
Abstract
The kinetochore is a macromolecular structure that assembles on the centromeres of chromosomes and provides the major attachment point for spindle microtubules during mitosis. In Trypanosoma brucei, the proteins that make up the kinetochore are highly divergent; the inner kinetochore comprises at least 20 distinct and essential proteins (KKT1–20) that include four protein kinases—CLK1 (also known as KKT10), CLK2 (also known as KKT19), KKT2 and KKT3. Here, we report the identification and characterization of the amidobenzimidazoles (AB) protein kinase inhibitors that show nanomolar potency against T. brucei bloodstream forms, Leishmania and Trypanosoma cruzi. We performed target deconvolution analysis using a selection of 29 T. brucei mutants that overexpress known essential protein kinases, and identified CLK1 as a primary target. Biochemical studies and the co-crystal structure of CLK1 in complex with AB1 show that the irreversible competitive inhibition of CLK1 is dependent on a Michael acceptor forming an irreversible bond with Cys 215 in the ATP-binding pocket, a residue that is not present in human CLK1, thereby providing selectivity. Chemical inhibition of CLK1 impairs inner kinetochore recruitment and compromises cell-cycle progression, leading to cell death. This research highlights a unique drug target for trypanosomatid parasitic protozoa and a new chemical tool for investigating the function of their divergent kinetochores.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 digital issues and online access to articles
$119.00 per year
only $9.92 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout





Similar content being viewed by others


Data availability
Any data that support the findings of this study are available from the corresponding authors on request. T. brucei CLK1 kinase domain in complex with covalent aminobenzimidazole inhibitor AB1 is annotated at PDB under the accession number 6Q2A. Source data are provided with this paper.
References
Integrating Neglected Tropical Diseases into Global Health and Development: Fourth World Health Organization Report on Neglected Tropical Diseases (WHO, 2017).
Mesu, V. K. B. K. et al. Oral fexinidazole for late-stage African Trypanosoma brucei gambiense trypanosomiasis: a pivotal multicentre, randomised, non-inferiority trial. Lancet 391, 144–154 (2018).
Dickie, E. A. et al. New drugs for human African trypanosomiasis: a twenty first century success story. Trop. Med. Infect. Dis. 5, 29 (2020).
Khare, S. et al. Proteasome inhibition for treatment of leishmaniasis, Chagas disease and sleeping sickness. Nature 537, 229–233 (2016).
Wyllie, S. et al. Cyclin-dependent kinase 12 is a drug target for visceral leishmaniasis. Nature 560, 192–197 (2018).
Rao, S. P. S. et al. Drug discovery for kinetoplastid diseases: future directions. ACS Infect. Dis. 5, 152–157 (2019).
Myburgh, E. et al. In vivo imaging of trypanosome-brain interactions and development of a rapid screening test for drugs against CNS stage trypanosomiasis. PLoS Negl. Trop. Dis. 7, e2384 (2013).
Lelais, G. et al. Discovery of (R,E)-N-(7-chloro-1-(1-[4-(dimethylamino)but-2-enoyl]azepan-3-yl)-1H-benzo[d]imidazol-2-yl)-2-methylisonicotinamide (EGF816), a novel, potent, and WT sparing covalent inhibitor of oncogenic (L858R, ex19del) and resistant (T790M) EGFR mutants for the treatment of EGFR mutant non-small-cell lung cancers. J. Med. Chem. 59, 6671–6689 (2016).
Parsons, M., Worthey, E. A., Ward, P. N. & Mottram, J. C. Comparative analysis of the kinomes of three pathogenic trypanosomatids: Leishmania major, Trypanosoma brucei and Trypanosoma cruzi. BMC Genom. 6, 127 (2005).
Begolo, D., Erben, E. & Clayton, C. Drug target identification using a trypanosome overexpression library. Antimicrob. Agents Chemother. 58, 6260–6264 (2014).
McNamara, C. W. et al. Targeting Plasmodium PI(4)K to eliminate malaria. Nature 504, 248–253 (2013).
Moeslein, F. M., Myers, M. P. & Landreth, G. E. The CLK family kinases, CLK1 and CLK2, phosphorylate and activate the tyrosine phosphatase, PTP-1B. J. Biol. Chem. 274, 26697–26704 (1999).
Jones, N. G. et al. Regulators of Trypanosoma brucei cell cycle progression and differentiation identified using a kinome-wide RNAi screen. PLoS Pathog. 10, e1003886 (2014).
Akiyoshi, B. & Gull, K. Discovery of unconventional kinetochores in kinetoplastids. Cell 156, 1247–1258 (2014).
Nishino, M., Choy, J. W., Gushwa, N. N. & Oses-Prieto, J. A. Hypothemycin, a fungal natural product, identifies therapeutic targets in Trypanosoma brucei. eLife 9, e00712 (2013).
Liu, Q. et al. Developing irreversible inhibitors of the protein kinase cysteinome. Chem. Biol. 20, 146–159 (2013).
Westhorpe, F. G. & Straight, A. F. Chromosome segregation: reconstituting the kinetochore. Curr. Biol. 26, R1242–R1245 (2016).
D’Archivio, S. & Wickstead, B. Trypanosome outer kinetochore proteins suggest conservation of chromosome segregation machinery across eukaryotes. J. Cell Biol. 216, 379–391 (2017).
Kabani, S., Waterfall, M. & Matthews, K. R. Cell-cycle synchronisation of bloodstream forms of Trypanosoma brucei using Vybrant DyeCycle Violet-based sorting. Mol. Biochem. Parasitol. 169, 59–62 (2010).
Torrie, L. S. et al. Identification of inhibitors of an unconventional Trypanosoma brucei kinetochore kinase. PLoS ONE 14, e0217828 (2019).
Ishii, M. & Akiyoshi, B. Characterization of unconventional kinetochore kinases KKT10/19 in Trypanosoma brucei. J. Cell Sci. 133, jcs240978 (2020).
Mackey, Z. B., Koupparis, K., Nishino, M. & McKerrow, J. H. High-throughput analysis of an RNAi library identifies novel kinase targets in Trypanosoma brucei. Chem. Biol. Drug Des. 78, 454–463 (2011).
Roskoski, R. Properties of FDA-approved small molecule protein kinase inhibitors. Pharmacol. Res. 144, 19–50 (2019).
Wyllie, S. et al. Preclinical candidate for the treatment of visceral leishmaniasis that acts through proteasome inhibition. Proc. Natl Acad. Sci. USA 116, 9318–9323 (2019).
Alam, M. M. et al. Validation of the protein kinase PfCLK3 as a multistage cross-species malarial drug target. Science 365, eaau1682 (2019).
van Hooff, J. J., Tromer, E., van Wijk, L. M., Snel, B. & Kops, G. J. Evolutionary dynamics of the kinetochore network in eukaryotes as revealed by comparative genomics. EMBO Rep. 18, 1559–1571 (2017).
Duncan, S. M. et al. Conditional gene deletion with DiCre demonstrates an essential role for CRK3 in Leishmania mexicana cell cycle regulation. Mol. Microbiol. 100, 931–944 (2016).
Buckner, F. S., Verlinde, C. L., La Flamme, A. C. & Van Voorhis, W. C. Efficient technique for screening drugs for activity against Trypanosoma cruzi using parasites expressing beta-galactosidase. Antimicrob. Agents Chemother. 40, 2592–2597 (1996).
Alsford, S., Kawahara, T., Glover, L. & Horn, D. Tagging a T. brucei RRNA locus improves stable transfection efficiency and circumvents inducible expression position effects. Mol. Biochem. Parasitol. 144, 142–148 (2005).
Dean, S., Sunter, J. D. & Wheeler, R. J. TrypTag.org: a trypanosome genome-wide protein localisation resource. Trends Parasitol. 33, 80–82 (2017).
Kaiser, M. et al. Antitrypanosomal activity of fexinidazole, a new oral nitroimidazole drug candidate for treatment of sleeping sickness. Antimicrob. Agents Chemother. 55, 5602–5608 (2011).
Alsford, S. & Horn, D. Single-locus targeting constructs for reliable regulated RNAi and transgene expression in Trypanosoma brucei. Mol. Biochem. Parasitol. 161, 76–79 (2008).
Thuita, J. K. et al. Efficacy of the diamidine DB75 and its prodrug DB289, against murine models of human African trypanosomiasis. Acta Trop. 108, 6–10 (2008).
Waters, N. J., Jones, R., Williams, G. & Sohal, B. Validation of a rapid equilibrium dialysis approach for the measurement of plasma protein binding. J. Pharm. Sci. 97, 4586–4595 (2008).
Kalvass, J. C., Tess, D. A., Giragossian, C., Linhares, M. C. & Maurer, T. S. Influence of microsomal concentration on apparent intrinsic clearance: implications for scaling in vitro data. Drug Metab. Dispos. 29, 1332–1336 (2001).
Winter, G. Xia2: an expert system for macromolecular crystallography data reduction. J. Appl. Crystallogr. 43, 186–190 (2010).
Kabsch, W. XDS. Acta Crystallogr. D 66, 125–132 (2010).
Evans, P. R. & Murshudov, G. N. How good are my data and what is the resolution? Acta Crystallogr. D 69, 1204–1214 (2013).
McCoy, A. J. et al. Phaser crystallographic software. J. Appl. Crystallogr. 40, 658–674 (2007).
Walter, A. et al. Molecular structures of cdc2-like kinases in complex with a new inhibitor chemotype. PLoS ONE 13, e0196761 (2018).
Adams, P. D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D 66, 213–221 (2010).
Emsley, P., Lohkamp, B., Scott, W. G. & Cowtan, K. Features and development of Coot. Acta Crystallogr. D 66, 486–501 (2010).
Acknowledgements
We thank our colleagues at The Bioscience Technology Facility of University of York who provided insight and expertise that assisted our microscopy and flow cytometry research; and G. Lelais, V. Manoharan, R. Palkar, V. Lim, C. Noble and W. K. Fei for their technical support. This research was supported by the Wellcome Trust (nos. 069712, 103024 and 108517). J.C.M. is a Wellcome Trust Investigator (no. 200807). M.P.B. and J.C.M. are part of the Wellcome Centre of Integrative Parasitology (104111/Z/14/Z).
Author information
Authors and Affiliations
Contributions
J.C.M., T.T.D. and S.P.S.R. planned the studies. Compound synthesis, compound docking, protein kinase assays and library screening were performed by H.X.Y.K., D.Patra, Y.-L.C., S.B.L., S.W., F.S. and J.J.; HAT animal studies were performed by E.M., R.R., M.P.B. and M.K.; individual protein kinase cell lines were generated by E.B. and M.S.; compound target deconvolution, TbCLK1 functional characterization, immunofluorescence and data analyses were performed by M.S. and J.B.T.C.; protein recombinant production, kinase assays and MS were performed by E.F., D.Patra, C.B.-L. and M.S.; crystallization and structure determination were performed by E.O. and X.M.; CLK1 mutant plasmids were designed and prepared by M.S., D.Paape and R.M.; J.C.M., S.P.S.R. and M.S. prepared and wrote the manuscript. All of the authors reviewed, edited and approved the paper. J.C.M., S.P.S.R., M.P.B. and T.T.D. obtained funding.
Corresponding authors
Ethics declarations
Competing interests
S.P.S.R., T.T.D., J.J., F.S., Y.-L.C., S.B.L. and X.M. have shares in Novartis. All other authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 High-throughput screening of kinase library and characterization of AB1 effect on T. brucei.
a, Identification of amidobenzimidazole (AB) series as a scaffold worthy of progression. Single point screen of ten thousand compounds from kinase library were screened against T. b. brucei parasite for growth inhibition at 10 µM. 2264 cpds showed >50% growth inhibition, of which 306 compounds were prioritized based on the cluster analysis and compound availability for 10 point dose response testing. 232 cpds showed <10 µM EC50, further analysis based on physicochemical properties [calculated octanol-water partition coefficient (cLogP) < 4.5; polar surface area (PSA) < 110; molecular weight (MW) < 500], removal of unfavourable chemical moeities and selectivity index of >10 (cytotoxicity in human hepatocytes HepG2 CC50 / T. b. brucei EC50) lead to identification of amidobenzimidazole series. b, Time to kill analysis of AB1 compound. Note the concentration and time dependent killing of bloodstream form of T. b. brucei parasites by the AB1 compound. Data shown represent the mean ± SEM (n = 3 independent biological replicates). c, The absolute concentrations (ACcure) required to achieve sterile cure under in vitro conditions with incubation of compound at various time points. Note that as the time of incubation increases, ACcure reduces. Mean data from two independent biological replicates are shown.
Extended Data Fig. 2 AB1 reduces parasitaemia in a GVR35 strain CNS mouse model of human African trypanosomiasis and pharmacokinetics of AB1.
a, Whole body in vivo imaging of bioluminescent T. brucei before and after AB1 treatment; T. b. brucei (GVR35-VSL2) –infected mice (Day 21) were orally treated with 100 mg kg−1 AB1 once-daily for 7 days (n = 6 mice, grey) or left untreated (n = 3 mice, green); Data represent the mean ± standard deviation of each mice group, and symbols show whole body bioluminescence values for individual mice; mice were euthanized between days 27 and 35. b, Bioluminescence images of mice showing ventral views of two representative mice from the untreated and AB1-treated groups over the full course of infection. The colour scale indicates bioluminescent radiance in photons/sec/cm2/sr. ND, not detected. Blood parasitemia (in parasites mL−1, red font below image) and whole mouse total flux (in photons per second, black font above image) values of each animal are shown. The same two representative mice are shown for all time points. c, Total blood concentration of AB1 in mice at various time points after last dose from the T. b. brucei GVR35 strain CNS mouse model of HAT. At each time point, 3 mice were bled to collect samples, each point represents mean ± standard deviation. d, Free brain concentrations of AB1 compound in mice calculated by taking into consideration the brain to plasma ratio (0.5), mice plasma protein binding (94%) and rat brain tissue binding (>99%). Each point represents free brain concentrations for n = 3 mice and mean ± standard deviation is shown. Note the AB1 concentrations were below T. b. brucei EC50 and ACcure.
Extended Data Fig. 3 CLK1 target validation and CLK1 over-expression toxicity blockage by AB1.
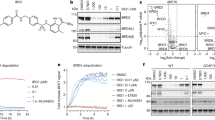
a, CLK1 depletion halts cell proliferation. Growth curves of CLK1-depleted bloodstream trypanosomes after RNAi induction with tetracycline (dashed line) is compared with uninduced (solid line) as control. Data represent the mean of two biological replicates. b, Efficiency of CLK1 RNAi. Upper panel: Endogenous CLK1::12-myc (predicted MW 70.5 kDa) was detected with anti-myc antibody at different points after RNAi induction. EF-1 alpha protein expression was used as the loading control. Lower panel: Mean intensity of the bands were determined from the Western blot in the upper panel. Density signal of CLK1::12-myc was normalised to EF1α for each time point and the uninduced control and relative expression of CLK1::12-myc to uninduced control is shown. Data are representative from one of two independent biological replicates with similar results. c, Depletion of CLK1 induces a G2/M cell cycle arrest. Flow cytometry analysis of DNA content in CLK1-depleted cells (dark grey) and in the uninduced control (light grey) 24 and 48 h after RNAi induction with 1 μg mL-1 tetracycline. Mean ± SEM (n = 3); P values were calculated using a Two-tailed Student’s t-tests comparing G2/M percentage with uninduced control where ** p = 2.7 ×10-3 (24 h), *** p = 1.6 ×10-4 (48 h). d, CLK1 overexpression impairs parasite growth. Top: 48 h growth curve of induced TbCLK1 over-expression (OE) compared with uninduced cell line and parental T. b. brucei 2T1 wild-type parasites. Data represent the mean ± standard deviation of three biological replicates where P-values were calculated using a Two-tailed Student’s t-test; *** p = 5.9 ×10-5. Bottom: CLK1-6-myc overexpression of two clones was assessed by western blot, using an anti-myc antibody. EF-1 alpha protein expression was used as the loading control. e, Incubation with AB1 impairs toxicity of CLK1 overexpression. Parental 2T1 and CLK1 over-expression (OE) cell lines were pre-incubated or not 18 h with 60 nM of AB1 and AB2 and then incubated with tetracycline during 48 h. Bars show the mean ± SEM (n = 3 biological replicates), and symbols represent the individual parasite counts. *** p = 1.6 ×10-4 compared to induced CLK1 OE preincubated with DMSO. f, CLK1 target validation using non-toxic concentrations of AB1. 24 h pre-treatment with AB1 non-toxic concentrations (60 nM) reduced the loss of fitness caused by CLK1 overexpression induced by tetracycline. Bars represent the mean ± SEM (n = 4 biological replicates) of the percentage of maximal survival in each condition, and symbols represent the individual replicates. p < 0.001 compared to DMSO control for all the points.
Extended Data Fig. 4 CLK1 and CLK2 protein sequence alignment and CLK1 conservation across trypanosomatids.
a, Alignment of CLK1/KKT10 and CLK2/KKT19 protein sequence in T. brucei. The protein sequences of CLK1 (Tb927.11.12410) and CLK2 (Tb927.11.12420) were aligned in CLC Genomics Workbench 8 and the consensus graph is shown below the corresponding alignment. Residue Cysteine 215 is shown in blue. b, Left: CLK1 and CLK2 mRNA levels measured by quantitative RT-PCR (bar graphs) in the 2T1 parental cell line, using GADPH as housekeeping gene. Mean ± SEM (n = 3 biological replicates). Right: Protein levels of Ty-YFP-tagged KKT10 and KKT19 were assessed by western blot. Representative of two independent clones is shown. EF-1 alpha protein expression was used as the loading control. c, The protein sequences of CLK1/KKT10 (Tb927.11.12410) and orthologues from Leishmania mexicana (LmxM.09.0400) and Trypanosoma cruzi (TcCLB.509027.60) were aligned in CLC Genomics Workbench 8. The consensus graph is shown below the corresponding alignment. Conserved residue Cysteine 215 is shown in blue.
Extended Data Fig. 5 Similarity of hEGFR and CLK1 binding sites highlighting potential of AB1 to covalently bind to TbCLK1.
a Sequence alignment of hEGFR (Uniprot id: P00533), hCLK1 (Uniprot id: P49759) and TbCLK1 show similarity in binding site residues with presence of covalent cysteine (red) in TbCLK1 explaining selectivity over hCLK1. b Comparison of ribbon diagram x-ray crystal structure of hCLK1 (PDB: 6FT8), TbCLK1 (PDB: 6Q2A), and hEGFR (5FEQ). The kinase components are coloured and labelled in TbCLK1 (middle panel). Note the conservation of cysteine in the kinase domain in TbCLK1 (C215) and hEGFR (C797), whereas in the same position, hCLK1 has serine 249. c An overlay of TbCLK1 with hCLK1 (left) and hEGFR (right) structures.
Extended Data Fig. 6 Covalent binding of AB1 to TbCLK1.
a, Summary of thermal shift assay of TbCLK1 and TbCLK1 C215A proteins incubated with DMSO, AB0 or AB1 compounds. Data represents mean ± standard deviation (n = 3 technical replicates) for △Tm. Note the significant increase in Tm for AB1 compound bound to CLK1 but not CLK1 C215A. The Tm was calculated by fitting the fluorescence vs. temperature data to the Boltzmann equation and represents the point where half the protein is unfolded. b, Mass spectrum chromatograms of TbCLK1 (left) and TbCLK1 C215A proteins (right) incubated with DMSO (control), AB1 or AB2 compounds. Y-axis and X-axis represent the counts and deconvoluted Mass (amu) respectively. Calculated differences in base mass and the difference in mass after treatment is shown in the table. Note the increase in mass of 475 Da only for AB1 treatment with TbCLK1 protein, which corresponds to the mass of AB1. No increase in the mass was seen for DMSO or AB2 in both CLK1 and CLK1 C215A protein. Treatment of CLK1 C215A with AB1 also did not result in an increase in mass, clearly showing covalent interaction of the Michael acceptor with C215 of TbCLK1 (n = 1).
Extended Data Fig. 7 Expression of CLK1 C215A mutant in T. brucei.
a, Expression of CLK1 C215A results in a G2/M cell cycle arrest. Induction of recoded CLK1 C215A and C215S mutants were expressed for 24 h with tetracycline and cell cycle distribution was determined by flow cytometry and compared with 2T1 parental cell line. Bars represent the mean ± SEM (n = 3 independent experiments). P values were calculated using a Two-tailed Student’s t-test comparing G2/M phase with parental 2T1 control where *** p = 6.0 ×10-6. b, Constitutive expression of recoded CLK1 C215S halts CLK1 RNAi growth inhibition. Parasites were induced or not with 1 μg mL-1 tetracycline at 1 ×105 parasites ml-1 and daily diluted up to 48 h, where ~ 25% reduction in parasite growth was observed. Data represent mean ± SEM (n = 4 technical replicates) showing cumulative cell growth across time. The P values were calculated using a two-tailed Student’s t-test comparing the recoded mutant with parental uninduced control where *** p = 7.9 ×10-4. A representative Western blot of expression of recoded CLK1 C215S mutant is shown in the right panel. The expression of recoded CLK1 was detected using an anti-HA antibody. EF-1 alpha protein expression was used as the loading control. c, CLK1 C215S mutant impairs the parasite growth effect of 48 h with 5x AB1 EC50 treatment. Growth curve of parasites expressing recoded CLK1 C215S (green) and parental 2T1 cell line (blue) (5 ×105 parasites ml-1) treated with 5x AB1 EC50 for 48 h. Data represents mean ± SEM (n = 3 technical replicates). *** P values were calculated using a Two-tailed Student’s t-test comparing the recoded CLK1 C215S cell line with parental control at each time point, 18 h (p = 3.0 ×10-6), 24 h (p = 1.1 ×10-4) and 48 h (p = 2.5 ×10-7). d, T. brucei expressing recoded HA-tagged TbCLK1 and C215S mutant were incubated induced or not with 1 μg mL-1 tetracycline for 24 h and then treated with 5x EC50 AB1 for 6 h. The expression of recoded CLK1 was detected using an anti-HA antibody. EF-1α protein expression was used as the loading control. The size increase of CLK1 is showed with an arrow. Data shown is representative from one of the three biological replicates with similar results.
Extended Data Fig. 8 Cell cycle analysis of synchronized T. brucei after treatment with AB1.
a, Diagram of 2C (blue) and 4C (orange) cell cycle synchronised bloodstream form T. brucei using Vybrant DyeCycle Violet-based cell sorting. The selected area shows those cells that were gated for sorting. b, c, After release, 2C and 4C synchronised parasites were treated or not with 5x EC50 AB1. Then, cell samples were collected every hour for DNA flow cytometry analysis of 2C (in blue) and 4C (orange) populations, up to 6 hours. A representative micrograph after 6 hours of treatment is also provided in each condition. Scale bar, 2μm. The data are representative of two biological experiments.
Supplementary information
Supplementary Information
Supplementary Methods and Supplementary Table 1 (containing data collection and refinement statistics).
Source data
Source Data Fig. 1
Source Data for Fig. 1a,c.
Source Data Fig. 2
Source Data for Fig. 2b–e.
Source Data Fig. 4
Source Data for Fig. 4a–e.
Source Data Fig. 4
Unprocessed western blot for Fig. 4c.
Source Data Fig. 5
Source Data for Fig. 5a.
Source Data Extended Data Fig. 1
Source Data for Extended Data Fig. 1b,c.
Source Data Extended Data Fig. 2
Source Data for Extended Data Fig. 2a,c,d.
Source Data Extended Data Fig. 3
Source Data for Extended Data Fig. 3a–f.
Source Data Extended Data Fig. 3
Unprocessed western blot for Extended Data Fig. 3b,d.
Source Data Extended Data Fig. 4
Source Data for Extended Data Fig. 4b.
Source Data Extended Data Fig. 4
Unprocessed western blot for Extended Data Fig. 4b.
Source Data Extended Data Fig. 6
Source Data for Extended Data Fig. 6a.
Source Data Extended Data Fig. 7
Source Data for Extended Data Fig. 7a–c.
Source Data Extended Data Fig. 7
Unprocessed western blot for Extended Data Fig. 7b (left) and d (right).
Source Data Extended Data Fig. 8
Source Data for Extended Data Fig. 8a,b.
Source Data Fig. 3
Source Data for Fig. 3a,b.
Rights and permissions
About this article

Cite this article
Saldivia, M., Fang, E., Ma, X. et al. Targeting the trypanosome kinetochore with CLK1 protein kinase inhibitors. Nat Microbiol 5, 1207–1216 (2020). https://doi.org/10.1038/s41564-020-0745-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41564-020-0745-6
This article is cited by
-
Genome-wide subcellular protein map for the flagellate parasite Trypanosoma brucei
Nature Microbiology (2023)
-
Anti-trypanosomatid drug discovery: progress and challenges
Nature Reviews Microbiology (2023)
-
CLK1/CLK2-driven signalling at the Leishmania kinetochore is captured by spatially referenced proximity phosphoproteomics
Communications Biology (2022)
-
Genome-scale RNA interference profiling of Trypanosoma brucei cell cycle progression defects
Nature Communications (2022)
-
Systematic functional analysis of Leishmania protein kinases identifies regulators of differentiation or survival
Nature Communications (2021)
