the insets. The reaction was carried out in the presence of 100 mM phosphate.
Bifidobacterium species govern aromatic lactic acid profiles during early infancy
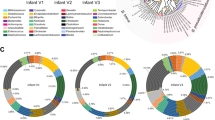
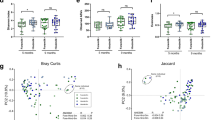
To study the dynamics of Bifidobacterium species establishment and aromatic lactic acids in early infancy, we established the Copenhagen Infant Gut (CIG) cohort including 25 healthy breastfed or mixed-fed infants, sampled every 2–4 weeks from birth until the age of 6 months (Supplementary Data 2a,b) for microbiome profiling (Supplementary Data 2c and Extended Data Fig. 3) and targeted metabolite quantification including aromatic lactic acids (Supplementary Table 4 and Supplementary Data 2d). The subject-specific gut microbiota profiles revealed a highly individualized species composition (Extended Data Fig. 3c and Extended Data Fig. 4a). As expected from a cohort mainly containing vaginally born, breastfed infants, the gut microbiota was highly dominated by Bifidobacterium (average of 64.2%) and among the top ten dominating taxa, B. longum (38.5%), B. breve (9.1%), B. bifidum (7.9%), B. catenulatum group (6.4%) and B. dentium (1.7%) were found (Extended Data Fig. 3a,b), with the remaining Bifidobacterium species being assigned to B. scardovii (0.24%), B. adolescentis (0.15%) and B. animalis/pseudolongum (0.10%) (Supplementary Data 2e). PCoA using Bray–Curtis dissimilarities, revealed a separation of the communities across samples based on relative abundance of the five dominating Bifidobacterium species, B. longum, B. bifidum, B. breve, B. catenulatum group and B. dentium (Fig. 4a and Extended Data Fig. 4b–f). Community abundance of B. longum, B. bifidum and B. breve but not B. catenulatum group and B. dentium (Fig. 4a) matched the measured faecal concentrations of the aromatic lactic acids (Fig. 4b). On the basis of quantitative PCR (qPCR) estimated total bacterial load of all samples, we calculated absolute abundances of each bacterial taxon in the 16S rRNA amplicon dataset and defined infant-type Bifidobacterium species as the summarized abundance of B. longum, B. bifidum, B. breve and B. scardovii. We observed a significant increase in the absolute abundance of infant-type Bifidobacterium species from birth to around 6 months of age and this occurred concurrently with a progressive increase in the faecal concentrations of ILA, PLA and 4-OH-PLA and a progressive decrease in faecal abundances of HMO residuals (Extended Data Fig. 5). We confirmed by linear mixed models31 adjusting for subject and age that the absolute abundances of the infant-type Bifidobacterium species were positively associated with faecal levels of ILA, PLA and 4-OH-PLA and additionally negatively associated with abundances of HMOs residuals in faeces (Fig. 4c). Among all bacterial taxa detected, B. longum, B. bifidum and B. breve were most strongly associated with faecal levels of ILA, PLA and 4-OH-PLA (Supplementary Data 2g). These associations were also evident within individuals when using repeated measures correlations32 (Extended Data Fig. 6) and across individuals at each sampling point using Spearman’s rank correlations (Extended Data Fig. 7). Furthermore, re-analysing the microbiome data at the amplicon sequence variant (ASV) level showed very similar results (Extended Data Fig. 8a). Finally, qPCR targeting B. longum ssp. longum, B. longum ssp. infantis, B. breve and B. bifidum confirmed the associations to aromatic lactic acids and HMOs (Extended Data Fig. 8b). Notably, we found that both subspecies of B. longum were associated with the aromatic lactic acids but mainly B. longum ssp. infantis and B. bifidum were associated with the HMO residuals in faeces (Extended Data Fig. 8b). To further corroborate our findings regarding the relevance in early life and impact of breastfeeding, we mined a published metagenomic dataset from faecal samples from a cohort of 98 Swedish mother–infants pairs5 for bifidobacterial metagenome-assembled genomes (MAGs) containing the aldh gene. This analysis revealed a significantly higher abundance of aldh-containing MAGs in exclusively breastfed (compared to mixed- or formula-fed) infants at 4 months and in partially breastfed (compared to weaned) infants at 12 months of age (Extended Data Fig. 9). In addition, we found very low abundance of aldh-containing MAGs in the mothers and a significant decline of these MAGs in infants after introduction to solid foods (4 versus 12 months of age). We have thus established a link between breastfeeding, degradation of HMOs, abundance of specific Bifidobacterium species and concentrations of aromatic lactic acids in early infancy.

a,b, PCoA plots of Bray–Curtis dissimilarities (n = 234 (i)), coloured according to relative abundance of Bifidobacterium (a) or log10-transformed concentration (nmol per g of faeces) of aromatic lactic acids (sum of ILA, PLA and 4-OH-PLA) (b) in faeces of infants participating in the CIG cohort. Dashed lined circles include communities dominated (relative abundance >50%) by B. longum, B. bifidum, B. breve, B. catenulatum group or B. dentium (B. adolescentis, B. scardovii and B. animalis/pseudolongum never dominated any of the communities; Extended Data Figs. 3 and 4). (i) Six samples were omitted from the analyses due to low read counts (<8,000), and one sample was omitted due to no available metabolomics data. c, Heatmap illustrating linear mixed-model coefficients (two-sided test, adjusted for subject and age) between the absolute abundance of Bifidobacterium species (cells per g of faeces) and faecal concentrations (nmol per g of faeces) of aromatic lactic acids (ILA, PLA and 4-OH-PLA, n = 240 (ii)) or faecal relative abundances of HMOs (2′FL/3FL, 2′/3-O-fucosyllactose; 3′SL/6′SL, 3′/6′-O-sialyllactose; LNT/LNnT, lacto-N-tetraose/lacto-N-neotetraose; n = 228 (iii)) in the CIG cohort. Infant-type Bifidobacterium species is the sum of absolute abundances of B. longum, B. breve, B. bifidum and B. scardovii. Statistical significance was evaluated by FDR-corrected P values indicated by asterisks with *P < 0.05, **P < 0.01, ***P < 0.001 and ****P < 0.0001. (ii) One sample was omitted due to no available metabolomics data. (iii) Twelve samples were omitted due to the infants no longer being breastfed and one due to no available metabolomics data. d–f, Absolute abundance of Bifidobacterium species (average relative abundance >1% of total community) and concentrations of ILA, PLA and 4-OH-PLA in faeces of selected individuals from the CIG cohort. Values of bacterial counts <106 cells per g of faeces and metabolite concentrations <1 nmol per g of faeces are not shown. Summarized absolute abundance of infant-type Bifidobacterium species is indicated with grey background shading. d, Breastfed infants colonized early with infant-type Bifidobacterium species and with concurrent high concentrations of ILA, PLA and 4-OH-PLA through the first 6 months of life. e, Infants colonized late with infant-type Bifidobacterium species and with concurrent low concentrations of ILA, PLA and 4-OH-PLA. f, Infants with recorded oral antibiotics intake during the first 6 months of life. Similar dynamics of the remaining infants can be seen in Extended Data Fig. 10.
Examination of the Bifidobacterium and aromatic lactic acid dynamics in each of the 25 infants during the first 6 months of life (Fig. 4d–f, Extended Data Fig. 10 and Supplementary Fig. 10) revealed that breastfed infants that were early colonized by infant-type Bifidobacterium species (n = 17) consistently showed high concentrations of aromatic lactic acids in faeces (Fig. 4d and Extended Data Fig. 10a). In contrast, infants with delayed infant-type Bifidobacterium species colonization (n = 5) showed considerably lower concentrations of the aromatic lactic acids, in particular of ILA, despite breastfeeding (Fig. 4e and Extended Data Fig. 10b). We noticed among the latter, infants CIG08 and CIG09 were twins, born late preterm and dominated by an operational taxonomic unit (OTU) assigned to Clostridium neonatale (Extended Data Fig. 3c and Supplementary Data 2c) in accordance with previous reports on C. neonatale overgrowth33 and delayed Bifidobacterium colonization34,35,36,37 in preterm infants. Infant CIG07 who also showed delayed colonization with infant-type Bifidobacterium, was mixed-fed throughout the whole period and predominantly colonized with E. coli and Clostridium species (Extended Data Fig. 3c). Infant CIG18 had relatively low faecal concentrations of aromatic lactic acids until age 172 days, when B. breve replaced B. dentium (Fig. 4e), consistent with the fact that B. dentium lacks the aldh gene while B. breve contains it (Fig. 2b and Supplementary Table 3). Finally, in the three infants treated with antibiotics during our study, Bifidobacterium species abundances were temporarily decreased simultaneously with reduced concentrations of the aromatic lactic acids (Fig. 4f). Together, these results demonstrate that HMO-using infant-type Bifidobacterium species determine the abundance of aromatic lactic acids in the infant gut. Yet, the impact of early/late Bifidobacterium colonization, preterm delivery, exposure to antibiotics and formula supplementation with respect to bifidobacterial aromatic lactic acid production warrants further investigation.
Indolelactate modulates immune responses via AhR and HCA3
The tryptophan-derived metabolite ILA was consistently measured in the faeces of breastfed infants at 0–6 months (Supplementary Table 4) and 9 months of age (Supplementary Table 2). Microbial tryptophan catabolites have been found to contribute to intestinal and systemic homoeostasis, in particular by their ability to bind the AhR14. Furthermore, aromatic lactic acids have been found to activate HCA3 (ref. 38), which is involved in the regulation of immune function and energy homoeostasis39,40. In accordance with previous reports15,16, we observed modest but significant dose-dependent increases in agonistic activity of ILA in both rat and human AhR reporter gene cell lines (Supplementary Fig. 11). Furthermore, all three aromatic amino lactic acids, and especially ILA, showed very potent and dose-dependent agonistic activity towards the HCA3 in a reporter cell line assay (Supplementary Fig. 12), in agreement with previous reports38,40. To investigate the relationship between gut microbiota, aromatic amino acid metabolites and AhR signalling, the AhR activity induced by sterile-filtered faecal water from selected CIG infants (Fig. 4d–f) was associated with the most abundant bacterial taxa (Fig. 5a) and all quantified aromatic amino acid metabolites (n = 19) in the same samples (Fig. 5b). This revealed that, among other taxa, in particular the infant-type Bifidobacterium species were positively associated with AhR activity across individuals using linear mixed models adjusted for subject and age (Fig. 5a) as well as within individuals using repeated measures correlations (Supplementary Fig. 13a). Of all the aromatic amino acid metabolites measured, only faecal concentrations of ILA were significantly positively associated with AhR activity (Fig. 5b and Supplementary Fig. 13b).

a,b, Scatter plot of subject- and age-adjusted linear mixed-model β coefficients (versus FDR-corrected P values, two-sided tests) obtained from associations between AhR activity of faecal water (n = 111) from selected CIG infants (Fig. 4d–f, n = 11) in a reporter cell line assay and absolute abundance of faecal bacterial taxa (relative abundance >0.1%, n = 40, circles) (a) or quantities of aromatic amino acid metabolites (n = 19, triangles) (b) measured in the same samples. Coloured circles/triangles mark taxa/metabolite measures that are significantly positively (red) or negatively (blue) associated with AhR activity within an FDR-corrected P of 0.1 (dashed line). ILA, indolelactic acid; Tyr, tyrosine.
Since ILA on absorption in the gut is circulated in the body18, we next asked whether ILA affects immune function via AhR and HCA3. Since the human AhR has adapted to sense microbial tryptophan catabolites41 and only humans and other hominids contain HCA3 (ref. 38), we isolated immune cells from human blood and assessed the impact of ILA on their function. Specifically, we cultured isolated human CD4+ T cells under TH17-polarizing conditions and assessed IL-22 production on exposure to ILA. Interestingly, ILA induced the production of IL-22, an effector cytokine produced by TH17 cells after AhR stimulation42,43,44, in a dose-dependent manner (Fig. 6a). Conversely, the addition of AhR antagonist CH-223191 inhibited IL-22 production, further corroborating that ILA acts through AhR to induce IL-22 production (Fig. 6b). We also isolated monocytes from human blood, where both AhR (ref. 45) and HCA3 (ref. 46) are expressed, stimulated the cells with E. coli lipopolysaccharide (LPS) and interferon-gamma (IFN-γ) to induce pro-inflammatory conditions, and assessed IL-12p70 production on ILA exposure. ILA reduced pro-inflammatory IL-12p70 production in a dose-dependent manner (Fig. 6c). Addition of CH-223191 blocked the ILA-induced inhibition of IL-12p70 production, confirming that ILA also acts through AhR in human monocytes (Fig. 6d). Furthermore, ILA-induced inhibition of IL-12p70 was prevented, when using knockdown of HCA3 by small interfering RNA (siRNA), supporting that ILA also acts as an anti-inflammatory agent via HCA3 in human monocytes (Fig. 6e). Thus, ILA affects human immune responses via AhR and HCA3-dependent pathways, suggesting that Bifidobacterium-derived ILA is a highly relevant AhR and HCA3 agonist that may impact immune responses in early life.

a, Fold change in IL-22 production in purified human CD4+ T cells cultured for 3 d under TH17-polarizing conditions in the presence of ILA at 5, 50 and 200 µM as compared to vehicle (DMSO control). ILA5µm versus vehicle, P = 0.54; ILA50µm versus vehicle, P = 0.031; ILA200µm versus vehicle, P = 0.0098. b, Fold changes over vehicle (DMSO control) in IL-22 production in human purified CD4+ T cells cultured under TH17-polarizing conditions in the presence of 200 µM ILA with and without the AhR antagonist CH-223191. ILA200µm versus ILA200µm + AhRant, P = 0.0005. c, Fold change in IL-12p70 production in purified human monocytes stimulated with LPS and IFN-γ in the presence of ILA at 5, 50 and 200 µM as compared to vehicle (DMSO control). ILA5µm versus vehicle, P = 0.27; ILA50µm versus vehicle, P = 0.041; ILA200µm versus vehicle, P = 0.031. d, Fold changes over vehicle (DMSO control) in IL-12p70 production in purified human monocytes stimulated with LPS and IFN-γ in the presence of 200 µM ILA with and without the AhR antagonist CH-223191. ILA200µm versus ILA200µm + AhRant, P = 0.033. e, Fold changes over vehicle (DMSO control) in IL-12p70 production in purified human monocytes stimulated with LPS and IFN-γ in the presence of 200 µM ILA and ScrRNA or HCA3 siRNA. ILA200µm + ScrRNA versus ILA200µm + HCA3 siRNA, P = 0.027. For all panels, bars show mean ± s.d.. Each dot represents data from an individual donor (n = 4–6), and, for both T cells and monocytes, the measurements are derived from two-to-three independent experiments. Statistical significance was evaluated by two-sided paired (a and c) or two-sided unpaired (b, d and e) t-tests (with Welch’s correction for b and asterisks indicating *P < 0.05, **P < 0.01, ***P < 0.001. For a and c, the absolute cytokine values were compared between ILA- versus vehicle-treated cells. For b, d and e, the fold changes over vehicle (ratios) were compared between ILA- and ILA + antagonist/siRNA-treated cells. Dashed lines indicate a fold change over vehicle of 1.
Discussion
The importance of intestinal commensal bacteria in regulation of the intestinal barrier function and immune development during infancy is well established47,48. Yet, specifically the symbiotic role of the breastmilk-promoted Bifidobacterium species, which are highly abundant in many breastfed infants, remains largely unknown. Here, we identified an ALDH, which catalyses the last step of the conversion of aromatic amino acids into their respective aromatic lactic acids in the infant gut. This ALDH was different in terms of amino acid sequence compared to the ALDH previously identified in Clostridiales species (C. sporogenes, C. cadaveris and P. anaerobius)18. Furthermore, these species are not prevalent nor abundant in the infant gut and have previously been found to convert the aromatic lactic acids into aromatic propionic acids as end products18. We show that only the infant-type Bifidobacterium species, adapted to breastfeeding by their HMO-transport and degradation genes providing them with a colonization advantage in infant gut13,49,50,51,52, contain the ALDH gene. This fits the observation that Bifidobacterium species commonly isolated from the infant gut in vitro produce relative higher levels of ILA compared with adult- or animal-associated Bifidobacterium species53. Our enzymatic assays showed strong adaptation of ALDH towards indolepyruvic acid, resulting preferentially in the formation of ILA. Importantly, our data suggest that the production of the AhR agonist ILA by breastmilk-promoted Bifidobacterium is a key determinant of AhR-dependent signalling in the gut during infancy. Previous studies have found that ILA decreases inflammation in intestinal cells through activation of AhR54,55. Here, we show ex vivo that ILA impacts human primary immune cells via AhR- and HCA3-dependent pathways. The observed dose- and AhR-dependent stimulation of IL-22 secretion by ILA may reflect a mechanism by which infant-type Bifidobacterium species impact intestinal homoeostasis in early life, as IL-22 for example provides protection against gastrointestinal pathogens56,57,58 and promotes mucus production57,59 and epithelial function60. Further, the AhR- and HCA3-dependent inhibitory effect of ILA on IL-12p70 secretion by monocytes may constitute a means by which infant-type Bifidobacterium species contribute to the regulation of the pro-inflammatory responses to LPS derived from Enterobacteriaceae species that also often colonize the neonatal/infant gut. While the functional implications of aromatic lactic acids remain to be fully characterized, the phenomenon observed here is probably of fundamental importance, since HCA3, which is only expressed in humans and other hominids38, is involved in the regulation of immune functions and energy homoeostasis39,40. Furthermore, AhR signalling is involved in protection against gastrointestinal pathogens15 and plays a key role in enhancement of intestinal barrier function61,62, immune development19,63,64,65, attenuation of induced colitis66, autoimmunity67,68,69 and metabolic syndrome61. In addition, ILA and PLA have been shown in vitro to have direct antibacterial70,71 and antifungal properties72,73. Therefore, our findings provide a rationale for further investigation of the implications of aromatic lactic acids in infant health and immune development.
Methods
Human study populations and metadata
SKOT cohort
The discovery cohort consisted of a random subset of 59 healthy infants (30 male, 29 female) of the observational SKOT I cohort22. No statistical method was used to predetermine sample size. The infants were originally recruited from Copenhagen and Frederiksberg regions by random selection from the National Danish Civil Registry74. Inclusion criteria were single birth and full-term delivery, absence of chronic illness and age of 9 months ± 2 weeks at inclusion. Mode of delivery, gender, age at sampling, use of medication, breast- and formula-feeding prevalence, as well as exclusive and total breastfeeding duration and age of introduction to solid foods were recorded by parental questionnaires (Supplementary Data 1a,b). Anthropometrics, full dietary assessment and other relevant metadata have been published previously4,75. Faecal samples were obtained at 9 months ± 2 weeks of age and were stored at –80 °C until DNA extraction, as described previously4. Urine samples were collected by the use of cotton balls placed in the infants’ disposable nappies from which the urine was squeezed into a sterile tube and stored at –80 °C. In cases of faeces in the nappy, the urine sample was discarded. The study protocol was approved by the Committees on Biomedical Research Ethics for the Capital Region of Denmark (H-KF-2007-0003) and The Data Protection Agency (2002-54-0938, 2007-54-026) approved the study. Informed consent was obtained from all parents of infants participating in the SKOT I study. The parents did not receive any compensation.
CIG cohort
The validation cohort, CIG, consisted of 25 healthy infants (12 male, 13 female), mostly vaginally born (23/25) and full-term (23/25) delivered. No statistical method was used to predetermine sample size. However, on the basis of the original observations using the SKOT cohort, we estimated that 25 infants with multiple time points would be sufficient to demonstrate the dynamics between Bifidobacterium species and aromatic lactic acids. Infants in CIG were recruited through social media and limited to the Copenhagen region. Parents collected faecal samples approximately every second week, starting from the first week of life until 6 months of age (within weeks 0, 2, 4, 6, 8, 10, 12, 16, 20 and 24), ending with a total of 269 samples. Parents were instructed to collect faecal samples from nappies into sterile faeces collection tubes (Sarstedt) and immediately store them at –18 °C in a home freezer until transportation to the Technical University of Denmark where the samples were stored at –80 °C until sample preparation. Gender, preterm versus full-term birth, mode of delivery, infant/maternal antibiotics, feeding patterns (breastmilk versus formula) and introduction to solid foods were recorded (Supplementary Data 2a,b). The Data Protection Agency (18/02459) approved the study. The Committees on Biomedical Research Ethics for the Capital Region of Denmark confirmed that the CIG study was not notifiable according to the Act on Research Ethics Review of Health Research Projects (paragraph 1, subsection 4), as the study only concerned the faecal microbial composition and activity and not the health of the children. Informed consent was obtained from all parents of infants participating in the CIG study. In addition, parents of twins gave informed consent to publish data from the twins although the parents themselves would be able to identify their children using indirect identifiers. The parents did not receive any compensation.
Gut microbiota analysis
16S rRNA gene amplicon sequencing
Sample preparation and sequencing were performed as previously described4 using a subset of 59 faecal samples originating from infants participating in the SKOT I cohort and 241 faecal samples from 25 infants participating in the CIG cohort (data from a total of 28 samples were missing due to insufficient sample material (n = 1), insufficient DNA extraction/lack of PCR product (n = 20), very low number of sequencing reads (n = 6) or resemblance of community to sequenced blank buffer DNA extraction negative controls (n = 1)). Briefly, DNA was extracted from 250 mg of faeces or blank buffer negative controls (PowerLyzer PowerSoil DNA isolation kit, MoBio 12855-100) and the V3 region of the 16S rRNA gene was amplified (30 s at 98 °C, 24–30 cycles of 15 s at 98 °C and 30 s at 72 °C, followed by 5 min at 72 °C) using non-degenerate universal barcoded primers including sequencing adaptors (Supplementary Table 5)76 and then sequenced with the Ion OneTouch and Ion PGM platform with a 318-Chip v.2. Sequences from SKOT and CIG were analysed separately. Briefly, they were demultiplexed according to barcode and trimmed as previously described76,77 in CLC Genomic Workbench (v.8.5 CLCbio, Qiagen). Quality filtering (-fastq_filter, MAX_EE(SKOT) =2.0, MAX_EE(CIG) =1.0), dereplication, OTU clustering (-cluster_otus, minsize 4), chimaera filtering (-uchime_ref, RDP v.9 database), mapping of reads to OTUs (-usearch_global, id 97%) and generation of OTU tables (python, uc2otutab.py) were done according to the UPARSE pipeline78. In QIIME (ref. 79), OTU tables (nOTUs(SKOT) = 545, nOTUs(CIG) = 478) were filtered to include only OTUs with abundance across all samples above 0.005% of the total OTU counts (nOTUs(SKOT) = 258, nOTUs(CIG) = 145), ending up with 21,781 ± 13,110 (mean ± s.d.) reads for SKOT I and 40,156 ± 17,614 (mean ± s.d.) reads for CIG. OTU relative abundances within samples were then estimated by total sum scaling. Taxonomy was assigned to the OTUs using the rdp classifier with confidence threshold 0.5 (ref. 80) and the GreenGenes database v.13.8 (ref. 81). Estimating species composition in the CIG cohort, the OTUs detected with identical taxonomy were collapsed and using a cutoff of average relative abundance of 0.1%, only 39 bacterial species/taxa remained, representing 97.5% of total community (Supplementary Data 2c and Extended Data Fig. 3c). On the basis of PyNAST alignment of representative OTU sequences from each cohort separately, a phylogenetic tree was created with FastTree, as described previously77. Alpha diversity (Shannon index) and beta diversity (weighted and unweighted UniFrac distances, abundance weighted and binary Bray–Curtis were calculated in QIIME, with the sequencing depth rarefied to 2,000 (SKOT)/8,000 (CIG) sequences per sample. To investigate Bifidobacterium species composition, OTU sequences classified as Bifidobacterium according to the GreenGenes database v.13.8 were filtered to remove low-abundant OTUs (cutoff 0.1% of total Bifidobacterium) and the taxonomy of these resulting OTUs (nOTUs(SKOT) = 23, nOTUs(CIG) = 8) was confirmed by BLAST (ref. 82) search against the 16S rRNA gene sequence database at the National Center for Biotechnology Information (NCBI). The top BLAST hit indicated species annotation (Supplementary Data 1g and 2e). OTUs were collapsed into Bifidobacterium species (B. longum, B. bifidum, B. breve, B. catenulatum group, B. adolescentis, B. scardovii, B. dentium and B. animalis/pseudolongum) on the basis of the top BLAST hit (Supplementary Data 1g and 2e). In addition, to validate the findings from the OTU analysis, we performed ASV analysis on the CIG cohort samples using the DADA2 pipeline v.1.14 (ref. 83) with the demultiplexed and trimmed reads and the same cutoffs as for the OTU analysis (MAX_EE = 1, ASVs filtered to include only those with abundance across all samples above 0.005% of the total ASV counts), resulting in a total of 211 ASVs and 13 ASVs assigned to Bifidobacterium (using the RDP database v.18) above the abundance cutoff of 0.1% of the total Bifidobacterium population (Supplementary Data 2f). As for OTUs, the taxonomy for individual Bifidobacterium ASVs was confirmed by BLAST search against the 16S rRNA gene sequence database at NCBI. Infant-type Bifidobacterium species were defined as the summarized abundance of B. longum, B. bifidum, B. breve and B. scardovii. CIG individuals were grouped on the basis of colonization with infant-type Bifidobacterium species, into those with early colonization (colonized within first month reaching average relative abundance >40% during first 6 months, n = 17) and late colonization (not detectable or on average <0.5% of total community within the first 3 months of life, n = 5), as well as those associated with antibiotics (at least one episode of recorded oral antibiotics during the first 6 months of life, n = 3).
Quantitative PCR
Total bacterial load (universal primers) and absolute abundances of B. longum ssp. longum, B. longum ssp. infantis (subspecies-specific primers), B. bifidum and B. breve (species-specific primers) were estimated by qPCR, using the primers listed in Supplementary Table 5. Each reaction was performed (in triplicates) with 5 µl of PCR-grade water, 1.5 µl of forward and reverse primer, 10 µl of SYBR Green I Master 2X (LightCycler 480 SYBR Green I Master, Roche) and 2 µl of template DNA, in a total volume of 20 µl. Standard curves were generated from tenfold serial dilutions of linearized plasmid (containing 108–100 gene copies per µl), constructed by cloning a PCR-amplified 199-base pair (bp) fragment of the 16S rRNA gene (V3 region) of E. coli (ATCC 25922) or a 307-bp fragment of the Blon0915 gene84 of B. longum ssp. infantis (DSM 20088) or a 301-bp fragment of the BL0274 gene85 of B. longum ssp. longum (DSM 20219) into a pCR4-Blunt-TOPO (Invitrogen) or pCRII-Blunt-TOPO vector (Invitrogen). For B. bifidum (16S rRNA gene) and B. breve (groEL gene), tenfold serial dilutions of DNA (containing 107–100 gene copies per µl) extracted from pure cultures of the type strains (DSM 20456 and DSM 20213) were used for standard curves. Plates were run on the LightCycler 480 Instrument II (Roche) with the programme including 5 min of pre-incubation at 95 °C, followed by 45 cycles with 15 s at 95 °C, 15 s at 50–72 °C and 15 s at 72 °C and a subsequent melting curve analysis including 5 min at 95 °C, 1 min at 65 °C and continuous temperature increase (ramp rate 0.11 °C s–1) until 98 °C. Data were analysed with the LightCycler 480 Software (v.1.5) (Roche). Bacterial load data (using the universal primers) were used to estimate absolute abundances of each microbial taxa by multiplying with relative abundances derived from 16S rRNA gene amplicon sequencing.
Bifidobacterium strains and growth experiments
Aromatic lactic acid production by Bifidobacterium type strains
Bifidobacterium type strains (Supplementary Table 6) were cultivated on MRSc (MRS containing 2% (w/v) glucose and supplemented with 0.05% (w/v) l-cysteine) agar plates for 48 h at 37 °C anaerobically. Single colonies were dissolved in 5.0 ml of prereduced MRSc broth and incubated for 24 h at 37 °C anaerobically with shake. The overnight (ON) cultures were washed (10,000g, room temperature, 5 min) and resuspended in sterile 0.9% NaCl water, diluted 1:20 (in triplicates) in prereduced MRSc or MRSc + HMOs (MRS broth without glucose but supplemented with 2.0% (w/v) HMO mixture and 0.05% (w/v) l-cysteine) and re-incubated at 37 °C anaerobically for 72 h, after which optical density OD600nm was measured and the culture supernatants (16,000g, 5 min, 4 °C) were analysed by UPLC–MS. The individual HMOs were kindly donated by Glycom A/S; 2′-O-fucosyllactose (2′FL), 3-O-fucosyllactose (3FL), lacto-N-tetraose (LNT), lacto-N-neotetraose (LNnT), 6′-O-sialyllactose (6′SL), 3′-O-sialyllactose (3′SL), together representing the three types of structures found in human breastmilk (fucosylated, sialylated and neutral core). On the basis of the HMO composition in breastmilk86,87, these were mixed in a ratio of 53% 2′FL, 18% 3FL, 13% LNT, 5% LNnT, 7% 6′SL and 4% 3′SL in sterile water to obtain a representative HMO mix used in the in vitro experiments at 2% (w/v).
Growth experiment with B. longum ssp. longum 105-A strains
B. longum ssp. longum 105-A (JCM 31944)30 was obtained from Japan Collection of Microorganisms (RIKEN BioResource Research Center). B. longum ssp. longum 105-A strains (WT, insertional mutant (type 4 ldh::pMSK127) and plasmid-complemented mutant (type 4 ldh::pMSK127/ pMSK128 (Pxfp-type 4 ldh)); Supplementary Table 6) were cultivated on MRSc or MRSc-Chl (MRSc supplemented with 2.5 μg ml–1 of chloramphenicol) agar plates for 48 h at 37 °C anaerobically. Single colonies were dissolved in 5.4 ml of MRSc or MRSc-Chl broth, tenfold serially diluted and incubated for 15 h at 37 °C anaerobically with shake. The most diluted culture (exponential phase) was washed in same medium (10,000g, room temperature, 5 min) and resuspended in MRSc or MRSc-Chl broth to yield OD600nm = 1 and subsequently diluted 1:40 in prewarmed and reduced MRSc or MRSc-Chl broth (in triplicates), before incubation at 37 °C, anaerobically with shake. The cultures were sampled (500 μl) every hour for OD600nm measurements and the culture supernatants (16,000g, 4°C, 5 min) from early (13 h) stationary phase were analysed by UPLC–MS for aromatic amino metabolites and by GC–MS for lactate.
Identification of aldh gene/operon in Bifidobacterium strains, metagenomic data and homology searches
From the full genome sequences (available at NCBI Genome, https://www.ncbi.nlm.nih.gov/genome/) of Bifidobacterium type strains included in this study (Supplementary Table 6) all genes annotated as LDHs were aligned (gap cost 10, gap extension cost 1) and subsequently a phylogenetic tree (Algorithm = Neighbor-Joining, Distance measure = Jukes-Cantor, 100 bootstrap replications) was constructed in CLC Main Workbench (v.7.6.3, CLCbio, Qiagen). The tree was visualized by use of the FigTree software v.1.4.3 (http://tree.bio.ed.ac.uk/software/figtree/). For identification of aldh (type 4 ldh) in Bifidobacterium strains, all complete human gut-associated Bifidobacterium genomes (n = 127) including plasmids were retrieved from NCBI Genome and aldh genes were identified using NCBI tBLASTn with default settings and a cutoff of 70% identity and 70% query coverage. Aligned genomic nucleotide sequences were translated and verified to match LDHs using reciprocal BLASTx against NCBI’s non-redundant database. In addition, the ALDH amino acid sequence (translated from the aldh nucleotide sequence) of B. longum ssp. longum 105-A was aligned (gap cost 10, gap extension cost 1) with the ALDH amino acid sequences of the B. longum ssp. longum, B. longum ssp. infantis, B. bifidum, B. breve and B. scardovii type strains and pairwise amino acid identity percentages were calculated in CLC Main Workbench. Potential non-bifidobacterial ALDH homologues were searched for by BLASTp analysis of the 105-A amino acid sequence against the non-redundant protein sequence database and the Swiss-Prot database using a cutoff of 60% amino acid sequence identity. Further, comparison of aldh gene cluster/operon in 12 Bifidobacterium type strains (Supplementary Fig. 4) was conducted by pairwise alignments in MBGD (Microbial Genome Database for Comparative Analysis; http://mbgd.genome.ad.jp/). The amino acid sequences of the gene cluster from B. pseudolongum ssp. pseudolongum type strain was collected from NCBI database (https://www.ncbi.nlm.nih.gov/genome/) and was used for comparison with that from B. animalis ssp. animalis type strain. Using 193 infant samples collected at 4 and 12 months of age with data on feeding practice available and data from 98 mothers5, we used IGGsearch and IGGdb v.1.0.0 (ref. 88) to identify Bifidobacterium MAGs. MAGs were included in the analysis if they passed the following criteria: --min-reads-gene=2 --min-perc-genes=40 --min-sp-quality=75. For each Bifidobacterium MAG identified, we used the representative genome to search for aldh genes. The aldh genes were identified using NCBI tBLASTn with default settings and a cutoff of 70% identity and 70% query coverage.
Recombinant expression of aldh (type 4 ldh)
Chemically competent cells for recombinant expression
E. coli LMG194 ON culture (200 μl) was inoculated into 5 ml of Luria-Bertani (LB) medium and incubated at 37 °C, 250 r.p.m. until OD600nm = 0.5, at which time the culture was centrifuged for 5 min at 10,000g at 4 °C and supernatant discarded. Cell pellet was resuspended in ice-cold 1.8 ml of 10 mM MgSO4 (Sigma, M2643) and centrifuged for 2 min at 5,000g at 0 °C. Supernatant was discarded and cell pellet resuspended in 1.8 ml of ice-cold 50 mM CaCl2 (Merck, 1.02083.0250), incubated on ice for 20 min and centrifuged for 2 min at 5,000g at 0 °C. Cell pellet was resuspended in 0.2 ml of ice-cold 100 mM CaCl2, 10 mM MgSO4 and placed on ice until transformation.
Cloning and recombinant expression
Genomic DNA was extracted (PowerLyzer PowerSoil DNA isolation kit, MoBio 12855-100) from colony material of B. longum ssp. infantis DSM 20088T (ATCC 15697). To amplify the type 4 ldh gene, 50 ng of template DNA was mixed with 5 μl of 10× PCR buffer, 0.5 μl (50 mM) of dNTP mix, 1 μl (10 μM) of forward primer (ldh4_F, 5′-ACCATGGTCACTATGAACCG-3′), 1 μl (10 μM) of reverse primer (ldh4_R, 5′-AATCACAGCAGCCCCTTG-3′) and 1 μl (1 U μl–1) of Platinum Taq DNA polymerase (Invitrogen, 10966-018) in a 50 μl total reaction volume. The PCR programme included 2 min at 94 °C, 35 cycles of 30 s at 94 °C, 30 s at 55 °C, 60 s at 72 °C, followed by a final extension 10 min at 72 °C. The PCR product was purified (MinElute PCR purification kit, Qiagen, 28004) and 4 μl was mixed with 1 μl of salt solution (1.2 M NaCl, 0.06 M MgCl2) and 1 μl of pBAD-TOPO plasmid (Invitrogen, K4300-01) and incubated for 5 min at room temperature. A total of 2 μl of the cloning mixture was transformed into 50 μl of One Shot TOP10 Competent Cells (Invitrogen, K4300-01) by gentle mix, incubation 15 min on ice and heat-shock for 30 s at 42 °C. A total of 250 μl of S.O.C medium (Invitrogen, K4300-01) was added and incubated at 37 °C for 1 h at 200 r.p.m. and subsequently spread on LB-AMP (LB supplemented with 20 μg ml–1 Ampicillin (Sigma, A9518)) agar plates and incubated at 37 °C ON. Transformants were picked and clean streaked on LB-AMP agar plates, incubated at 37 °C ON and afterwards single colonies of each transformant was inoculated into 5 ml of LB-AMP broth and incubated at 37 °C for 15 h at 250 r.p.m. Plasmid DNA was isolated (QIAprep Spin Miniprep Kit, Qiagen, 27104) from each transformant and subsequently 5 μl of plasmid DNA (80–100 ng μl–1) was mixed with 5 μl (5 pmol μl–1) pBAD forward (5′-ATGCCATAGCATTTTTATCC-3′) or reverse (5′-GATTTAATCTGTATCAGG-3′) sequencing primers (5 pmol μl–1) and shipped for sequencing at GATC (GATC-biotech). To remove the leader peptide in pBAD-TOPO, 10 μl of plasmid (0.1 μg) with correct insert was cut with FastDigest NcoI (Thermo Scientific, FD0563) for 10 min at 37 °C and the enzyme inactivated 15 min at 65 °C. Plasmid was ligated using 1 μl (1 U μl–1) T4 DNA Ligase (Invitrogen, 15224-017) for 5 min at room temperature and subsequently 2 μl of plasmid was transformed into 100 μl of chemically competent E. coli LMG194 cells by incubation on ice for 30 min, followed by heat-shock at 43 °C for 3 min and incubation on ice for 2 min. A total of 900 μl of LB medium was added and cells were incubated at 37 °C for 1 h at 250 r.p.m., before plating on LB-AMP agar plates and incubation at 37 °C ON. Transformants were picked, clean streaked and plasmid DNA isolated and sequenced as described above. A transformant with correct insert was selected for recombinant expression of the type 4 ldh gene; 2 ml of LB-AMP broth was inoculated with a single recombinant colony or the non-transformed E. coli LMG194 (negative control) and grown at 37 °C ON at 250 r.p.m. In 3× triplicates, 100 μl of the ON cultures (2×3× 100 μl of transformant culture + 1×3× 100 μl of non-transformed E. coli LMG194 culture) were diluted 100-fold into 9.9 ml of prewarmed LB-AMP/LB broth and grown at 37 °C, 250 r.p.m. until OD600nm ≈ 0.5, at which 9 ml of culture was added to 1 ml of mix of indolepyruvic acid, phenylpyruvic acid and 4-hydroxyphenylpyruvic acid (1 mg ml–1 each). The cultures were sampled (time zero) and subsequently 100 μl of 20% l-arabinose (or 100 μl of sterile water; control for induction) was added to induce gene expression and the cultures were re-incubated at 37 °C, 250 r.p.m., before sampling at 1 and 5 h post-induction for OD600nm measurements and assessment of production of aromatic lactic acids. For the latter, samples were centrifuged at 16,000g for 5 min at 4 °C and supernatants were stored at –20 °C for UPLC–MS analyses.
Construction of aldh (type 4 ldh) insertional mutant and aldh complemented strain
Transformation of B. longum ssp. longum 105-A
B. longum ssp. longum 105-A cells were grown to exponential phase at 37 °C in Gifu anaerobic liquid medium (Nissui Pharmaceutical, catalogue no. 05422), harvested by centrifugation and washed twice with ice-cold 1 mM ammonium citrate buffer containing 50 mM sucrose (pH 6.0). The cells were concentrated 200 times with the same buffer and used for electroporation with settings of 10 kV cm–1, 25 μF and 200 Ω. After recovery culturing in Gifu anaerobic liquid medium at 37 °C for 3 h, the cells were spread onto Gifu anaerobic agar containing antibiotics (30 μg ml–1 spectinomycin and/or 2.5 μg ml–1 chloramphenicol) for selection.
Insertional mutant construction and plasmid complementation
The type 4 ldh gene (BL105A_0985) of B. longum ssp. longum 105-A was disrupted by a plasmid-mediated single crossover event as described previously89. The plasmid used for disruption was constructed using the In-Fusion cloning kit (Clontech Laboratories, catalogue no. 639649). E. coli DH5α was used as a host for genetic manipulation. In brief, the internal region of the ldh gene (position 142–638 of the nucleotide sequence of BL105A_0985 (ref. 90; Supplementary Fig. 5) was amplified by PCR using a primer pair Pr-580/581 (Supplementary Table 7) and ligated with the BamHI-digested pBS423 fragment carrying pUC ori and a spectinomycin-resistance gene29. The resulting plasmid pMSK127 was introduced into B. longum ssp. longum 105-A by electroporation to be integrated into type 4 ldh locus by single crossover recombination (type 4 ldh::pMSK127). Type 4 ldh disruption was confirmed by genomic PCR with a primer pair (Pr-543/546) designed to anneal outside of the gene (Supplementary Fig. 5 and Supplementary Table 7). The amplified fragment was also sequenced to ensure the correct recombination event. Complementation plasmid pMSK128 was constructed by ligating PCR-amplified xfp (xylulose 5-phosphate/fructose 6-phosphate phosphoketolase) promoter region (Pxfp) and the type 4 ldh coding region with PstI- and SalI-digested pBFS38 (ref. 91) using the In-Fusion cloning kit, by which type 4 ldh was placed under the control of Pxfp. Primer pairs of Pr-598/Pr-599 and Pr-600/Pr-601 were used for amplifying Pxfp from pBFS48 (ref. 91) and the type 4 ldh gene from the B. longum ssp. longum 105-A genome, respectively (Supplementary Table 7). The resulting plasmid was electroporated into type 4 ldh::pMSK127 to give type 4 ldh::pMSK127/ pMSK128 (Pxfp-type4_ldh) (Supplementary Fig. 5).
Biochemical characterization of ALDH (type 4 LDH)
Recombinant expression and purification
Type 4 LDH (BL105A_0985) was recombinantly expressed as a non-tagged form. The gene was amplified by PCR using the genomic DNA of B. longum ssp. longum 105-A as a template and a primer pair of Pr-617 (5′-GGTGGTGGTGCTCGAGTCACAGCAGCCCCTCGCAG-3′) and Pr-635 (5′-AAGGAGATATACATATGGTCACTATGAACCGC-3′). Underlined bases indicate 15 bp for In-Fusion cloning (Clontech). The amplified DNA fragment was inserted into the NdeI and XhoI site of pET23b(+) (Novagen) using an In-Fusion HD cloning kit (Clontech). The resulting plasmid was introduced into E. coli BL21 (DE3) ΔlacZ carrying pRARE2 (ref. 89) and the transformant was cultured in LB medium supplemented with ampicillin (100 μg ml−1) and chloramphenicol (7.5 μg ml−1). When OD600nm reached 0.5, isopropyl β-d-thiogalactopyranoside was added at a final concentration of 0.02 mM to induce the protein expression. The culture was incubated for 4 d at 18 °C, harvested by centrifugation and resuspended in 50 mM potassium phosphate buffer (KPB; pH 7.0) supplemented with 1 mM 2-mercaptoethanol (2-ME) and 200 μM phenylmethane sulfonyl fluoride. Following cell disruption by sonication, the cleared lysate was saturated with ammonium sulfate (40–60%). The resulting precipitate was dissolved, dialysed against 20 mM KPB (pH 7.0) containing 1 mM 2-ME and concentrated by Amicon Ultra 10 K centrifugal device (Merck Millipore). The sample was then loaded onto an Affigel blue column (Bio-Rad) preequilibrated with 20 mM KPB (pH 7.0) containing 1 mM 2-ME and eluted by the same buffer containing 1 M NaCl. The protein was further purified by a Mono Q 5/50 (GE Healthcare; a linear gradient of 0–1 M NaCl in 20 mM Tris-HCl (pH 8.0) containing 1 mM 2-ME) and Superdex 200 Increase 10/300 GL column (GE Healthcare; 10 mM KPB (pH 7.0) containing 50 mM NaCl and 1 mM 2-ME). Protein concentration was determined by measuring the absorbance at 280 nm based on a theoretical extinction coefficient of 26,470 M–1 cm–1.
Enzyme assay
The standard reaction mixture contained 100 mM KPB (pH 8.0), 1 mM 2-ME, 0.1 mM β-NADH and the substrate. The reaction was initiated by adding the enzyme and the mixture was incubated at 37 °C for an appropriate time, in which the linearity of the reaction rate was observed. The substrate concentrations were varied between 0.01 and 0.25 mM for indolepyruvic acid 1.5 and 10.5 mM for phenylpyruvic acid, 2 and 24 mM for 4-hydroxyphenylpyruvic acid and 2.5 and 40 mM for pyruvic acid. The enzyme was used at the concentrations of 0.22 nM for indolepyruvic acid, 1.47 nM for phenylpyruvic acid, 0.12 nM for 4-hydroxyphenylpyruvic acid and 88.50 nM for pyruvic acid. The reducing reactions of phenylpyruvic acid and pyruvic acid were continuously monitored by measuring the decrease of the absorbance at 340 nm (NADH consumption). When 4-hydroxyphenylpyruvic acid and 4-indolepyruvic acid were used as the substrates, the reaction products 4-OH-PLA and ILA were quantified by HPLC after the termination of the reactions by adding 5% (w/v) trichloroacetic acid. HPLC analysis was performed using a Waters e2695 separation module (Waters) equipped with a LiChrospher 100 RP-18 column (250 × 4 mm, φ = 5 μm; Merck Millipore) at 50 °C. Following equilibration with a mixture of 10% solvent A (50% methanol, 0.05% trifluoroacetic acid) and 90% solvent B (0.05% trifluoroacetic acid) at a flow rate of 1 ml min–1, the concentration of solvent A was linearly increased to 100% for 25 min and maintained at 100% for additional 15 min. The 4-OH-PLA and ILA were detected by a Waters 2475 Fluorescence Detector with λex 277 nm and λem 301 nm and λex 282 nm and λem 349 nm, respectively. The standard curves were created using the known concentrations of both compounds. Experiments were performed at least in duplicate. Physicochemical property of the enzyme was examined by using 1 mM phenylpyruvic acid as a substrate. The effects of metal ions (0.1 mM each) on the enzyme activity was examined using 50 mM MES (2-(N-morpholino)ethanesulfonic acid) buffer (pH 7.0). EDTA was added at the final concentration of 0.1, 0.5 or 1 mM. The optimal pH was determined using 50 mM KPB (pH 6.0–8.5) and TAPS (N-Tris(hydroxymethyl)methyl-3-aminopropanesulfonic acid) buffer (pH 8.0–9.0). The thermostability was evaluated by the residual activities after incubating the enzyme (1.0 mg ml–1 in 10 mM KPB (pH 7.0) containing 50 mM NaCl and 1 mM 2-ME) at the indicated temperatures for 30 min before the assay. Fructose 1,6-bisphosphate, shikimate-3-phosphate, d-erythrose-4-phosphate and phosphoenolpyruvic acid were added to the reaction mixtures at the concentrations of 0.1 and 1 mM to examine their heterotropic effects. KPB, TAPS buffer or HEPES (4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) buffer (pH 8.0 each) containing 1 and 4 mM phenylpyruvic acid as a substrate were used. The effect of phosphate ion was analysed by adding various concentration of KPB (pH 8.0) into 10 mM HEPES buffer (pH 8.0). All experiments were conducted at least in duplicate. In the subsequent kinetic analysis, we used phosphate ion at the concentration of 100 mM because (1) no saturation was obtained for phosphate under the tested conditions (Supplementary Fig. 9a), (2) the intracellular phosphate concentration in Gram-positive bacteria is known to be 130 mM at maximum92 and (3) the strong homotrophic effect of the substrate phenylpyruvic acid was observed only in the presence of 10 mM phosphate ion.
In vivo monocolonization experiments
Germ-free (GF) Swiss Webster mice (Tac:SW, originally obtained from Taconic Biosciences) were bred and housed within GF isolators (Scanbur) in type II Makrolon cages (Techniplast) with bedding, nesting material, hiding place and a wooden block at the National Food Institute, Technical University of Denmark. The mice were fed an irradiated standard Altromin 1314 chow (Brogaarden) and the environment was maintained on a 12 h light/12 h dark cycle at a constant temperature of 22 ± 1 °C, with air humidity of 55 ± 5% relative humidity and change of air 50 times per hour. The GF condition of the mice before inoculation of bacteria was confirmed by plating of faecal sample suspensions on blood agar plates (Statens Serum Institut) incubated both aerobically and anaerobically. In two separate experiments, pregnant GF mice were randomized to be colonized with either B. longum 105-A WT (n = 4) or aldh (type 4 ldh) mutant (n = 5) by a single oral gavage (200 μl, ≈5 × 107 c.f.u. per dose) 1 week before giving birth. The monocolonized offspring (nwildtype = 21, 12 males and 9 females; naldhmutant = 29, 18 males and 11 females) were euthanized at 4 weeks of age by cervical dislocation and dissected to collect caecal contents. Successful colonization with B. longum and absence of contamination in monocolonized offspring was confirmed by cultivation of caecal content on MRSc and blood agar plates incubated both aerobically and anaerobically. Aromatic lactic acids were quantified from caecal content. All mouse experiments were approved by the Danish Animal Experiments Inspectorate (license no. 2015-15-0201-00553) and carried out in accordance with existing Danish guidelines for experimental animal welfare.
Metabolomics
Chemicals
Authentic standards of the aromatic amino acids and derivatives (Supplementary Table 1) were obtained from Sigma-Aldrich, whereas isotope-labelled aromatic amino acids used as internal standards (l-phenylalanine (ring-d5, 98%), l-tyrosine (ring-d4, 98%), l-tryptophan (indole-d5, 98%) and indoleacetic acid (2,2-d2, 96%)) of the highest purity grade available were obtained from Cambridge Isotope Laboratories.
Extraction of metabolites from faecal samples
Faecal samples (100–500 mg) from the SKOT (n = 59) and the CIG cohort (n = 267, data from two samples missing due to insufficient sample material (n = 1) and problems detecting the internal standards (n = 1)) were diluted 1:2 with sterile MQ water, vortexed for 10 s and centrifuged at 16,000g, 4 °C for 5 min. Subsequently, the supernatant liquor was transferred to a new tube and centrifuged again at 16,000g, 4 °C for 10 min. Finally, an aliquot of 150–300 µl was stored at –20 °C. All samples were later thawed at 4 °C, centrifuged at 16,000g, 4 °C for 5 min, and diluted in a total volume of 80 µl of water corresponding to a 1:5 dilution of the faecal sample. To each sample, 20 µl of internal standard mix (4 µg ml–1) and 240 µl of acetonitrile were added. The tubes were vortexed for 10 s and left at –20 °C for 10 min to precipitate the proteins. The tubes were then centrifuged at 16,000g, 4 °C for 10 min, and each supernatant (320 µl) was transferred to a new tube, which was dried with nitrogen gas. Subsequently, the residues were reconstituted in 80 µl of water (equalling a 1:5 dilution of the faecal sample with internal standards having a concentration of 1 µg ml–1), vortexed for 10 s, centrifuged at 16,000g, 4 °C for 5 min and transferred to an LC vial, which was stored at −20 °C until analysis.
Extraction of metabolites from urine samples
Urine samples (n = 49) from the SKOT cohort were thawed in a refrigerator and all procedures during the sample preparation were carried out at 0–4 °C using an ice bath. The subjects were randomized between analytical batches by placing all the samples from the each subject in the same 96-well plate. The run order of the samples was randomized within the analytical batch. Urine samples were centrifuged at 3,000g for 2 min at 4 °C. A total of 150 µl of each urine sample were added to separate wells and diluted with 150 µl of diluent (MQ water: formic acid (99.9:0.1, v/v) / internal standard mixture (100 µg ml–1) (90:10, v/v). A blank sample (diluent), standard mixture of external standard containing 44 biologically relevant metabolites (metabolomics standard)93 and pooled sample containing equal amounts of each sample (20 µl) were added to spare wells as quality control samples. The plates were stored at −80 °C until the analysis. Immediately before analysis, the plates were thawed and mixed by vortex stirring for 10 min.
Extraction of metabolites from in vitro fermentation samples
Supernatants from in vitro fermentations were thawed at 4 °C, centrifuged at 16,000g, 4 °C for 10 min, before 80 µl was transferred to a new tube. To each sample, 20 µl of internal standard (40 µg ml–1) and 300 µl of acetonitrile were added. The tubes were vortexed for 10 s and left at –20 °C for 10 min to precipitate the proteins. Following, the tubes were centrifuged at 16,000g, 4 °C for 10 min before 50 µl of each sample was diluted with 50 µl of sterile water and transferred to an LC vial (equalling a 1:10 dilution of the sample with internal standards having a concentration of 1 µg ml–1).
Metabolic profiling of faecal, caecal and in vitro samples using UPLC–MS
Aromatic amino acids and derivatives (Supplementary Table 1) of faecal and in vitro samples were quantified by a semiquantitative UPLC–MS method94. In brief, samples were analysed in random order. For the analysis of the CIG faecal samples, a pooled quality control (QC) sample was injected for every ten samples. In all cases, five standard mix solutions (0.1, 0.5, 1, 2 and 4 μg ml–1) were analysed once for every ten samples to obtain a standard curve for every ten samples. For each sample, a volume of 2 µl was injected into a ultraperformance liquid chromatography quadrupole time-of-flight mass spectrometry (UPLC-QTOF-MS) system consisting of Dionex Ultimate 3000 RS liquid chromatograph (Thermo Scientific) coupled to a Bruker maXis time-of-flight mass spectrometer equipped with an electrospray interphase (Bruker Daltonics) operating in positive mode. The analytes were separated on a Poroshell 120 SB-C18 column with a dimension of 2.1 × 100 mm and 2.7 μm particle size (Agilent Technologies) as previously published94. Aromatic amino acids and derivatives were detected by selected ions and semiquantified by isotopic internal standards with similar molecular structures as listed in Supplementary Table 1. The recoveries of the internal standards varied but were, relative to each other, in general rather consistent (Supplementary Fig. 14) emphasizing that while the absolute concentrations may not be accurate due to lack of isotope-labelled internal standards for each single analyte, the relative metabolite concentrations across samples were robust with the applied LC-MS method. Data were processed using QuantAnalysis v.2.2 (Bruker Daltonics) and bracket calibration curves for every ten lumen samples were obtained for each metabolite. The calibration curves were established by plotting the peak area ratios of all of the analytes with respect to the internal standard against the concentrations of the calibration standards. The calibration curves were fitted to a quadratic regression.
For untargeted metabolomics, the raw UPLC–MS data, obtained by analysis of the CIG faecal samples in positive ionization mode, were converted to mzXML files using Bruker Compass DataAnalysis 4.2 software (Bruker Daltonics) and preprocessed as previously reported95 using the R packpage XCMS (v.1.38.0; ref. 96). Noise filtering settings included that features should be detected in minimum 50% of the samples. A data table was generated comprising mass-to-charge (m/z), retention time and intensity (peak area) for each feature in the every sample. The data were normalized to the total intensity and log-transformed. Subsequently, features with a coefficient of variation >0.3 in the QC samples and features with a retention time <0.5 min were excluded from the data. Parent ion masses of HMO compounds of interest (2′FL/3FL, LNT/LNnT, 3′SL/6′SL) were searched in the cleaned dataset with 0.02 Da m/z and 0.02 min retention time tolerance. Subsequently, the identities of the features of interest were confirmed at level 1 (ref. 97) by tandem mass spectrometry and comparison to authentic standards (Supplementary Table 8). Of notice, HMO isomers could not be distinguished with the method applied due to identical retention times.
Metabolic profiling of urine samples using UPLC–MS
The samples were analysed by UPLC-QTOF-MS equipped with an electrospray ionization (ESI) (Waters Corporation). Reverse phase HSS T3 C18 column (2.1 × 100 mm, 1.8 µm) coupled with a precolumn (VanGuard HSS T3 C18 column (2.1 × 5 mm, 1.8 µm)) were used for chromatographic separation. A total of 5 µl of each well was injected into the mobile phase A (0.1% formic acid in MQ water), mobile phase B (10% 1 M ammonium acetate in methanol), mobile phase C (methanol) and mobile phase D (isopropanol). Mobile phase gradient during the run time of 10 min was as follows: start condition (100% A), 0.75 min (100% A), 6 min (100% C), 6.5 min (70% B, 30% D), 8 min (70% B, 30% D), 8.1 min (70% B, 30% D), 9 min (100% A) and 10 min (100% A). The flow rate gradient was as follows: start condition (0.4 ml min–1), 0.75 min (0.4 ml min–1), 6 min (0.5 ml min–1), 6.5 min (0.5 ml min–1), 8 min (0.6 ml min–1), 8.10 min (0.4 ml min–1), 9 min (0.4 ml min–1) and 10 min (0.4 ml min–1). ESI was operated in negative mode with 3.0 kV capillary probe voltage. The cone voltage and the collision energy were set at 30 kV and 5 eV, respectively. Ion source and desolvation gas (nitrogen) temperature were 120 and 400 °C while sampling cone and desolvation gas flow rates were 50 and 1,000 l h–1. Scan time set as 0.08 s with 0.02 s interscan time for both modes. Data were acquired in centroid mode with mass range between 50 and 1,500 Da. Leucine-enkephalin (500 ng ml–1) was infused as the lock-spray agent to calibrate the mass accuracy every 5 s with 1 s scan time. Quality control samples were used to evaluate possible contamination, monitoring the changes in mass accuracy, retention time and instrumental sensitivity drifts93,98.
The raw data were converted to netCDF format using DataBridge Software v.3.5 (Waters) and imported into MZmine v.2.28 (ref. 99). A subset of samples was used to optimize the preprocessing parameters for the positive and negative mode data separately. Optimized preprocessing parameters are listed in Supplementary Table 9. Data preprocessing was used with the following steps: mass detection, chromatogram builder, chromatogram deconvolution, deisotoping, peak alignment and gap filling. After the preprocessing, each detected peak was represented by a feature defined with a retention time, m/z and peak area.
The data matrix was imported into MATLAB R2015b (MathWorks). Features that were present in the blanks, were very early and late eluting (retention time < 0.30 and retention time > 9.46 min), potential isotopes, duplicates as well as features with masses indicating multiple charges were removed from the dataset using an in-house algorithm. The data were normalized using unit length normalization to correct the variation in urine concentration. Parent ion masses of the aromatic lactic acids (ILA, PLA and 4-OH-PLA) were searched in the cleaned dataset with 0.02 Da m/z and 0.02 s retention time tolerance. A linear regression model was used feature-wise to correct for batch differences and instrumental sensitivity drifts100. The aromatic lactic acids were confirmed at level 1 (ref. 97) by comparison to authentic standards and by tandem mass spectrometry using the same experimental conditions (Supplementary Figs. 1–3).
Lactic acid production by B. longum ssp. longum 105-A strains using GC–MS
The lactic acid production of the B. longum ssp. longum 105-A WT, type 4 ldh mutant and type 4 ldh complemented strains were assessed in supernatants obtained after 13 h of growth (early stationary phase) by gas chromatography–mass spectrometry (GC–MS) on methyl chloroformate derivatization using a slightly modified version of the protocol previously described101. All samples were analysed in a randomized order. Analysis was performed using GC (7890B, Agilent Technologies) coupled with a quadrupole detector (59977B, Agilent Technologies). The system was controlled by ChemStation (Agilent Technologies). Raw data was converted to netCDF format using Chemstation, before the data was imported and processed in Matlab R2014b (Mathworks) using the PARADISe software102.
Rat AhR reporter gene assay
Rat hepatoma cells (H4IIE) stably transfected with a luciferase reporter gene under the control of AhR (pGudLuc1.1) were used. The cells were kindly provided by M. S. Denison (University of California, Davis). The assay was conducted as previously described103, where cells were incubated for ∼22 h in Minimum Essential Medium (MEM) α with 1% fetal bovine serum (FBS) and 1% penicillin/streptomycin/fungizone. Chemical exposure was performed for 24 h and successively luminescence was measured. Cell viability was analysed by measuring ATP levels with the CellTiter-Glo Luminescent Assay according to the manufacturer’s instruction (Promega). As a positive control, 2,3,7,8-tetrachlorodibenzo-p-dioxin was used. Three experiments in triplicates were conducted with five twofold dilutions of ILA and IAld ranging from 12.5 to 200 µM with a constant vehicle concentration in all wells. Further, sterile-filtered faecal water (10 mg faeces per ml MQ water) obtained from all samples (n = 119) of 11 selected CIG infants (Fig. 4d–f) were run in technical triplicates in the assay. Only mild toxicity that did not correlate with AhR-induced luminescence signal was observed for some faecal water samples.
Human AhR reporter gene assay
ILA and IAld (positive control)15 were tested for activation of the human AhR. AhR Reporter Cells from Indigo Biosciences (catalogue no. IB06001) that include a luciferase reporter gene functionally linked to an AhR-responsive promoter were used. The assay was run according to the instructions of the manufacturer (technical manual v.6.0) with the reference agonist MeBIO as the positive control. Three experiments in triplicates were conducted with five twofold dilutions of ILA and IAld ranging from 12.5 to 200 µM with a constant vehicle concentration in all wells. No cytotoxicity was observed for any of the tests as determined by a resazurin toxicity assay.
Human HCA3 receptor assay
The aromatic lactic acids (ILA, PLA and 4-OH-PLA) were tested for activation of the HCA3 receptor, which is a Gαi-coupled receptor (GPCR). The cAMP Hunter eXpress GPR109B CHO-K1 GPCR Assay for chemiluminescence detection of cAMP was used (DiscoveRx Corporation, catalogue no. 95-0141E2CP2M). Following ligand stimulation of cells overexpressing the HCA3 receptor, the functional status of the receptor was monitored by measuring cellular cAMP levels using a homogeneous, competitive immunoassay based on Enzyme Fragment Complementation technology. The assay was run in agonist mode in a 96-well plate format according to the instructions of the manufacturer (DiscoveRx Corporation) in the presence of 15 µM forskolin. Eleven threefold dilutions of ILA ranging from 0.03 to 1,574 µM and of PLA and OH-PLA ranging from 0.02 to 1,000 µM were tested twice in duplicates.
Ex vivo stimulation of human immune cells
Human buffy coats were acquired from the Copenhagen University Hospital (Rigshospitalet) from healthy anonymous donors. Use of the buffy coat material from healthy anonymous donors was approved by the Blood bank at Rigshospitalet, Copenhagen, under the jurisdiction of Region H. Prior written informed consent was obtained according to the Declaration of Helsinki. Blood samples were handled in accordance with guidelines put forward in the ‘Transfusion Medicine Standards’ by the Danish Society for Clinical Immunology (www.dski.dk).
Isolation, cell culture and stimulation of T cells
Peripheral blood mononuclear cells (PBMCs) were isolated from whole blood by density centrifugation on Lymphoprep and cryopreserved at −150 °C in FBS with 10% DMSO until the day of cell culture. For cultivation, PBMCs were thawed and CD4+ T cells isolated using EasySep Human CD4+ T Cell Isolation Kit (Stemcell, 17952) following the manufacturers protocol. In short, ~2.5 × 107 PBMCs were incubated for 5 min at room temperature in 500 µl of IMDM-medium containing 50 µl of CD4+ T cell isolation cocktail, followed by the addition of 50 µl of RapidSpheres. Subsequently, the volume was topped up to 2.5 ml with IMDM-medium, the cells placed in an EasySep magnet (Stemcell) and incubated at room temperature for 3 min. The pure CD4+ T cell fraction was obtained by pouring the enriched non-bound cell fraction into a new tube. Enriched CD4+ T cells were cultured in TH17-polarizing culture medium (IMDM supplemented with 10% FCS, 20 mM HEPES (pH 7.4), 50 µM 2-mercaptoethanol, 2 mM l-glutamine and penicillin–streptomycin (10,000 U ml–1), 30 ng ml–1 IL-6, 10 ng ml–1 IL-1β, 0.5 ng ml–1 TGFβ-1, 10 ng ml–1 IL-23, 25 µl ml–1 ImmunoCult Human CD3/CD28 T cell activator for 3 d at 37 °C and 5% CO2 in Falcon polystyrene 48-well plates (Thermo Fisher, 10059110). Each culture condition contained 0.2% DMSO with or without the indicated amounts of ILA and/or the AhR-inhibitor CH-223191. After 3 d of culture, supernatants were collected for ELISA and frozen down until further use. The ELISA to detect IL-22 was performed in technical duplicates using the ELISA MAX Deluxe Set Human IL-22 kit (Biolegend, 434504) following the supplied manufacturer’s protocol. In short, a Nunc MaxiSorb flat-bottom 96-well plate (Thermo Fischer, 44-2404-21) was coated for 12 h at 4 °C with IL-22 coating antibody followed by four rounds of washing with PBS + 0.05% Tween-20. The washed plate was blocked with supplied assay diluent A buffer for 1 h at room temperature and 400 r.p.m., washed four more times with PBS + 0.05% Tween-20 and incubated with cell culture supernatants for 2 h at room temperature and 400 r.p.m. Serially diluted standard controls and a blank control were included as a reference. To detect bound IL-22, the plate was washed four times with PBS + 0.05% Tween-20 and incubated with IL-22 detection antibody for 1 h at room temperature and 400 r.p.m. After four further washing steps with PBS + 0.05% Tween-20, Avidin-HRP was added for 30 min at room temperature and 400 r.p.m. To detect HRP activity, the plate was washed five times with PBS + 0.05% Tween-20 followed by an incubation with Solution F substrate solution in the dark at room temperature. HRP activity was stopped after 20 min using 1 M H2SO4 and the optical density recorded (absorption at 450 nm) using a PowerWave HT Microplate Spectrophotometer (BioTek Instruments). Values below limit of detection (16 pg ml–1) of the kit were set to LOD/2. Sources and identifiers of all reagents used are given in Supplementary Table 10.
Isolation, cell culture and stimulation of monocytes
PBMCs were isolated by Ficoll-Paque (GE Healthcare) density centrifugation. Monocytes were isolated to >92% purity using the Pan Monocyte Isolation kit (Miltenyi Biotec). Cells were stained with diluted (1:25) CD14-PE-Cy7 (eBiosciences) and diluted (1:25) CD16-FITC (Biolegend) to determine the purity of monocytes (Supplementary Fig. 15) by flow cytometry (BD FACS Canto II). Monocytes were cultured in culture medium (RPMI 1640 (Lonza) containing 2 mM l-glutamine (Lonza), 10% heat-inactivated fetal bovine serum (Lonza), 1% penicillin/streptomycin (Lonza), 50 µM 2-ME (Sigma-Aldrich)) in a humidified 37 °C, 5% CO2 incubator. ILA (Sigma I5508), dissolved in a maximum of 0.1% DMSO, was added to the cells, with the final concentrations of ILA 5, 50 or 200 μM, respectively. Vehicle (0.1% DMSO) was added as a control. LPS (TLR4 ligand, E. coli O26:B6, Sigma L2654) at 100 ng ml–1 (final concentration) and IFN-γ (RD285-IF-100) at 10 ng ml–1 (final concentration) were then added to the cells and they were stimulated for 18 h. To determine the contribution of AhR, monocytes were pretreated for 1 h before addition of above compounds with the AhR antagonist CH-223191 (Sigma C8124) at 10 μM. For HCA3 silencing, where no specific antagonist is available, we performed reverse transfection using Lipofectamine RNAiMAX in Opti-MEM (Life Technologies) added 10 nM of scrambled siRNA (ScrRNA) (Thermo Fisher 4390846) or HCA3-specific siRNA (Thermo Fisher 4427037), reaching a maximum knockdown of 79% on the basis of qPCR validation (HCA3 primer (Thermo Fisher 4448892) versus GAPDH) as compared to ScrRNA targeted cells. Monocytes were pretreated with the siRNA constructs for 24 h before stimulation as above. Viability of cells was >97% as analysed by flow cytometry (BD FACS Canto II) using SYTOX AAD. Supernatants of stimulated cells were harvested after 18 h and kept at –80 °C until analysis. IL-12p70 was quantified using Meso Scale Discovery kits as previously detailed104.
Statistics and reproducibility
All experiments were performed with full factorial (biological and technical) replication. Data collection and analysis were not performed blind to the conditions of the experiments. No data were excluded from the analyses, except in the CIG cohort, six samples were omitted from the PCoA due to low read counts (<8,000), for correlation analyses between Bifidobacterium species and HMO residuals in faeces, 12 samples with no reported breastfeeding were excluded. One donor was excluded from the 200 µM indolelactate (ILA) stimulation of monocytes, since something went wrong during the stimulation. Statistical analyses were performed using QIIME v.1.9 (ref. 79), R v.3.1 (ref. 105) and GraphPad Prism v.8.1 (GraphPad Software). If data were normally distributed (evaluated by visual inspection and D’Agostino–Pearson test), parametric statistical tests were used, whereas non-parametric tests were used with non-normally distributed data. OTU distance/dissimilarity matrices were generated from OTU tables with rarefied read counts (2,000 sequences per sample for SKOT and 8,000 sequences per sample for CIG) and ordination (PCoA, beta_diversity.py script, default settings) and statistical tests (ADONIS and PERMDISP tests, compare_categories.py script, permutations = 999, default settings) of OTU distance/dissimilarity matrices were performed in QIIME. PCoA plots were illustrated (ggplot function) in R using the ggplot2 (v.3.3.3) package106. PCA of metabolite concentrations was performed in R using the prcomp function with zero-centring and autoscaling and illustrated using the ggbiplot function107 within ggplot2. Spearman’s rank correlations were performed in GraphPad Prism, whereas repeated measures correlation analyses and linear mixed models were performed in R using the packages rmcorr (v.0.4.3)32 (rmcorr function with default settings) and maaslin2 (v.1.0.0)31, respectively. In MaAsLin2 linear mixed models (Maaslin2 function) subject and age were included as random effects and the individual faecal aromatic lactic acids or HMOs as fixed effects, with default settings, except that total sum scaling normalization was not performed when including absolute abundances of bacterial taxa. The faecal HMO data were log10-transformed. Heatmaps and hierarchical clustering of correlation coefficient were generated in R using the heatmap.2 function (default settings) within the gplots (v.3.1.1) package108 and visualized in GraphPad Prism. Longitudinal metabolite and taxonomic abundance were modelled using LOESS regression and associations between taxa, metabolites and in vitro AhR activity (log10-transformed) were modelled using linear regression in R using the ggplot function (method = ’loess’ or ‘lm’) within the ggplot2 package106. For enzyme kinetics, the parameters (kcat, K0.5 and Hill coefficient nH) were calculated by curve-fitting the experimental data to the Hill equation, using GraphPad Prism. Two-tailed paired or unpaired Student’s t-test or two-tailed non-parametric Mann–Whitney U-test were performed when comparing two groups. For comparison of more than two groups, statistical significance was evaluated by one-way analysis of variance (ANOVA) or the non-parametric Kruskal–Wallis test. P <0.05 were considered statistically significant. When applicable, P values were corrected for multiple testing by the Benjamini–Hochberg false discovery rate (FDR)109 using a cutoff of 0.1.
Reporting Summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
All 16S rRNA gene amplicon sequencing data were deposited in the Sequence Read Archive (SRA) under the BioProjects PRJNA273694 (SKOT) and PRJNA554596 (CIG). The following databases were used: GreenGenes 16S rRNA database (https://greengenes.secondgenome.com/), RDP database (https://rdp.cme.msu.edu/), 16S rRNA gene sequence database at NCBI (https://ncbiinsights.ncbi.nlm.nih.gov/2020/02/21/rrna-databases/), non-redundant protein sequence database at NCBI (https://www.ncbi.nlm.nih.gov/protein/), Swiss-Prot database (https://www.uniprot.org/), MBGD (Microbial Genome Database for Comparative Analysis; http://mbgd.genome.ad.jp/) and the genome database at NCBI (https://www.ncbi.nlm.nih.gov/genome/). Metabolomics data (concentrations of aromatic amino acid metabolites) from SKOT and CIG cohorts are available in Supplementary Data 1e and 2d. Source data are provided with this paper.
Code availability
No custom code was used in the analyses. R scripts are available on request.
References
Victora, C. G. et al. Breastfeeding in the 21st century: epidemiology, mechanisms, and lifelong effect. Lancet 387, 475–490 (2016).
Horta, B. L., Loret de Mola, C. & Victora, C. G. Long-term consequences of breastfeeding on cholesterol, obesity, systolic blood pressure and type 2 diabetes: a systematic review and meta-analysis. Acta Paediatr. 104, 30–37 (2015).
Oddy, W. H. Breastfeeding, childhood asthma, and allergic disease. Ann. Nutr. Metab. 70, 26–36 (2017).
Laursen, M. F. et al. Infant gut microbiota development is driven by transition to family foods independent of maternal obesity. Msphere https://doi.org/10.1128/mSphere.00069-15 (2016).
Bäckhed, F. et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 17, 690–703 (2015).
Stewart, C. J. et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 562, 583–588 (2018).
Bode, L. The functional biology of human milk oligosaccharides. Early Hum. Dev. 91, 619–622 (2015).
Gensollen, T., Iyer, S. S., Kasper, D. L. & Blumberg, R. S. How colonization by microbiota in early life shapes the immune system. Science 352, 539–544 (2016).
Stokholm, J. et al. Maturation of the gut microbiome and risk of asthma in childhood. Nat. Commun. 9, 141 (2018).
Fujimura, K. E. et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 22, 1187–1191 (2016).
Khosravi, A. & Mazmanian, S. K. Disruption of the gut microbiome as a risk factor for microbial infections. Curr. Opin. Microbiol. 16, 221–227 (2013).
Fukuda, S. et al. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 469, 543–547 (2011).
Vatanen, T. et al. The human gut microbiome in early-onset type 1 diabetes from the TEDDY study. Nature 562, 589–594 (2018).
Roager, H. M. & Licht, T. R. Microbial tryptophan catabolites in health and disease. Nat. Commun. 9, 3294 (2018).
Zelante, T. et al. Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 39, 372–385 (2013).
Cervantes-Barragan, L. et al. Lactobacillus reuteri induces gut intraepithelial CD4+CD8αα+ T cells. Science https://doi.org/10.1126/science.aah5825 (2017).
Natividad, J. M. et al. Impaired aryl hydrocarbon receptor ligand production by the gut microbiota is a key factor in metabolic syndrome. Cell Metab. https://doi.org/10.1016/j.cmet.2018.07.001 (2018).
Dodd, D. et al. A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites. Nature 551, 648–652 (2017).
Guo, X. et al. Innate lymphoid cells control early colonization resistance against intestinal pathogens through ID2-dependent regulation of the microbiota. Immunity 42, 731–743 (2015).
Hoyles, L. et al. Molecular phenomics and metagenomics of hepatic steatosis in non-diabetic obese women. Nat. Med. 24, 1070–1080 (2018).
Krishnan, S. et al. Gut microbiota-derived tryptophan metabolites modulate inflammatory response in hepatocytes and macrophages. Cell Rep. 23, 1099–1111 (2018).
Madsen, A. L., Schack-Nielsen, L., Larnkjaer, A., Mølgaard, C. & Michaelsen, K. F. Determinants of blood glucose and insulin in healthy 9-month-old term Danish infants; the SKOT cohort. Diabet. Med. 27, 1350–1357 (2010).
Kato, K. et al. Age-related changes in the composition of gut bifidobacterium species. Curr. Microbiol. 74, 987–995 (2017).
Koga, Y. et al. Age-associated effect of kestose on Faecalibacterium prausnitzii and symptoms in the atopic dermatitis infants. Pediatr. Res. 80, 844–851 (2016).
Li, X., Jiang, B., Pan, B., Mu, W. & Zhang, T. Purification and partial characterization of Lactobacillus species SK007 lactate dehydrogenase (LDH) catalyzing phenylpyruvic acid (PPA) conversion into phenyllactic acid (PLA). J. Agric. Food Chem. 56, 2392–2399 (2008).
Koide, S., Iwata, S., Matsuzawa, H. & Ohta, T. Crystallization of allosteric l-lactate dehydrogenase from Thermus caldophilus and preliminary crystallographic data. J. Biochem. 109, 6–7 (1991).
Takashi, M., So, I., Hiroshi, S., Haruhiko, M. & Takahisa, O. Sequence and characteristics of the Bifidobacterium longum gene encoding l-lactate dehydrogenase and the primary structure of the enzyme: a new feature of the allosteric site. Gene 85, 161–168 (1989).
Bottacini, F. et al. Global transcriptional landscape and promoter mapping of the gut commensal Bifidobacterium breve UCC2003. BMC Genomics 18, 991 (2017).
Hirayama, Y. et al. Development of a double-crossover markerless gene deletion system in Bifidobacterium longum: functional analysis of the α-galactosidase gene for raffinose assimilation. Appl. Environ. Microbiol. 78, 4984–4994 (2012).
Matsumura, H., Takeuchi, A. & Kano, Y. Construction of Escherichia coli–Bifidobacterium longum shuttle vector transforming B. longum 105-A and 108-A. Biosci. Biotechnol. Biochem. 61, 1211–1212 (1997).
Mallick, H. et al. Multivariable association discovery in population-scale meta-omics studies 3. Preprint at bioRxiv https://doi.org/10.1101/2021.01.20.427420 (2021).
Bakdash, J. Z. & Marusich, L. R. Repeated measures correlation. Front. Psychol. 8, 456 (2017).
Alfa, M. J. et al. An outbreak of necrotizing enterocolitis associated with a novel Clostridium species in a neonatal intensive care unit. Clin. Infect. Dis. 35, S101–S105 (2002).
Butel, M.-J. et al. Conditions of bifidobacterial colonization in preterm infants: a prospective analysis. J. Pediatr. Gastroenterol. Nutr. 44, 577–582 (2007).
Moles, L. et al. Bacterial diversity in meconium of preterm neonates and evolution of their fecal microbiota during the first month of life. PLoS ONE 8, e66986 (2013).
Arboleya, S. et al. Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol. Ecol. 79, 763–772 (2012).
Korpela, K. et al. Intestinal microbiota development and gestational age in preterm neonates. Sci. Rep. 8, 2453 (2018).
Peters, A. et al. Metabolites of lactic acid bacteria present in fermented foods are highly potent agonists of human hydroxycarboxylic acid receptor 3. PLoS Genet. 15, e1008145 (2019).
Ahmed, K. et al. Deorphanization of GPR109B as a receptor for the β-oxidation intermediate 3-OH-octanoic acid and its role in the regulation of lipolysis. J. Biol. Chem. 284, 21928–21933 (2009).
Irukayama-Tomobe, Y. et al. Aromatic d-amino acids act as chemoattractant factors for human leukocytes through a G protein-coupled receptor, GPR109B. Proc. Natl Acad. Sci. USA 106, 3930–3934 (2009).
Hubbard, T. D. et al. Adaptation of the human aryl hydrocarbon receptor to sense microbiota-derived indoles. Sci. Rep. 5, 12689 (2015).
Quintana, F. J. et al. Control of Treg and TH17 cell differentiation by the aryl hydrocarbon receptor. Nature 453, 65–71 (2008).
Veldhoen, M., Hirota, K., Christensen, J., O’Garra, A. & Stockinger, B. Natural agonists for aryl hydrocarbon receptor in culture medium are essential for optimal differentiation of Th17 T cells. J. Exp. Med. 206, 43–49 (2009).
Veldhoen, M. et al. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 453, 106–109 (2008).
Komura, K., Hayashi, S. I., Makino, I., Poellinger, L. & Tanaka, H. Aryl hydrocarbon receptor/dioxin receptor in human monocytes and macrophages. Mol. Cell. Biochem. 226, 107–117 (2001).
Offermanns, S. Hydroxy-carboxylic acid receptor actions in metabolism. Trends Endocrinol. Metab. 28, 227–236 (2017).
Tamburini, S., Shen, N., Wu, H. C. & Clemente, J. C. The microbiome in early life: implications for health outcomes. Nat. Med. 22, 713–722 (2016).
Dominguez-Bello, M. G., Godoy-Vitorino, F., Knight, R. & Blaser, M. J. Role of the microbiome in human development. Gut 68, 1108–1114 (2019).
Yamada, C. et al. Molecular insight into evolution of symbiosis between breast-fed infants and a member of the human gut microbiome Bifidobacterium longum. Cell Chem. Biol. 24, 515–524 (2017).
Katayama, T. Host-derived glycans serve as selected nutrients for the gut microbe: human milk oligosaccharides and bifidobacteria. Biosci. Biotechnol. Biochem. 80, 621–632 (2016).
Thomson, P., Medina, D. A. & Garrido, D. Human milk oligosaccharides and infant gut bifidobacteria: molecular strategies for their utilization. Food Microbiol 75, 37–46 (2018).
Sakanaka, M. et al. Evolutionary adaptation in fucosyllactose uptake systems supports bifidobacteria–infant symbiosis. Sci. Adv. 5, eaaw7696 (2019).
Sakurai, T., Odamaki, T. & Xiao, J.-Z. Production of indole-3-lactic acid by bifidobacterium strains isolated from human infants. Microorganisms 7, 340 (2019).
Meng, D. et al. Indole-3-lactic acid, a metabolite of tryptophan, secreted by Bifidobacterium longum subspecies infantis is anti-inflammatory in the immature intestine. Pediatr. Res. 88, 209–217 (2020).
Ehrlich, A. M. et al. Indole-3-lactic acid associated with Bifidobacterium-dominated microbiota significantly decreases inflammation in intestinal epithelial cells. BMC Microbiol. 20, 357 (2020).
Neil, J. A. et al. IFN-I and IL-22 mediate protective effects of intestinal viral infection. Nat. Microbiol. 4, 1737–1749 (2019).
Turner, J.-E., Stockinger, B. & Helmby, H. IL-22 mediates goblet cell hyperplasia and worm expulsion in intestinal helminth infection. PLoS Pathog. 9, e1003698 (2013).
Zheng, Y. et al. Interleukin-22 mediates early host defense against attaching and effacing bacterial pathogens. Nat. Med. 14, 282–289 (2008).
Sugimoto, K. et al. IL-22 ameliorates intestinal inflammation in a mouse model of ulcerative colitis. J. Clin. Invest. 118, 534–544 (2008).
Keir, M. E., Yi, T., Lu, T. T. & Ghilardi, N. The role of IL-22 in intestinal health and disease. J. Exp. Med. 217, e20192195 (2020).
Natividad, J. M. et al. Impaired aryl hydrocarbon receptor ligand production by the gut microbiota is a key factor in metabolic syndrome. Cell Metab. 28, 737–749 (2018).
Metidji, A. et al. The environmental sensor AHR protects from inflammatory damage by maintaining intestinal stem cell homeostasis and barrier integrity. Immunity 49, 353–362 (2018).
Gomez de Aguero, M. et al. The maternal microbiota drives early postnatal innate immune development. Science 351, 1296–1302 (2016).
Kiss, E. A. et al. Natural aryl hydrocarbon receptor ligands control organogenesis of intestinal lymphoid follicles. Science 334, 1561–1565 (2011).
Qiu, J. et al. The aryl hydrocarbon receptor regulates gut immunity through modulation of innate lymphoid cells. Immunity 36, 92–104 (2012).
Lamas, B. et al. CARD9 impacts colitis by altering gut microbiota metabolism of tryptophan into aryl hydrocarbon receptor ligands. Nat. Med. 22, 598–605 (2016).
Dant, T. A. et al. T-cell expression of AhR inhibits the maintenance of pTreg cells in the gastrointestinal tract in acute GVHD. Blood 130, 348–359 (2017).
Wilck, N. et al. Salt-responsive gut commensal modulates TH17 axis and disease. Nature 551, 585–589 (2017).
Quintana, F. J. et al. Control of Treg and TH17 cell differentiation by the aryl hydrocarbon receptor. Nature 453, 65–71 (2008).
Ning, Y. et al. Antibacterial activity of phenyllactic acid against Listeria monocytogenes and Escherichia coli by dual mechanisms. Food Chem. 228, 533–540 (2017).
Rodríguez, N., Salgado, J. M., Cortés, S. & Domínguez, J. M. Antimicrobial activity of d-3-phenyllactic acid produced by fed-batch process against Salmonella enterica. Food Control 25, 274–284 (2012).
Narayanan, T. K. & Rao, G. R. Beta-indoleethanol and beta-indolelactic acid production by Candida species: their antibacterial and autoantibiotic action. Antimicrob. Agents Chemother. 9, 375–380 (1976).
Honoré, A. H. et al. Metabolic footprinting for investigation of antifungal properties of Lactobacillus paracasei. Anal. Bioanal. Chem. 408, 83–96 (2016).
Madsen, A. L., Larnkjær, A., Mølgaard, C. & Michaelsen, K. F. IGF-I and IGFBP-3 in healthy 9 month old infants from the SKOT cohort: breastfeeding, diet, and later obesity. Growth Horm. IGF Res. 21, 199–204 (2011).
Laursen, M. F. et al. Having older siblings is associated with gut microbiota development during early childhood. BMC Microbiol. 15, 154 (2015).
Laursen, M. F., Dalgaard, M. D. & Bahl, M. I. Genomic GC-content affects the accuracy of 16S rRNA gene sequencing based microbial profiling due to PCR bias. Front. Microbiol. https://doi.org/10.3389/fmicb.2017.01934 (2017).
Laursen, M. F. et al. Administration of two probiotic strains during early childhood does not affect the endogenous gut microbiota composition despite probiotic proliferation. BMC Microbiol. 17, 175 (2017).
Edgar, R. C. UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996–998 (2013).
Caporaso, J. G. et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336 (2010).
Wang, Q., Garrity, G. M., Tiedje, J. M. & Cole, J. R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 73, 5261–5267 (2007).
DeSantis, T. Z. et al. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 72, 5069–5072 (2006).
Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J. Basic local alignment search tool. J. Mol. Biol. 215, 403–410 (1990).
Callahan, B. J. et al. DADA2: high-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583 (2016).
Frese, S. A. et al. Persistence of supplemented Bifidobacterium longum subsp. infantis EVC001 in breastfed infants. Msphere https://doi.org/10.1128/mSphere.00501-17 (2017).
Lawley, B. et al. Differentiation of Bifidobacterium longum subspecies longum and infantis by quantitative PCR using functional gene targets. PeerJ 5, e3375 (2017).
Sprenger, N., Lee, L. Y., De Castro, C. A., Steenhout, P. & Thakkar, S. K. Longitudinal change of selected human milk oligosaccharides and association to infants’ growth, an observatory, single center, longitudinal cohort study. PLoS ONE 12, e0171814 (2017).
Xu, G. et al. Absolute quantitation of human milk oligosaccharides reveals phenotypic variations during lactation. J. Nutr. 147, 117–124 (2017).
Nayfach, S., Shi, Z. J., Seshadri, R., Pollard, K. S. & Kyrpides, N. C. New insights from uncultivated genomes of the global human gut microbiome. Nature 568, 505–510 (2019).
Sakurama, H. et al. Lacto-N-biosidase encoded by a novel gene of Bifidobacterium longum subspecies longum shows unique substrate specificity and requires a designated chaperone for its active expression. J. Biol. Chem. 288, 25194–25206 (2013).
Kanesaki, Y. et al. Complete genome sequence of Bifidobacterium longum 105-A, a strain with high transformation efficiency. Genome Announc. 2, e01311-14 (2014).
Sakanaka, M. et al. Functional analysis of bifidobacterial promoters in Bifidobacterium longum and Escherichia coli using the α-galactosidase gene as a reporter. J. Biosci. Bioeng. 118, 489–495 (2014).
Poolman, B., Nijssen, R. M. J. & Konings, W. N. Dependence of Streptococcus lactis phosphate transport on internal phosphate concentration and internal pH. J. Bacteriol. 169, 5373–5378 (1987).
Barri, T., Holmer-Jensen, J., Hermansen, K. & Dragsted, L. O. Metabolic fingerprinting of high-fat plasma samples processed by centrifugation- and filtration-based protein precipitation delineates significant differences in metabolite information coverage. Anal. Chim. Acta 718, 47–57 (2012).
Nielsen, L. N. et al. Glyphosate has limited short-term effects on commensal bacterial community composition in the gut environment due to sufficient aromatic amino acid levels. Environ. Pollut. 233, 364–376 (2018).
Roager, H. M. et al. Colonic transit time is related to bacterial metabolism and mucosal turnover in the gut. Nat. Microbiol. 1, 16093 (2016).
Smith, C. A., Want, E. J., O’Maille, G., Abagyan, R. & Siuzdak, G. XCMS: processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 78, 779–787 (2006).
Sumner, L. W. et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 3, 211–221 (2007).
Gürdeniz, G., Kristensen, M., Skov, T. & Dragsted, L. O. The effect of LC-MS data preprocessing methods on the selection of plasma biomarkers in fed vs. fasted rats. Metabolites 2, 77–99 (2012).
Pluskal, T., Castillo, S., Villar-Briones, A. & Orešič, M. MZmine 2: modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinformatics 11, 395 (2010).
van der Kloet, F. M., Bobeldijk, I., Verheij, E. R. & Jellema, R. H. Analytical error reduction using single point calibration for accurate and precise metabolomic phenotyping. J. Proteome Res. 8, 5132–5141 (2009).
Smart, K. F., Aggio, R. B. M., Van Houtte, J. R. & Villas-Bôas, S. G. Analytical platform for metabolome analysis of microbial cells using methyl chloroformate derivatization followed by gas chromatography–mass spectrometry. Nat. Protoc. 5, 1709–1729 (2010).
Johnsen, L. G., Skou, P. B., Khakimov, B. & Bro, R. Gas chromatography—mass spectrometry data processing made easy. J. Chromatogr. A 1503, 57–64 (2017).
Rosenmai, A. K. et al. Are structural analogues to bisphenol a safe alternatives? Toxicol. Sci. 139, 35–47 (2014).
Thysen, A. H. et al. Distinct immune phenotypes in infants developing asthma during childhood. Sci. Transl. Med. 12, 258 (2020).
R Core Team. R: A Language and Environment for Statistical Computing (R Foundation for Statistical Computing, 2013).
Wickhamm, H. ggplot2 Elegant Graphics for Data Analysis (Springer, 2009).
Vu, V. ggbiplot: a ggplot2 based biplot R v0.55 (2011).
Warnes, G. R. et al. gplots: various R programming tools for plotting data. R package v2.16.0 (2015).
Benjamini, Y. & Hochberg, Y. Controlling the False Discovery Rate—a practical and powerful approach to multiple testing. J. R. Stat. Soc. B 57, 289–300 (1995).
Smith, E. A. & Macfarlane, G. T. Enumeration of human colonic bacteria producing phenolic and indolic compounds: effects of pH, carbohydrate availability and retention time on dissimilatory aromatic amino acid metabolism. J. Appl. Bacteriol. 81, 288–302 (1996).
Smith, E. A. & Macfarlane, G. T. Formation of phenolic and indolic compounds by anaerobic bacteria in the human large intestine. Microb. Ecol. 33, 180–188 (1997).
Acknowledgements
We thank the children and families participating in the SKOT I study, which was supported by the Danish Directorate for Food, Fisheries and Agribusiness (grant no. 3304-FSE-06-0503), as well as the children and parents participating in the CIG cohort. Furthermore, we thank Aarstiderne A/S for providing a small gift for the CIG participants. We thank M. Danner Dalgaard at the Technical University of Denmark in-house facility (DTU Multi-Assay Core, DMAC) for performing the 16S rRNA gene sequencing, MS-Omics for performing the lactate analyses, Glycom A/S for kindly donating the HMOs and A. Schnipper for her efforts in supporting the work. We also thank S. Fukiya and A. Yokota (Research Faculty of Agriculture, Hokkaido University) for providing Bifidobacterium gene manipulation tools and for technical suggestions, Y. Sugiyama (Graduate School of Agriculture, Kyoto University) for helpful discussions on HPLC analysis, S. Maeda (Faculty of Bioresources and Environmental Sciences, Ishikawa Prefectural University) for technical support on enzyme purification, technician L. Buus Rosholm (Department of Biotechnology and Biomedicine, Technical University of Denmark) for performing the human monocyte experiments and technician B. Møller Plesning (National Food Institute, Technical University of Denmark) for running the AhR and HCAR-3 assays. We thank laboratory animal caretakers M. Danielsen, E. Erna Navntoft and K. Rene Worm (National Food institute, Technical University of Denmark). This work was supported by Augustinus Fonden (grant no. 17-2003 to H.M.R.); Hørslev Fonden (grant no. 203866 to H.M.R.); Beckett Fonden (grant no. 17-2-0551 to H.M.R.); Aase og Ejnar Danielsens Fond (grant no. 10-002019 to H.M.R.); the Innovation Fund Denmark (grant no. 11-116163/0603-00487B; Center for Gut, Grain and Greens to T.R.L.); JSPS-KAKENHI (18K14379 to M.S., 19K22277 to T.K.); JSPS Overseas Research Fellowships (201860637 to M.S.) and the Institute for Fermentation, Osaka (to M.S. and T.K.); ‘Diet-induced arrangement of the gut microbiome for improvement of cardiometabolic health’ (DINAMIC) under the Joint Programming Initiative, ‘A Healthy Diet for a Healthy Life’ (JPI-HDHL), supported by the Innovation Fund Denmark (grant no. 5195-00001B to L.O.D.); and ‘Biomarkers for infant fat mass development and nutrition’ under the ERA-HDHL joint transnational programme ‘Biomarkers for nutrition and health’, supported by the Innovation Fund Denmark (grant no. 4203-00005B to S.B.).
Author information
Authors and Affiliations
Contributions
H.M.R. and M.F.L. conceived and designed the study. M.F.L. prepared the samples for sequencing/qPCR and analysed the sequencing/qPCR data. H.M.R. prepared the samples for faecal/caecal and in vitro metabolome analyses and performed together with H.L.F. the targeted and untargeted metabolomics experiments. C.T.P. and L.O.D. performed the urine metabolomics. M.F.L. and M.S. performed the in vitro growth experiments, cloning and recombinant expression and mutant construction experiments. M.F.L. and H.M.R. performed the animal experiment with monocolonized mice. M.S. and T.K. performed enzyme kinetics. K.F.M. and C.M. designed the SKOT I study and M.V.L. managed the data. H.M.R. and M.F.L. designed the CIG cohort, recruited the study participants and managed the data. A.M.V. performed the AhR and HCA3 reporter assays. J.M.M. performed bioinformatics analysis of strain variation and MAGs. U.M. and S.B. performed ex vivo immune experiments. N.B., D.A. and A.R. contributed to the interpretation of the results. S.B., W.A., T.K., M.I.B. and T.R.L. contributed with expert supervision. H.M.R. and M.F.L. led the work, undertook the integrative data analyses and drafted the manuscript. All authors contributed to and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Nature Microbiology thanks Steven Frese and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Bifidobacterium species composition and aromatic lactic acids in partially breastfed and weaned infants aged 9 months from the SKOT cohort.
a, Pie chart of the average Bifidobacterium species composition given as percent of total Bifidobacterium abundance (n = 59 infants). Legend includes the average percent of each species compared to the total faecal microbiota community (See also Supplementary Data 1g). b, Left panel: Box and whiskers plot (line: median, box: IQR, whiskers: min-max) of faecal abundance of indolelactic acid (ILA), phenyllactic acid (PLA) and 4-hydroxyphenyllactic acid (4-OH-PLA) in partially breastfed (n = 24, red) and weaned (n = 35, blue) infants. Right panel: Box and whiskers plot (line: median, box: IQR, whiskers: min-max) of urine abundance of ILA, PLA and 4-OH-PLA in partially breastfed (n = 19, red) and weaned (n = 30, blue) infants. Statistical significance was evaluated by two-sided Mann–Whitney U-test. c, Heatmap of Spearman’s Rank correlation coefficients (two-sided tests) between the relative abundances of ILA, PLA and 4-OH-PLA measured in urine and the faecal relative abundance of Bifidobacterium species or faecal concentrations of ILA, PLA and 4-OH-PLA of the same infants (n = 49 infants). For all panels asterisks indicate statistical significance: *p < 0.05, **p < 0.01, ***p < 0.001 and ****p < 0.0001.
Extended Data Fig. 2 In vivo production of aromatic lactic acids in previously germ-free mice colonized with either B. longum 105-A WT or type 4 ldh mutant.
a, CFU counts of B. longum 105-A WT or type 4 ldh mutant from caecal content of mice monocolonized with either the WT (n = 21) or type 4 ldh mutant strain (n = 29). Bars and error bars indicate median ± IQR. b–d, caecal concentrations of the aromatic lactic acids (indolelactic acid (ILA), phenyllactic acid (PLA) and 4-hydroxyphenyllactic acid (4-OH-PLA)) in mice monocolonized with either the WT (n = 21) or type 4 ldh mutant strain (n = 29). Line and error bars indicate median ± IQR. For all panels statistical significance was evaluated by two-sided Mann–Whitney U-tests.
Extended Data Fig. 3 Faecal microbiota composition of the CIG infants.
a, Average relative abundance of the dominant faecal microbial taxa (average relative abundance >0.1%) across all samples (comprising 97.5% of all microbial taxa detected). b, Temporal development of the average gut microbiota composition and Shannon diversity index (marked with circles) across all individuals. c, Intra-individual temporal development of gut microbiota composition and Shannon diversity index (marked with circles). Dietary patterns and consumption of antibiotics are indicated for each individual. If nothing else is indicated, infants were singletons, vaginally born at term.
Extended Data Fig. 4 Beta diversity in CIG cohort.
a-f, Principal coordinates analysis (PCoA) plots of Bray–Curtis dissimilarities, based on all OTUs detected in CIG faecal samples (n = 234), coloured according to a, subject and b-f, relative abundances of Bifidobacterium longum, Bifidobacterium bifidum, Bifidobacterium breve, Bifidobacterium catenulatum group or Bifidobacterium dentium, respectively.
Extended Data Fig. 5 Temporal development in abundance of infant-type Bifidobacterium species, aromatic lactic acids, and human milk oligosaccharides in faeces from infants in the CIG cohort.
Scatter plots of a, age against absolute abundance of infant-type Bifidobacterium species (defined as the sum of absolute abundances of B. longum, B. breve, B. bifidum and B. scardovii) in faeces or b–d, age against faecal concentrations of aromatic lactic acids (ILA, PLA and 4-OH-PLA, n = 240) or e-g, age against relative faecal abundance of human milk oligosaccharides (2′FL/3FL, LNT/LNnT and 3′SL/6′SL, n = 228) during the first 6 months of life in the CIG cohort. A local polynomial regression (LOESS) fit is shown with coloured mean line and 95% CI shaded in grey. Statistical significance was evaluated by two-sided repeated measures correlations (rrm is the repeated measures correlation coefficient).
Extended Data Fig. 6 Associations between Infant-type Bifidobacterium species and aromatic lactic acids and human milk oligosaccharides in faeces from infants in the CIG cohort.
Scatter plots of the relationship between faecal absolute abundance of infant-type Bifidobacterium species, and a–c, faecal concentrations of aromatic lactic acids (ILA, PLA and 4-OH-PLA, n = 240) or d–f, relative faecal abundance of human milk oligosaccharides (2′FL/3FL, LNT/LNnT and 3′SL/6′SL, n = 228) in the CIG cohort. Statistical significance was evaluated by two-sided repeated measures correlations (rrm is the repeated measures correlation coefficient). Linear regression curve fits are shown with coloured mean line and 95% CI indicated in grey shading.
Extended Data Fig. 7 Correlations between relative abundance of bacterial taxa and concentrations of the aromatic lactic acids at each sampling point in the CIG cohort.
a, Spearman’s rank correlations between relative abundance of faecal bacterial genera (average relative abundance > 1%) and faecal concentrations of the aromatic lactic acids (ILA, PLA and 4-OH-PLA) at each sampling point. b, Spearman’s rank correlations between relative abundance of faecal Bifidobacterium species (average relative abundance > 0.1%) and faecal concentrations of the aromatic lactic acids at each sampling point. Infant-type Bifidobacterium species is the sum of the relative abundances of B. longum, B. breve, B. bifidum and B. scardovii. For both panels statistical significance was evaluated by uncorrected p-values (two-sided tests) indicated by asterisks with *p < 0.05, **p < 0.01, ***p < 0.001 and ****p < 0.0001.
Extended Data Fig. 8 Validation of associations between absolute abundance of Bifidobacterium species and faecal abundances of the aromatic lactic acids and human milk oligosaccharides in the CIG cohort.
Heatmaps illustrating linear mixed model coefficients (adjusted for subject and age) between the absolute abundance of Bifidobacterium species estimated by a, 16 S rRNA amplicon sequence variant analysis or b, species/subspecies-specific qPCR and faecal concentrations of aromatic lactic acids (ILA, PLA and 4-OH-PLA, n = 240) or relative faecal abundances of human milk oligosaccharides (2′FL/3FL, 3′SL/6′SL and LNT/LNnT, n = 228) in the CIG cohort. For both panels statistical significance was evaluated by false discovery rate-corrected p-values (two-sided tests) indicated by asterisks with *p < 0.05, **p < 0.01, ***p < 0.001 and ****p < 0.0001.
Extended Data Fig. 9 Relative abundance of high quality metagenome-assembled genomes (MAGs) with aldh (type 4 ldh) in 4 months and 12 months old infants and mothers.
Metagenome data were retrieved from Bäckhed et al [reference4]. Line, boxes and whiskers indicate median, IQR and ±1.5*IQR and statistical significance was evaluated by two-sided Mann–Whitney U-tests.
Extended Data Fig. 10 Absolute abundances of Bifidobacterium species and concentrations of aromatic lactic acids in faeces of infants in the CIG cohort.
a,b, Absolute abundance of Bifidobacterium species (average relative abundance >1% of total community) and concentrations of indolelactic acid (ILA), phenyllactic acid (PLA) and 4-hydroxyphenyllactic acid (4-OH-PLA) in faeces of individuals from the Copenhagen Infant Gut (CIG) cohort. Values of bacterial counts below 106 cells/g faeces and metabolite concentrations below 1 nmol/g faeces are not shown. Infant-type Bifidobacterium species is the sum of the absolute abundances of B. longum, B. breve, B. bifidum and B. scardovii and is indicated with grey background shading. a, Infants early colonized with infant-type Bifidobacterium species (colonized within first month reaching average relative abundance >40% during first 6 months), b, Infants with late colonization of infant-type Bifidobacterium species (not detectable or on average <0.5% of total community within the first 3 months of life).
Supplementary information
Supplementary Information
Supplementary Tables 1–10, Figs. 1–15 with images of uncropped gels and References.
Supplementary Data 1
SKOT infant cohort data.
Supplementary Data 2
CIG infant cohort data.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Source Data Fig. 4
Statistical source data.
Source Data Fig. 5
Statistical source data.
Source Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 1
Statistical source data.
Source Data Extended Data Fig. 2
Statistical source data.
Source Data Extended Data Fig. 3
Statistical source data.
Source Data Extended Data Fig. 4
Statistical source data.
Source Data Extended Data Fig. 5
Statistical source data.
Source Data Extended Data Fig. 6
Statistical source data.
Source Data Extended Data Fig. 7
Statistical source data.
Source Data Extended Data Fig. 8
Statistical source data.
Source Data Extended Data Fig. 9
Statistical source data.
Source Data Extended Data Fig. 10
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Laursen, M.F., Sakanaka, M., von Burg, N. et al. Bifidobacterium species associated with breastfeeding produce aromatic lactic acids in the infant gut. Nat Microbiol 6, 1367–1382 (2021). https://doi.org/10.1038/s41564-021-00970-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41564-021-00970-4
This article is cited by
-
Bacteroides uniformis degrades β-glucan to promote Lactobacillus johnsonii improving indole-3-lactic acid levels in alleviating colitis
Microbiome (2024)
-
A comparison of the breast milk microbiota from women diagnosed with gestational diabetes mellitus and women without gestational diabetes mellitus
BMC Pregnancy and Childbirth (2024)
-
Intestinal microbiota imbalance resulted by anti-Toxoplasma gondii immune responses aggravate gut and brain injury
Parasites & Vectors (2024)
-
Microbiota-derived indoles alleviate intestinal inflammation and modulate microbiome by microbial cross-feeding
Microbiome (2024)
-
Faecal microbiota and cytokine profiles of rural Cambodian infants linked to diet and diarrhoeal episodes
npj Biofilms and Microbiomes (2024)
