
Abstract
Archaeological and archaeogenetic evidence points to the Pontic–Caspian steppe zone between the Caucasus and the Black Sea as the crucible from which the earliest steppe pastoralist societies arose and spread, ultimately influencing populations from Europe to Inner Asia. However, little is known about their economic foundations and the factors that may have contributed to their extensive mobility. Here, we investigate dietary proteins within the dental calculus proteomes of 45 individuals spanning the Neolithic to Greco-Roman periods in the Pontic–Caspian Steppe and neighbouring South Caucasus, Oka–Volga–Don and East Urals regions. We find that sheep dairying accompanies the earliest forms of Eneolithic pastoralism in the North Caucasus. During the fourth millennium bc, Maykop and early Yamnaya populations also focused dairying exclusively on sheep while reserving cattle for traction and other purposes. We observe a breakdown in livestock specialization and an economic diversification of dairy herds coinciding with aridification during the subsequent late Yamnaya and North Caucasus Culture phases, followed by severe climate deterioration during the Catacomb and Lola periods. The need for additional pastures to support these herds may have driven the heightened mobility of the Middle and Late Bronze Age periods. Following a hiatus of more than 500 years, the North Caucasian steppe was repopulated by Early Iron Age societies with a broad mobile dairy economy, including a new focus on horse milking.
Similar content being viewed by others

Main
During the early to mid-Holocene (ca. 9.0–3.5 thousand years ago (kya)), dairying played a vital role in the development of human food systems across Europe, Africa and Asia1,2,3,4,5,6,7,8. Early agropastoral societies raised livestock animals that could provide them with milk, meat, wool, leather and traction9, and milk rose to prominence as an especially important, nutrient-rich food source. Milk is rich in protein, fat, sugar (lactose), vitamins and minerals, such as calcium10, and the water content in milk can be relied on in times of drought or scarcity11,12. Although milk itself is highly perishable, it can be transformed through microbial fermentation and other forms of manipulation into more stable products, such as yogurt, butter, ghee, cheese and curds, that can be stored for longer periods in surplus13,14,15.
First attested in Anatolia during the seventh and sixth millennia bc3,6, ruminant dairying subsequently spread to both Europe and Africa by the late sixth millennium bc4,16, but less is known about its initial dispersals into Asia17,18,19. One major vector by which dairying spread was the Eurasian steppe, an enormous expanse of grasslands stretching 6,000 km from the Carpathian Basin to Mongolia. Recent studies have traced the introduction of dairying in Mongolia to ca. 3000 bc with the appearance of mobile steppe herders associated with the Early Bronze Age Afanasievo culture2, a group with close genetic and cultural ties to pastoralists on the Pontic–Caspian steppe, most notably the Yamnaya culture (ca. 3300–2500 bc)20,21,22,23. Populations from the Pontic–Caspian steppe are also linked to Late Neolithic and Bronze Age westward expansions, including the emergence of the Corded Ware (2900–2200 bc) and Bell Beaker (2750–1800 bc) phenomena in Europe24,25,26,27. Understanding the population and economic history of the Pontic–Caspian steppe, the source region for these continental-scale expansions during the third millennium bc, is critical for revealing the main factors that drove the heightened mobility of Eneolithic and Early Bronze Age pastoralists in Eurasia.
When Pontic–Caspian steppe populations first began dairying and how their animal management strategies may have influenced their mobility and subsequent migrations remain poorly known. From the Mesolithic through the Eneolithic, populations living in the southern Russian plain and Caucasus region primarily hunted local wild game, which included aurochs (Bos primigenius), saiga antelope (Saiga tatarica), red deer (Cervus elaphus), tarpan (Equus ferus), onager (Equus hemionus) and wild boar (Sus scrofa), as well as birds, fish and molluscs28,29,30,31. Animal husbandry of domesticated sheep (Ovis aries), goats (Capra hircus), cattle (Bos taurus) and pigs (Sus scrofa) spread to the North Caucasian steppe from Anatolia during the fifth millennium bc by either a circum-Pontic route28 or by crossing the Caucasus mountains from the south32,33,34,35. By the mid-fifth millennium bc, agropastoralists of the Cucuteni–Trypillia culture in Ukraine were regularly interacting with steppe populations north of the Black Sea36, and Eneolithic populations genetically related to South Caucasian and Anatolian agropastoralist groups had become established in the North Caucasus piedmont steppe32,33,37 and were part of a broader Mesopotamian interaction sphere38,39.
After the introduction of animal husbandry to the region, Bronze Age steppe populations innovated a new economic system of mobile pastoralism focused on sheep and cattle40, and settlements became effectively absent on the steppe for the next two millennia40,41. This new, more mobile form of pastoralism is first evident among Steppe Late Maykop groups (3500–2900 bc), who fall broadly within the Late Maykop cultural sphere but are genetically distinct from their higher-elevation counterparts33, and fully mobile pastoralism subsequently became the predominant subsistence strategy on the steppe with the Yamnaya culture (3,300–2,500 bc)41. Horse domestication occurred during the third millennium on the Pontic–Caspian steppe42,43, and, by the late third and early second millennium bc, domestic horses were increasingly part of the steppe mobile pastoralist economy44 and had even spread to Anatolia and Mesopotamia through Pontic–Caspian–Transcaucasian interaction networks45. Mobile pastoralism continued among the Catacomb (2800–2200 bc) and North Caucasus Culture (NCC; 2800–2400 bc) groups in the steppe until worsening climatic conditions and aridification ca. 2300–2200 bc, in association with the 4.2 kya climate event46,47, ultimately led to an abandonment of the steppe region by 1700 bc40,41. Despite their cultural differences, recent palaeogenomic analysis has shown that these Bronze Age steppe populations were genetically highly similar33, which may, in part, reflect their mobile lifestyles and persistent multicultural interactions over millennia40.
Throughout the Pontic–Caspian steppe, sheep, goat and cattle dominate most studied steppe archaeofaunal collections from the fourth to second millennia bc41,48,49. Wheeled transport in the form of wagons first appears in kurgans (burial mounds) of the Steppe Late Maykop in the second half of the fourth millennium bc50, and such technology is argued to be essential for enabling the household mobility required for mobile pastoralism40. Oxen teams dated to the same period and, later, horses and chariots in the second millennium bc, further facilitated mobility51. Sheep wool was present in the North Caucasus by the early third millennium bc, possibly having originated in Anatolia, and the use of wool subsequently spread across the steppe and into Inner Asia during the second millennium bc52. Among the region’s major secondary products, dairying is argued to have possibly emerged first50, in part because dairying was already well established in both Anatolia and surrounding regions by the sixth millennium bc6,53,54,55, whereas evidence for traction and wool are only attested millennia later. Nevertheless, current evidence for early dairying in the Pontic–Caspian steppe is, until now, only attested on its eastern fringes7. Previous isotopic studies have been unable to identify clear indications of dairy consumption, finding instead non-specific evidence for high consumption of animal protein and a highly complex isoscape, reflecting both ecological diversity and temporal climatic shifts41,48,56. However, the isotopic data suggest a stronger contribution of sheep or goat products to the human diet than those from cattle41. Few zooarchaeological studies have systematically investigated herd management and mortality profiles in the region, but the earliest agropastoralist communities in the North Caucasus piedmont steppe were not thought to have engaged in dairying49. Likewise, there are few indications of animal management for milk production among Neolithic agropastoralist communities in the South Caucasus42. Rather, it is only in the second millennium bc that zooarchaeological studies from Late Bronze Age settlements in the Caucasus have found clear evidence for the deliberate keeping of sheep for milk production57,58, and it is only later during the Iron Age that cattle show mortality profiles consistent with dairying59.
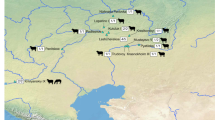
The absence of settlements on the steppe and the near-exclusive archaeological focus on mortuary contexts have made it difficult to reconstruct the nature and extent of dairying in the wider North Caucasian pastoralist economy. In this article, we apply high-resolution tandem mass spectrometry to human dental calculus from 45 individuals at 29 sites in the North Caucasus (n = 27) and the neighbouring South Caucasus (n = 9), Oka–Volga–Don (n = 7) and East Urals (n = 2) regions (Fig. 1a,b, Supplementary Data 1 and Supplementary Information) to identify evidence of consumed dairy proteins in populations spanning the Neolithic to the Greco-Roman periods (ca. 6000 bc to 200 ad). We find that dairy products were consumed in the North Caucasus from the late fifth millennium bc onwards and that a dairy-inclusive subsistence characterizes even the Eneolithic populations in the piedmont and steppe zones. Dairy consumption was prevalent for all analysed periods and ecotones in the North Caucasus, with milk proteins identified in 26 of 27 tested individuals. We identify an initial, near-exclusive dairying focus on sheep among the Maykop, Steppe Maykop and early Yamnaya, followed by diversification within the late Yamnaya, NCC and Catacomb cultures during the Middle Bronze Age to additionally incorporate goat and cattle milking. Later, during the Early Iron Age, we observe direct evidence of horse milk consumption in association with pre-Scythian groups repopulating the steppe after a centuries-long hiatus. In the South Caucasus, we identify evidence of cattle milking (ca. 3700 bc) nearly 1,000 years before we first observe it in the North Caucasus (ca. 2700 bc), and, in the Oka–Volga–Don region, we observe limited evidence of dairying, beginning only during the second millennium bc.

a, Map of study area and major cultural regions mentioned in the text: Oka–Volga–Don, East Urals, North Caucasus, South Caucasus and Anatolia. Extent of the Pontic–Caspian steppe is shown in grey. Inset: enhanced view of North Caucasus sites. b, Timeline of sites and individuals analysed in this study. Individuals are organized by region, with archaeological culture or period indicated by colour corresponding to the legend. White circles indicate median calibrated radiocarbon dates, and error bars are 2 s.d. Coloured bars display the time spans conventionally associated with the archaeological cultures and time periods. c, Milk protein evidence by individual, displayed as total PSM count to the milk proteins BLG, alpha-lactalbumin and alpha-S1-casein. Consensus livestock assignment was determined by parsimony. aTwo dental calculus samples were analysed from ZO2002. Basemap is from https://www.naturalearthdata.com/.
Results
Milk proteins were identified in 34 of 45 analysed individuals across all time periods (Fig. 1c and Supplementary Data 1). Protein recovery in 31 individuals was sufficient to allow the identification of major ruminant livestock milks from sheep (Ovis), goat (Capra) and/or cattle (Bos/Bovinae), whereas the milk proteins of three individuals were represented by non-specific bovid peptides, indicating either sheep or cattle. Additionally, one individual had taxonomically distinctive peptide spectral matches (PSMs) to Equus milk proteins. Beta-lactoglobulin (BLG), which was detected for all dairy livestock (Fig. 2), was the most prevalent and abundant milk protein detected, a pattern consistent with previous studies of dental calculus2,6,60. In addition to BLG, which was identified in all 34 milk-positive individuals, we also identified the whey protein alpha-lactalbumin in two individuals and the curd protein alpha-S1-casein in two individuals. All dental calculus samples yielded proteomes consistent with an oral microbiome profile, and age-associated N/Q deamidation was a top modification across the dataset (Supplementary Data 2). Milk protein peptide-level deamidation is reported in Supplementary Data 3. No dietary proteins were detected in non-template extraction controls (Supplementary Data 3).

a, Overall, most BLG sequences were highly conserved among bovids (left) but distinct from equids (right). Spectra originate from AY2005 and MK5018. b, Among bovids, the BLG C-terminus peptide distinguishes caprines (left) and bovines (right). Spectra originate from VS2001 and VS2001. c, The most frequently observed peptide reliably distinguishes Ovis (upper left), Capra (lower left) and Equus (lower right) but cannot distinguish Ovis and Bovinae due to the ambiguity of the sixth residue, which may be aspartic acid (Bovinae) or deamidated asparagine (Ovis)6 (upper right). Spectra originate from KUG007, RK4002, VS2001 and MK5018. The b- and y-ion series is shown at the top left of each spectrum, and taxonomically informative residues within the peptide sequence are highlighted in pink. A comprehensive list of all identified PSMs and taxonomic assignments is provided in Supplementary Data 3.
North Caucasus
North of the Caucasus mountains, within a geographically and culturally contiguous region that encompasses the piedmont zone, steppe and southern Russian plain, we analysed dietary proteins within the dental calculus proteomes of 27 individuals, including three Eneolithic, 23 Bronze Age and one Early Iron Age individual. Overall, we identified milk proteins in 96% of individuals (n = 26) (Fig. 1c) and observed high levels of milk protein PSMs per individual (mean 47 ± 27; Supplementary Data 3), with milk peptides often being among the most abundant peptides identified in the dental calculus proteomes. Among Eneolithic individuals, two of three were positive for milk proteins. The oldest individual from this region in our study, PG2001 from the piedmont site of Progress 2 and dated to 4338–4074 bc, indicates that dairying has been a feature of the region’s economy since at least the late fifth millennium bc. During the fourth and third millennia bc, we observed a continued reliance on dairying among all analysed Maykop and Steppe Maykop individuals (ca. 3900–2900 bc; n = 7), both in the piedmont and steppe zones as well as in all Yamnaya individuals (ca. 3300–2500 bc; n = 3). Notably, we detected only Ovis milk proteins at Eneolithic, Early Maykop, Late Maykop, Steppe Late Maykop and early Yamnaya sites, suggesting that dairying was a specialized activity focused on sheep during the fourth and fifth millennia bc (Fig. 3a,b). At the start of the early third millennium bc, we identified a broad shift in pastoralist practices towards more diversified dairying based on sheep, goat and cattle milk (Fig. 3c,d). Milk proteins from these three ruminant species were identified among individuals associated with the late Yamnaya (ca. 2850–2500 bc; n = 1), NCC (ca. 2800–2400 bc; n = 4), Catacomb (ca. 2800–2400 bc; n = 1), late NCC (ca. 2200–1650 bc, n = 1) and Lola/post-Catacomb (ca. 2200–1650 bc; n = 6) cultures, with most individuals having consumed the dairy products of two or three different animal milks in the form of sheep and goat milk, sheep and cattle milk or sheep, goat and cattle milk (Fig. 1 and Supplementary Data 3). Finally, during the Early Iron Age (eighth–fifth centuries bc), we observed the incorporation of horse (Equus) milk into the dairy economy (Fig. 3e), with Ovis, Capra, Bos and Equus milk proteins identified in the dental calculus of individual MK5018.

a, During the Eneolithic and initial Bronze Age, dairying focused on sheep in the North Caucasus from 4200 bc onwards. b, Sheep dairying continued during the Early Bronze Age among the Maykop, Steppe Maykop and early Yamnaya. c, After 2800 bc, goat and cattle dairying appeared for the first time in the steppe, and diversified dairy economies of sheep, goats and cattle characterize the late Yamnaya, NCC and Catacomb cultures. d, Diversified dairy economies persisted among the post-Catacomb and Lola cultures, but with an increased focus on sheep and goats as environmental conditions declined. e, During the Late Bronze Age, the North Caucasus steppe was largely depopulated, and ca. 1700 bc a centuries-long hiatus began that corresponded to a period of extreme aridity. Dashed line shows the southern extent of depopulation. f, After 1000 bc, post-hiatus groups repopulated the steppe in the Early Iron Age, resuming sheep, goat and cattle milking and also introducing horse milking. Site colours and animal symbols correspond to those in Fig. 1. All tested individuals in the map extent are shown, including those without evidence of milk protein.
South Caucasus
In the South Caucasus, we analysed dietary proteins within the dental calculus proteomes of nine individuals dating from the Neolithic to Greco-Roman periods and identified milk proteins in half of the analysed individuals (Fig. 1c and Supplementary Data 3). No milk proteins were detected in the earliest individual, MTT001, dated to 5879–5562 bc from the Neolithic site of Mentesh Tepe associated with the Shomutepe–Shulaveri Culture. However, milk proteins were detected from the fourth millennium bc onwards in individuals dating to the Chalcolithic at Alkhantepe (n = 1), the Middle Bronze Age at Qızqala (n = 2), the Iron Age at Göytepe (n = 1) and the Greco-Roman period at Qabala (n = 1). Unlike in the North Caucasus, we did not observe an early focus on sheep dairying; rather, the earliest detected milk protein, identified in individual ALX002 dating to 3776–3651 bc, was assigned to cattle (Bovinae). Overall, we identified cattle (Bovinae), goat (Capra) and sheep (Ovis) milk protein in the South Caucasus but no horse (Equus) milk in any period there (Fig. 3).
Oka–Volga–Don region
In the Oka–Volga–Don region, we analysed dietary proteins within the dental calculus proteomes of seven individuals dating from the Eneolithic through the Middle Bronze Age (Fig. 1c and Supplementary Data 3). Despite excellent protein recovery, no milk proteins were detected in an individual from the Neolithic–Bronze Age site of Ksizovo 6, dating to 5837–5670 bc, nor from individuals associated with the Sredny Stog culture (n = 2) at the Eneolithic–Bronze Age site of Vasilevsky Kordon 27, dating to ca. 3600–3100 bc. Milk proteins were also absent from individual RAV002, dating to 3514–3356 bc, and from two Middle Bronze Age individuals from the Shagara cemetery, dating to 2572–1893 bc. Only an individual at the site of Rovenka tested positive for milk proteins. This individual, RVK001, was associated with a late Catacomb culture site, dating to 2339–2148 bc, and was positive for sheep (Ovis), goat (Capra) and cattle (Bovinae) milk proteins (Fig. 1c).
East Urals region
We analysed two individuals from the East Urals region at the site of Neplyuyevka associated with the Late Bronze Age Srubnaya–Alakul cultural variant and dating to ca. 1900–1600 bc (Fig. 1c). We detected milk proteins for both individuals, identifying sheep (Ovis) and cattle (Bovinae) peptide sequences for NEP008 and non-specific bovid peptide sequences indicating either sheep or cattle for NEP013 (Supplementary Data 3).
Discussion
Eneolithic populations practiced dairy pastoralism
Our results provide robust evidence that sheep dairying was practiced among fifth millennium bc Eneolithic groups in the North Caucasus piedmont and steppe zones. This finding resolves long-standing questions about the antiquity of dairying in the North Caucasus47, as well as the species focus of early dairy herds, and it contributes to a growing body of evidence that dairying was likely introduced with domesticated livestock into the North Caucasus from the south. Recent palaeogenomic studies identified a genetic cline connecting Neolithic populations in eastern Anatolia and the South Caucasus that likely formed as early as 6500 bc32, and continued population interaction into the Chalcolithic and Early Bronze Age periods (5500–3000 bc) suggests that these regions maintained close contact, with animal husbandry focused on pigs and ruminants also spreading via this corridor61,62. Early agropastoralists living in the northern Caucasus foothills associated with the Darkveti–Meshoko Eneolithic culture (ca. 4500–4000 bc) have a clear genetic connection to populations south of the Caucasus exhibiting Anatolian ancestry33, suggesting a trans-Caucasian population expansion.
Although it has been speculated that dairying may have spread to the North Caucasus via these southern connections50, few systematic zooarchaeological studies have been conducted, and the Eneolithic/Chalcolithic layers at the piedmont site of Meshoko Cave, which are among the best studied for the period49, have yielded limited faunal remains, primarily of pigs, sheep, goats and cattle slaughtered at various ages. Subsequent attempts to clarify the agropastoralist economy using stable isotope analysis41,48 have yielded equivocal results as to whether dairying was an Eneolithic or Bronze Age innovation in the North Caucasus. Here, through the identification of taxonomically informative peptides from the milk-specific protein BLG, we confirmed sheep milk consumption by Eneolithic individuals at the sites of Progress 2 and Kurganny 1. Notably, we found that dairy consumption was evident among individuals lacking Anatolian ancestry, such as PG200133, demonstrating that the adoption of dairying by North Caucasian transitional foragers was already underway during the late fifth millennium bc, which precedes Yamnaya expansions by a millennium.
Maykop and Steppe Maykop dairy focused on sheep not cattle
With the start of the fourth millennium bc, we found a continued reliance on dairy pastoralism revealed by ubiquitous evidence of milk consumption among all tested Maykop and Steppe Maykop individuals, further clarifying the high dependence of these groups on animal products41,47. Surprisingly, however, the dairy economy retained an apparent focus on sheep. Although sheep are known to have been important livestock for these groups40,47, cattle feature more prominently at Maykop mortuary sites. They are modelled into gold and silver figurines63, and an emphasis on the power and mobility of cattle is visible in funerary offerings of cattle crania, yokes and nose rings representing oxen teams50. Cattle also appear in bone assemblages at Maykop settlements49. The perishability of the major sheep secondary products of milk and wool, in contrast to the high visibility of cattle-associated material culture and skeletal remains, may have contributed to a biased understanding of the relative importance and roles of these livestock at Early Bronze Age sites64. Our results suggest that cattle were not important dairy livestock during this period and that there was probably a sharp division in livestock use among the Maykop and Steppe Maykop groups41, with sheep being the primary targets of dairying and cattle mainly being used for traction and as a signifier of social identity and status.
Dairy livestock diversified during Middle Bronze Age
A change in dairying strategy to focus on more livestock species coincides with the Yamnaya horizon. Following the Maykop period, mobility expanded ever further with Yamnaya groups, who became the first permanently mobile pastoralists17,44,65,66. Although two early Yamnaya individuals analysed here yielded evidence of only sheep milk product consumption, a more diversified profile comprising sheep, goat and cattle milk was observed for a late Yamnaya individual at the site of Zolotarevka (ZO2002). This trend towards reliance on a broader range of dairy livestock continued and intensified during the Middle Bronze Age, when we observed a general diversification of pastoralist diets to include sheep, goat and cattle milk routinely. Most individuals of the Middle Bronze Age Catacomb, NCC, Late NCC and Lola cultures tested in this study consumed the dairy products of two or three livestock species. Palaeoecological studies have indicated that climate began to shift during the late Yamnaya phase, which also coincided with the first appearance of the Catacomb and NCC groups48. Before this, the climate experienced by the Maykop, Steppe Maykop and early Yamnaya was more favourable67,68 and conducive to regular, short-distance annual mobility47,48. Subsequent aridification encouraged increased mobility, resulting in the exploitation of a wider range of steppe environments beyond the traditional Yamnaya cultural sphere to support livestock herds40,48. The shift to more diverse dairy herds in the North Caucasus also overlaps in time with Yamnaya expansions into southeastern Europe, as well as the parallel rise and expansion of the Corded Ware complex across northeastern and central Europe27, suggesting that these events may be related to broader changes occurring within steppe and forest–steppe pastoralist societies at the time. Our results suggest that an initial diversification of production strategies to include sheep, goat and cattle milk may have functioned as an adaptation to an increasingly turbulent ecological setting, but this subsequently led to overgrazing and lasting damage to pastures due to ground compaction, soil nutrient loss and decreasing plant biomass48,69. At the end of the third millennium bc, coinciding with the emergence of the Lola culture, an intensified drought caused deflation and salinization of the soils in the already dwindling regional watersheds40,69. During the Lola period, water-demanding cattle may have decreased in dairying importance from the preceding Catacomb and NCC periods, as only one of six Lola individuals yielded evidence for cattle milk consumption. After 1700 bc, the steppe and piedmont zones of the Northern Caucasus appear to have been largely depopulated until the ninth or eighth century bc57,70,71, whereas pastoralist groups continued to occupy the high plateaus of the Caucasus Mountains72.
Post-Bronze Age adoption of horse milking
In our dataset, we found no evidence of horse milk consumption until the ninth century bc, when Early Iron Age groups repopulated the North Caucasus steppe and piedmont zones33,41. Horses are well adapted to steppe environments, and recent palaeogenomic research has identified the lower Don–Volga region, possibly as early as the mid-sixth millennium bc, as the domestication centre of the DOM2 horses that characterize present-day lineages43,45. From the Pleistocene until the Bronze Age, horses were hunted on the Pontic–Caspian steppe and have long been symbolically represented in figurines and ritual deposits28,73. Horses are also useful for steppe pastoralists because of their digging (tebenevka) reflex, which allows them to graze through thick snow deposits, thereby opening up winter pasture for ruminants48,74,75. In the North Caucasus, skeletal remains of the ancestors of DOM2 horses are sporadically found in steppe kurgans from the Late Maykop period onwards43, but the role of horses in these pastoralist societies is unclear. The first undisputed evidence of horse traction dates to ca. 2000 bc at the site of Sintashta east of the Urals, where elaborate horse chariot burials have been found in Middle and Late Bronze Age kurgans51,76,77. Earlier Bronze Age wagons, such as those associated with the Late Maykop, Yamnaya and Catacomb cultures, had been pulled by oxen teams50. Herding on horseback, which may have begun ca. 2200 bc with the selection of traits suitable for riding43, would have enabled individual pastoralists to control more livestock at one time and to access pastures across a wider area75. Later, horses became particularly prominent in the archaeological record of Early Iron Age Scythians and Sarmatians, who used horses for cavalry78,79. In addition to traction and riding, horses can also be exploited for milk, which is traditionally fermented to produce an alcoholic beverage in contemporary Eurasian steppe cultures80,81. However, the origin of horse milking is not known. Isotopic evidence from lipids in pottery suggests that Przewalski’s horses, reflecting a separate domestication lineage (DOM1)76, may have been milked as early as the mid-fourth millennium bc at the site of Botai in northern Kazakhstan76,82. It is unclear what, if any, influence early milking at Botai had on the management of DOM2 lineages, the ancestors of modern domestic horses. Currently, the earliest proteomic evidence of horse milk consumption comes from two individuals with problematic dates at the Bronze Age site of Kriviyansky IX in the Lower Don region7 and, later, at the Late Bronze Age site of Uliastai Dood Denzh located in Mongolia, where the dental calculus of an individual dated to ca. 1200 bc with Sintashta-related ancestry yielded evidence of horse milk proteins2,20. Despite an apparent early presence of horse milking at Kriviyansky IX, dating to the third, or possibly fourth, millennium bc, we found no other evidence of horse milking in the North Caucasus region during the Early, Middle or Late Bronze Age. Rather, its late appearance in our dataset suggests that horse milking was a highly limited activity while diverse domestication pathways unfolded, and horses were used for various purposes. Horse milking may have been permanently established in the northern Caucasus only after a later reintroduction by pre-Scythian groups during the first millennium bc. Greek texts, such as The Iliad, later referred to these pre-Scythian steppe nomads as horse milk drinkers83.
Macroregional perspectives on the spread of dairying
The Pontic–Caspian steppe has long been recognized as a major centre for pastoralist innovation. Here we show that dairying was an early and enduring feature of the pastoralist economy not only in the Northern Caucasus, but also in the South Caucasus. In our dataset, we observed the earliest evidence of milk consumption in the South Caucasus at Alkhantepe, a Late Chalcolithic site with Leilatepe ceramics84,85. The contemporaneous Leilatepe and Early Maykop cultures share many features39,86, but we found that the agropastoralists at Alkhantepe were milking cattle, whereas we observed only sheep milking at Early and Late Maykop sites in the north. Sheep and cattle have different ecological needs, and, in particular, sheep require less water and can survive harsher winters than cattle. As such, environmental factors may have played a role in influencing the selection of dairy livestock in these two regions. During the third millennium bc, it is known that the economic importance of pastoralism increased in the South Caucasus, especially during the Kura–Araxes period56,87, but we did not have corresponding samples to examine this. Although steppe cultural elements, such as kurgans (burial mounds), had been present in the South Caucasus since the Late Chalcolithic88, kurgans greatly increased during the Middle Bronze Age89, and we next observed dairy product consumption at the Middle Bronze Age fortified agropastoral site of Qızqala, with ruminant dairy proteins present in both individuals analysed for this study. Although Middle Bronze Age cultures in both the North and South Caucasus largely became fully mobile to support their herds90, the inhabitants of Qızqala relied on a more flexible subsistence strategy that included both settlement occupation and seasonal movement of livestock89,91. Our results show a reliance on dairy technology for subsistence for these mobile pastoralists. Next, we found evidence of sheep milk consumption by one individual from an intrusive Late Bronze/Early Iron Age burial associated with the Khojaly–Gadabay culture at the Neolithic site of Göytepe. This is the earliest unequivocal evidence of sheep milking in our South Caucasus dataset. Later, during the Greco-Roman era, we observe evidence of sheep, goat and cattle milk at Qabala, a site associated with complex and intensive agriculture as well as with local herding.
Despite cultural interaction with adjacent communities of the Pontic–Caspian steppe, communities in the Oka–Volga–Don forested regions maintained economies based on hunting, gathering and fishing that were particularly suited to local ecozones. Stable isotope studies suggest that this was the prevailing economic strategy until the end of the third millennium bc during the Middle Bronze Age48,92,93 when Oka–Volga–Don communities transitioned to agropastoralist subsistence44. Although populations further to the east, between the Volga River and the Ural Mountains, practiced ruminant dairying from ca. 3000 bc onwards7, the near-complete lack of evidence for ruminant milk consumption from the seven individuals representing the Oka–Volga–Don region in our study is consistent with a late introduction of ruminant dairying west of the Volga, despite the fact that domesticated animals were introduced in small quantities during the late fourth millennium bc. Here, only one Catacomb-associated individual with cultural links to the steppe zone, recovered from the site of Rovenka, yielded ruminant milk proteins, which were sourced from sheep, goats and cattle.
In parallel to the expansion of pastoralism to the forest–steppe zone, contact and admixture with late farming groups in eastern Europe, such as Cucuteni–Trypillia and Globular Amphora, resulted in a mixed form of agropastoralism with heavy reliance on pastoralism94, followed by a subsequent eastward expansion of the Corded Ware complex during the third and early second millennia bc, which is also attested by archaeogenetic data22,95. This sphere of influence includes Fatyanovo/Balanovo and subsequent Abashevo, Sintashta, Andronovo, and Srubnaya groups94, and individuals associated with these cultures share very similar genetic profiles. We analysed two individuals linked to the Srubnaya culture at the Middle to Late Bronze Age site of Neplyuyevka in the region east of the Ural Mountains and identified evidence of ruminant milk consumption. Future work combining palaeogenomic and palaeodietary research could help to better clarify the relationships between these populations and the nature and spatio-temporal patterning of dairy technologies in this region.
Conclusion
Proteomic analysis of human dental calculus has revealed a dynamic trajectory of dairy pastoralism in the North Caucasus steppe and adjacent regions from the Eneolithic to the Greco-Roman periods. Dairying was integral for the spread of animal husbandry by groups crossing the Caucasus mountains from south to north during the Eneolithic, and it was quickly adopted and further developed into an effective and sustainable technology—dairy pastoralism—by neighbouring steppe communities. This innovation forms the basis of the Eurasian steppe lifestyle that continues until today. Initial pastoralist strategies focused on sheep dairying and cattle traction, whereas fully mobile pastoralism arose for the first time during the Yamnaya period. Deteriorating climatic conditions challenged steppe herders during the Middle and Late Bronze Ages, who responded by diversifying their set of dairying livestock and expanding their herding range, until the steppe was ultimately abandoned in the mid-second millennium bc. Later, following a centuries-long hiatus, the steppe was repopulated by Early Iron Age pastoralists who practised horse milking. The turbulent third millennium bc, during which vast stretches of Eurasia experienced social and demographic upheaval, is now coming into sharper focus. Climatic pressures and the needs of dairy herds altered how pastoralists used the North Caucasus steppe and may have contributed to the heightened mobility of third-millennium-bc steppe herders, whose descendants spread across Eurasia within the span of only a few centuries. Future research on the genomes of ancient dairying livestock and additional dental calculus proteomes from adjacent steppe populations north of the Black Sea and east of the Urals will help to further clarify the origins and dispersals of dairying breeds and practices that promoted the lasting cultural and subsistence traditions that reshaped the Eurasian steppe zone and profoundly transformed the Bronze Age Eurasian world.
Methods
Sampling
Dental calculus sampling was performed on site at archaeological institutions and museums and in a dedicated ancient biomolecules laboratory at the Max Planck Institute for the Science of Human History (MPI-SHH). Disposable nitrile gloves were worn during collection, and calculus was sampled using dental curettes that were replaced or cleaned with isopropanol between samples. Calculus was collected onto weighing paper and stored in microcentrifuge tubes. Samples were further analysed at the MPI-SHH ancient proteomics laboratory, where they were weighed and subsampled before protein extraction. Approximately 5–13 mg of dental calculus was used for each protein analysis.
Radiocarbon dating
A total of 24 new radiocarbon dates were obtained by accelerator mass spectrometry of bone and tooth material at: the Curt-Engelhorn-Zentrum Archäometrie in Mannheim, Germany; the Finnish Museum of Natural History (Hela) in Helsinki, Finland; the Oxford Radiocarbon Accelerator Unit in Oxford, United Kingdom; and the Russian Academy of Sciences in Moscow, Russia. Uncalibrated dates were successfully obtained for all but one tested sample (Supplementary Data 1). An additional 21 previously published radiocarbon dates for individuals in this study were also compiled and analysed, making the total number of directly dated individuals in this study 38 (45 total dates). Dates were calibrated using OxCal v.4.496 with the IntCal20 atmospheric curve97.
Liquid chromatography–tandem mass spectrometry and data analysis
Archaeological dental calculus samples from 45 individuals and 5 extraction non-template controls were processed using a filter-aided sample-preparation protocol, modified for ancient proteins (https://doi.org/10.17504/protocols.io.7vwhn7e). In brief, dental calculus was demineralized in 0.5 M EDTA, and proteins were solubilized and reduced using SDS lysis buffer (4% SDS, 0.1 M DTT, 0.1 M Tris HCl). Buffer exchange in 8 M urea and total protein isolation were performed using a Microcon 30 kDa centrifugal filter unit with an Ultracel-30 membrane (Millipore), followed by alkylation using iodoacetamide. Following buffer replacement with triethylammonium bicarbonate (TEAB; 0.05 M), the proteins were digested overnight with sequencing-grade modified trypsin (Promega) at 37 °C. Peptides were recovered by centrifugation in TEAB and acidified with trifluoroacetic acid to pH <3 and desalted using C18 stage tips (Pierce). Peptides were analysed by liquid chromatography–tandem mass spectrometry using a Q-Exactive mass spectrometer (Thermo Fisher Scientific) coupled to an ACQUITY UPLC M-Class system (Waters AG) at the Functional Genomics Center Zurich of the University/ETH Zurich. Spectra were acquired from 300–1,700 m/z with an automatic gain control target of 3 × 106, a resolution of 70,000 (at 200 m/z) and a maximum injection time of 110 ms. The quadrupole isolated precursor ions with a 2.0 m/z window, a 5 × 104 automated gain control value and a maximum fill time of 110 ms. Twelve of the most intense precursor ions for each MS1 scan were fragmented via high collision dissociation with a normalized collision energy of 25, scanned with a resolution of 35,000 (at 200 m/z) and a fixed first mass of 200 m/z. An intensity threshold of 9.1 × 103 was applied for MS2 selection, and singly charged ions were excluded. Filter criteria for MS2 selection were an intensity threshold of 9.1 × 103, and unassigned, singly charged ions were excluded. Selected precursor ions were put onto a dynamic exclusion list for 30 s. For liquid chromatography, the solvent composition at the two channels was 0.1% formic acid in water for channel A and 0.1% formic acid in acetonitrile for channel B. Next, 4 µl of each peptide sample was loaded onto a trap column (Symmetry C18, 100 Å, 5 µm, 180 µm × 20 mm; Waters AG) with a flow rate of 15 µl min−1 of 99% solvent A for 60 s at room temperature. Peptides eluting from the trap column were refocused and separated on a C18 column (HSS T3 C18, 100 Å, 1.8 µm, 75 µm × 250 mm; Waters AG). The column temperature was 50 °C. Peptides were separated over 73 min with the following gradient: 8–22% solvent B in 49 min, 22–32% solvent B in 11 min and 32–95% solvent B in 5 min. The column was cleaned with 95% solvent B for 5 min after the separation and re-equilibrated at loading condition for 8 min before initializing the next run. Potential contamination was monitored using extraction blanks.
Tandem mass spectra were converted to Mascot generic files by MSConvert version 3.0.11781 using the 100 most intense peaks in each spectra. All tandem mass spectrometry samples were analysed using Mascot (Matrix Science, version 2.6.0). Mascot was set up to search the SwissProt Release 2019_08 database (560,823 entries) assuming the digestion enzyme trypsin, with automatic decoy option. Mascot was searched with a fragment ion mass tolerance of 0.050 Da and a parent ion tolerance of 10.0 ppm. The number of missed cleavages was specified as one. Carbamidomethyl of cysteine was specified in Mascot as a fixed modification. Deamidation of asparagine and glutamine and oxidation of methionine and proline were specified in Mascot as variable modifications.
Scaffold version 4.9.0 (Proteome Software Inc.) was used to validate protein and peptide identifications for each sample. Peptide identifications were accepted if they could be established at greater than a 90% probability by the PeptideProphet algorithm. Protein identifications were accepted if they could be established at a greater than 95% probability and contained at least two unique peptides. Probabilities for proteins were assigned using the ProteinProphet algorithm98. Proteins that contained similar peptides that could not be differentiated based on tandem mass spectrometry analysis alone were grouped to satisfy the principles of parsimony, and proteins that shared significant peptide evidence were grouped into clusters. Peptide identifications were accepted if they could be established at a greater than 90% probability using the PeptideProphet algorithm99 with Scaffold delta-mass correction. Individual protein and peptide false discovery rates are listed in Supplementary Data 3.
Reporting Summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
Raw data files are available through the ProteomeXchange Consortium via the PRIDE partner repository under accession PDX027728. Source data are provided with this paper.
References
Bleasdale, M. et al. Ancient proteins provide evidence of dairy consumption in eastern Africa. Nat. Commun. 12, 632 (2021).
Wilkin, S. et al. Dairy pastoralism sustained eastern Eurasian steppe populations for 5,000 years. Nat. Ecol. Evol. 4, 346–355 (2020).
Evershed, R. P. et al. Earliest date for milk use in the Near East and southeastern Europe linked to cattle herding. Nature 455, 528–531 (2008).
Salque, M. et al. Earliest evidence for cheese making in the sixth millennium bc in northern Europe. Nature 493, 522–525 (2013).
Dunne, J. et al. First dairying in green Saharan Africa in the fifth millennium bc. Nature 486, 390–394 (2012).
Hendy, J. et al. Ancient proteins from ceramic vessels at Çatalhöyük West reveal the hidden cuisine of early farmers. Nat. Commun. 9, 4064 (2018).
Wilkin, S. et al. Dairying enabled Early Bronze Age Yamnaya steppe expansions. Nature 598, 629–633 (2021).
Jeong, C. et al. Bronze Age population dynamics and the rise of dairy pastoralism on the eastern Eurasian steppe. Proc. Natl Acad. Sci. USA 115, E11248–E11255 (2018).
Honeychurch, W. & Makarewicz, C. A. The archaeology of pastoral nomadism. Annu. Rev. Anthropol. 45, 341–359 (2016).
Weaver, C., Wijesinha-Bettoni, R. McMahon, D. & Spence, L. n Milk and Dairy Products in Human Nutrition (eds Muehlhoff, E., Bennett, A. & Macmahon, D.) 103–206 (Food and Agriculture Organization of the United Nations, 2013).
Tishkoff, S. A. et al. Convergent adaptation of human lactase persistence in Africa and Europe. Nat. Genet. 39, 31–40 (2007).
Ségurel, L. & Bon, C. On the evolution of lactase persistence in humans. Annu. Rev. Genomics Hum. Genet. 18, 297–319 (2017).
Metheny, K. B. & Beaudry, M. C. Archaeology of Food: An Encyclopedia (Rowman & Littlefield, 2015).
Wijesinha-Bettoni, R. & Burlingame, B. in Milk and Dairy Products in Human Nutrition (eds Muehlhoff, E., Bennett, A. & Macmahon, D.) 41–102 (Food and Agriculture Organization of the United Nations, 2013).
Bogaard, A. The archaeology of food surplus. World Archaeol. 49, 1–7 (2017).
Dunne, J., di Lernia, S., Chłodnicki, M., Kherbouche, F. & Evershed, R. P. Timing and pace of dairying inception and animal husbandry practices across Holocene North Africa. Quat. Int. 471, 147–159 (2018).
Frachetti, M. D. Multiregional emergence of mobile pastoralism and nonuniform institutional complexity across Eurasia. Curr. Anthropol. 53, 2–38 (2012).
Segurel, L. et al. Why and when was lactase persistence selected for? Insights from Central Asian herders and ancient DNA. PLoS Biol. 18, e3000742 (2020).
Taylor, W. T. T. et al. Evidence for early dispersal of domestic sheep into Central Asia. Nat. Hum. Behav. 5, 1169–1179 (2021).
Jeong, C. et al. A Dynamic 6,000-year genetic history of Eurasia’s Eastern Steppe. Cell 183, 890–904.e29 (2020).
Allentoft, M. E. et al. Population genomics of Bronze Age Eurasia. Nature 522, 167–172 (2015).
Mathieson, I. et al. Genome-wide patterns of selection in 230 ancient Eurasians. Nature 528, 499–503 (2015).
Kovalev, A. A. & Erdenebaatar, D. in Afanas’yevskiy sbornik [in Russian] (eds Stepanova, N. F. & Polyakov, A. V.) 91–108 (Azbuka, 2010).
Haak, W. et al. Massive migration from the steppe was a source for Indo-European languages in Europe. Nature 522, 207–211 (2015).
Olalde, I. et al. The Beaker phenomenon and the genomic transformation of northwest Europe. Nature 555, 190–196 (2018).
Sjögren, K.-G., Price, T. D. & Kristiansen, K. Diet and mobility in the corded ware of Central Europe. PLoS ONE 11, e0155083 (2016).
Papac, L. et al. Dynamic changes in genomic and social structures in third millennium bc central Europe. Sci. Adv. 7, eabi6941 (2021).
Benecke, N. Archaeozoological studies on the transition from the Mesolithic to the Neolithic in the North Pontic region. Anthropozoologica 25, 631–641 (1997).
Meshveliani, T. et al. Mesolithic hunters at Kotias Klde, Western Georgia: preliminary results. Paléorient 33, 47–58 (2007).
Vybornov, A., Kosintesev, P. & Kulkova, M. The origin of farming in the Lower Volga region. Documenta Praehistorica 62, 67–75 (2015).
Nishiaki, Y. et al. The Mesolithic-Neolithic interface in the Southern Caucasus: 2016–2017 excavations at Damjili Cave, West Azerbaijan. Archaeol. Res. Asia 19, 100140 (2019).
Skourtanioti, E. et al. Genomic history of Neolithic to Bronze Age Anatolia, Northern Levant, and Southern Caucasus. Cell 181, 1158–1175.e28 (2020).
Wang, C.-C. et al. Ancient human genome-wide data from a 3000-year interval in the Caucasus corresponds with eco-geographic regions. Nat. Commun. 10, 590 (2019).
Gorelik, A. F., Cybrij, A. & Cybrij, V. Zu kaukasischen und vorderasiatischen Einflüssen bei der Neolithisierung im unteren Donbecken. Eurasia Antiq. 20, 143–170 (2017).
Reinhold, A., Belinskiy, A. B. & Korobov, D. S. in Archaeology of Mountain Landscapes: Interdisciplinary Research Strategies of Agro-Pastoralism in Upland Regions (ed Garcia-Molsosa, A.) Chapter 9 (State University of New York Press, 2022).
Rassamakin, Y. in Late Prehistoric Exploitation of the Eurasian Steppe (eds Levine, M. et al.) 59–182 (Univ. Cambridge, 1999).
Trifonov, V. in Adaptation of Paleolithic–Eneolithic Cultures to Changes in the Natural Environment in the North-West Caucasus [in Russian] (ed Trifonov, V.) 84–93 (Institute for the History of Material Culture RAS, 2009).
Kohl, P. L. & Trifonov, V. in The Cambridge World Prehistory (eds Renfrew, C. & Bahn, P.) 1571–595 (Cambridge Univ. Press, 2014).
Lyonnet, B. in Les Cultures du Caucase (VIe- IIIe Millénaires avant Notre ère): Leurs Relations avec le Proche-Orient (ed Lyonnet, B.) 133–161 (CNRS Éditions, 2007).
Shishlina, N. in Counterpoint: Essays in Archaeology and Heritage Studies in Honour of Professor Kristian Kristiansen (eds Bergerbrant, S. & Sabatini, S.) 53–60 (BAR International, 2013).
Knipper, C. et al. Diet and subsistence in Bronze Age pastoral communities from the southern Russian steppes and the North Caucasus. PLoS ONE 15, e0239861 (2020).
Benecke, N. in The Oxford Handbook of Zooarchaeology (eds Albarella, U. et al.) 319–332 (Oxford Univ. Press, 2017).
Librado, P. et al. The origins and spread of domestic horses from the Western Eurasian steppes. Nature 598, 634–640 (2021).
Anthony, D. W. The Horse, the Wheel, and Language: How Bronze-Age Riders from the Eurasian Steppes Shaped the Modern World. (Princeton Univ. Press, 2010).
Guimaraes, S. et al. Ancient DNA shows domestic horses were introduced in the southern Caucasus and Anatolia during the Bronze Age. Sci. Adv. 6, eabb0030 (2020).
Staubwasser, M. & Weiss, H. Holocene climate and cultural evolution in late prehistoric–early historic West Asia. Quat. Res. 66, 372–387 (2006).
Shishlina, N. Reconstruction of the Bronze Age of the Caspian Steppes: Life Styles and Life Ways of Pastoral Nomads (Archaeopress, 2008).
Shishlina, N., Sevastyanov, V. & Kuznetsova, O. Seasonal practices of prehistoric pastoralists from the south of the Russian plain based on the isotope data of modern and archaeological animal bones and plants. J. Archaeol. Sci. Rep. 21, 1247–1258 (2017).
Hambleton, E. & Maltby, M. The Animal Bones from Excavations in Meshoko Cave in the Northern Caucasus (Bournemouth Univ. & St Petersburg State Univ., 2016); http://eprints.bournemouth.ac.uk/24860/1/MeshokoCaveReportJuly2016.pdf
Reinhold, S. et al. in Appropriating Innovations: Entangled Knowledge in Eurasia, 5000‒1500 BC (eds Stockhammer, P. & Maran, J.) 78–97 (Oxbow Books, 2017).
Chechushkov, I. V. & Epimakhov, A. V. Eurasian steppe chariots and social complexity during the Bronze Age. J. World Prehistory 31, 435–483 (2018).
Shishlina, N., Orfinskaya, O. V. & Hommel, P. Bronze Age wool textile of the Northern Eurasia: new radiocarbon data. Nanotechnol. Russ. 15, 629–638 (2020).
Ethier, J. et al. Earliest expansion of animal husbandry beyond the Mediterranean zone in the sixth millennium bc. Sci. Rep. 7, 7146 (2017).
Stojanovski, D. et al. Living off the land: terrestrial-based diet and dairying in the farming communities of the Neolithic Balkans. PLoS ONE 15, e0237608 (2020).
McClure, S. B. et al. Fatty acid specific δ13C values reveal earliest Mediterranean cheese production 7,200 years ago. PLoS ONE 13, e0202807 (2018).
Herrscher, E. et al. Dietary inferences through stable isotope analysis at the Neolithic and Bronze Age in the southern Caucasus (sixth to first millenium bc, Azerbaijan): from environmental adaptation to social impacts. Am. J. Phys. Anthropol. 167, 856–875 (2018).
Reinhold, S., Korobov, D. & Belinskij, A. B. Landschaftsarchäologie im Nordkaukasus. Studien zu einer Neu Entdeckten bronzezeitlichen Kulturlandschaft im Hochgebirge des Nordkaukasus Vol. 38 (Habelt-Verlag, 2017).
Chazin, H. The Politics of Pasture: The Organization of Pastoral Practices and Political Authority in the Late Bronze Age in the South Caucasus (Univ. Chicago, 2016).
Uerpmann, M. & Uerpmann, H. P. Bronze and Iron Age animal economy at Didi-gora and Tqisbolo-gora (Kakhetia, Georgia). Archäol. Mittl. aus Iran und Turan 40, 169–264 (2008).
Warinner, C. et al. Direct evidence of milk consumption from ancient human dental calculus. Sci. Rep. 4, 7104 (2014).
Ricci, A., D’Anna, M. B., Lawrence, D., Helwing, B. & Aliyev, T. Human mobility and early sedentism: the Late Neolithic landscape of southern Azerbaijan. Antiquity 92, 1445–1461 (2018).
Berthon, R. Past, current and future contribution of zooarchaeology to the knowledge of the Neolithic and Chalcolithic cultures in South Caucasus. Stud. Cauc. Archaeol. 2, 4–30 (2014).
Piotrovskij, J. J. in Der Kaukasus zwischen Osteuropa und Vorderem Orient in der Bronze und Eisenzeit (eds Kašuba, M. T. et al.) Vol. 19 (Dietrich Reimer Verlag, 2020).
Korenevsky, S. N. The Most Ancient Farmers and Stockbreeders of the Pre-Caucasus Region: The Maikop–Novosvobodnoe Community, Problems of Internal Typology and Symbolism of Funerary Practices [in Russian] (Nauka, 2004).
Merpert, N. Y. A. The most ancient stockbreeders of the Volga-Ural interfluve [in Russian] (Nauke, 1974).
Kaiser, E. Das Dritte Jahrtausend im Osteuropäischen Steppenraum: Kulturhistorische Studien zu Prähistorischer Subsistenzwirtschaft und Interaktion mit Benachbarten Räumen. Vol. 37 (Freie Univ. Berlin, 2019).
Connor, S. E. & Kvavadze, E. V. Modelling late Quaternary changes in plant distribution, vegetation and climate using pollen data from Georgia, Caucasus. J. Biogeogr. 36, 529–545 (2009).
Davis, B. A. S., Brewer, S., Stevenson, A. C. & Guiot, J. The temperature of Europe during the Holocene reconstructed from pollen data. Quat. Sci. Rev. 22, 1701–1716 (2003).
Borisov, A. V., Demkina, T. S. & Demkin, V. A. Paleoenvironments and the Climate of Yerengei in the Bronze Age, IV-II Millennium bc [in Russian] (Nauka, 2006).
Malashev, V. Y. in Traditions of the Middle Sarmatian Culture in Burial Mounds of the Caspian Dagestan in the Second Half of the Second–First Half of the Fifth Century ad [in Russian] Vol. 35 85–97 (Bulletin of Volga State Univ., 2015).
Erlikh, V.R. in Achaemenid Impact in the Black Sea: Communication of Powers (eds Nieling, J. & Rehm, E.) 47–66 (Aarhus Univ. Press, 2010).
Reinhold, S., Belinskij, A. B. & Atabiev, B. H. in Der Kaukasus zwischen Osteuropa und Vorderem Orient in der Bronze und Eisenzeit (eds Kašuba, M. T. et al.) Vol. 19 (Dietrich Reimer Verlag, 2020).
Anthony, D. W. & Brown, D. R. Eneolithic horse exploitation in the Eurasian steppes: diet, ritual and riding. Antiquity 74, 75–86 (2000).
Taylor, W. T. T. et al. Early pastoral economies and herding transitions in Eastern Eurasia. Sci. Rep. 10, 1001 (2020).
Khazanov, A. M. Nomads and the Outside World (The Univ. of Wisconsin Press, 1994).
Gaunitz, C. et al. Ancient genomes revisit the ancestry of domestic and Przewalski’s horses. Science 360, 111–114 (2018).
Taylor, W. T. T. & Barrón-Ortiz, C. I. Rethinking the evidence for early horse domestication at Botai. Sci. Rep. 11, 7440 (2021).
Perevalov, S. M. The Sarmatian lance and the Sarmatian horse-riding posture. Anthropol. Archeol. Eurasia 40, 7–21 (2002).
Spasskaya, N. N., Kantorovich, A. R. & Maslov, V. E. in Steppes of Northern Eurasia: Proceedings of the IX International Symposium [in Russian] Vol. 9 764–768 (Institute of the Steppe of the Ural Branch of the Russian Academy of Sciences, 2021).
McGuire, G. Cultural histories of kumiss: tuberculosis, heritage and national health in post-Soviet Kazakhstan. Cent. Asian Surv. 36, 493–510 (2017).
Bat-Oyun, T., Erdenetsetseg, B., Shinoda, M., Ozaki, T. & Morinaga, Y. Who is making airag (fermented mare’s milk)? A nationwide survey of traditional food in mongolia. Nomad People 19, 7–29 (2015).
Outram, A. K. et al. The earliest horse harnessing and milking. Science 323, 1332–1335 (2009).
Butler, S. et al. The Iliad of Homer: Rendered into English Prose for the Use of Those who Cannot Read the Original (Longman’s, Green and Co., 1898).
Akhundov, T. I. Monuments of the Mugan steppe and preconditions for the settlement of early rarmers in the South Caucasus in the Neolithic–Eneolithic [in Russian]. Stratum Plus 2, 219–236 (2011).
Poulmarc’h, M. & Le Mort, F. Diversification of the funerary practices in the Southern Caucasus from the Neolithic to the Chalcolithic. Quat. Int. 395, 184–193 (2016).
Museyibli, N. Potter’s marks on Leilatepe culture pottery: Eastern Anatolian Chalcolithic traditions in the Caucasus. Mediterr. Archaeol. Archaeom. 16, 283–294 (2016).
Decaix, A. et al. in The Iranian Plateau during the Bronze Age: Development of Urbanisation, Production and Trade (eds Meyer, J.-W. et al.) 89–100 (MOM Éditions, 2019).
Lyonnet, B. et al. Late Chalcolithic Kurgans in Transcaucasia: the cemetery of Soyuq Bulaq (Azerbaijan). Archäol. Mittl. aus Iran und Turan 40, 27–44 (2008).
Nugent, S. E. Pastoralism and emergent complex settlement in the Middle Bronze Age, Azerbaijan: isotopic analyses of mobility strategies in transformation. Am. J. Phys. Anthropol. 171, 120–141 (2020).
Sagona, A. The Archaeology of the Caucasus: From Earliest Settlements to the Iron Age. (Cambridge Univ. Press, 2018).
Lau, H., Proctor, L., Gopnik, H. & Bakhshaliyev, V. Agropastoralism in middle Bronze through early Iron Age Naxçıvan: zooarchaeological and paleoethnobotanical data from Qızqala. J. Archaeol. Sci. Rep. 33, 102535 (2020).
Shishlina, N., Kaverzneva, E. & Fernandes, R. Subsistence strategies of Meshchera lowlands populations during the Eneolithic period — the Bronze Age: results from a multidisciplinary approach. J. Archaeol. Sci. Rep. 10, 74–81 (2016).
Schulting, R. J. & Richards, M. P. in A Bronze Age Landscape in the Russian Steppes: The Samara Valley Project (ed Anthony D.W. et al.) 127–149 (UCLA Cotsen Institute of Archaeology Press, 2016).
Nordqvist, K. & Heyd, V. The forgotten child of the wider Corded Ware family: Russian Fatyanovo culture in context. Proc. Prehist. Soc. 86, 65–94 (2020).
Narasimhan, V. M. et al. The formation of human populations in South and Central Asia. Science 365, eaat7487 (2019).
Ramsey, C. B. Bayesian analysis of radiocarbon dates. Radiocarbon 51, 337–360 (2009).
Reimer, P. J. et al. The IntCal20 northern hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 62, 725–757 (2020).
Nesvizhskii, A. I., Keller, A., Kolker, E. & Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 75, 4646–4658 (2003).
Keller, A., Nesvizhskii, A. I., Kolker, E. & Aebersold, R. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 74, 5383–5392 (2002).
Acknowledgements
This research was funded by the Max Planck Society, the European Research Council under the European Union’s Horizon 2020 research and innovation programme (grant agreement numbers 771234-PALEoRIDER, W.H.; 834616-ARCHCAUCASUS, S.H.; 678901-FoodTransforms, A.S.; and 804884-DAIRYCULTURES, C.W.), the ERA.Net RUS Plus initiative (S&T-277-BIOARCCAUCASUS, S.H., A.Bu.), and the Russian Science Foundation (RSF grant 21-18-00026, N.S.).
Funding
Open access funding provided by Max Planck Society.
Author information
Authors and Affiliations
Contributions
C.W., W.H., S.H., E.H., and J.K. designed the study. S.R., S.H., A.R.K., V.E.M., A. Belinsky, N.B., A. Buzhilova, F.G., B.L., P.G., B.J., J.E., E.I., E.H., S.N., K.M., P.O., K.N., N.S., E.K., A.I.K., A.A. Kalmykov, A.A. Khokhlov, R.V.S., R.K., S.V.S., E.S. and M.K. provided materials and resources. A.S. and R.H. performed laboratory experiments. A.S. analysed the data. C.W., A.S., S.R., T.H., S.H. and W.H. assisted with data interpretation. A.S., S.R., T.H., W.H. and C.W. wrote the manuscript with contributions from all co-authors.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Ecology & Evolution thanks Yimin Yang, Alan Outram, Rosa Jersie-Christensen and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information
Supplementary Discussion.
Supplementary Data 1
Sample information and radiocarbon dates.
Supplementary Data 2
Protein modifications.
Supplementary Data 3
Identified milk peptides in summary format and by individual.
Source data
Source Data Fig. 1
Statistical source data.
Source Data Fig. 2
Statistical source data.
Source Data Fig. 3
Statistical source data.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Scott, A., Reinhold, S., Hermes, T. et al. Emergence and intensification of dairying in the Caucasus and Eurasian steppes. Nat Ecol Evol 6, 813–822 (2022). https://doi.org/10.1038/s41559-022-01701-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41559-022-01701-6
This article is cited by
-
Widespread horse-based mobility arose around 2200 bce in Eurasia
Nature (2024)
-
Permafrost preservation reveals proteomic evidence for yak milk consumption in the 13th century
Communications Biology (2023)
-
Prehistoric population expansion in Central Asia promoted by the Altai Holocene Climatic Optimum
Nature Communications (2023)
-
Early contact between late farming and pastoralist societies in southeastern Europe
Nature (2023)
