
Abstract
Ketogenic diet (KD) alleviates refractory epilepsy and reduces seizures in children. However, the metabolic/cell biologic mechanisms by which the KD exerts its antiepileptic efficacy remain elusive. Herein, we report that KD-produced β-hydroxybutyric acid (BHB) augments brain gamma-aminobutyric acid (GABA) and the GABA/glutamate ratio to inhibit epilepsy. The KD ameliorated pentetrazol-induced epilepsy in mice. Mechanistically, KD-produced BHB, but not other ketone bodies, inhibited HDAC1/HDAC2, increased H3K27 acetylation, and transcriptionally upregulated SIRT4 and glutamate decarboxylase 1 (GAD1). BHB-induced SIRT4 de-carbamylated and inactivated glutamate dehydrogenase to preserve glutamate for GABA synthesis, and GAD1 upregulation increased mouse brain GABA/glutamate ratio to inhibit neuron excitation. BHB administration in mice inhibited epilepsy induced by pentetrazol. BHB-mediated relief of epilepsy required high GABA level and GABA/glutamate ratio. These results identified BHB as the major antiepileptic metabolite of the KD and suggested that BHB may serve as an alternative and less toxic antiepileptic agent than KD.
Similar content being viewed by others

Introduction
The ketogenic diet (KD) switches cell energy metabolism from carbohydrates to fatty acid and amino acid oxidation1, from which ketone bodies, such as β-hydroxybutyrate (BHB), acetone, and acetoacetate (AcAc), are generated2. A consensus is yet to be reached regarding whether physiologic effects of the KD, including weight loss3, decreasing blood sugar via normalizing beta cell function4, and cholesterol level reduction5, are due to altered energy metabolism or ketone body production6,7. One of the recognized efficacies of the KD is its beneficial neuronal effects. The KD has been reported to help treat brain disorders such as Parkinson’s disease8, Alzheimer’s disease9, multiple sclerosis10,11, sleep disorders12, autism13, and brain cancer14. Since its first clinical practice in epilepsy treatment in 192115, KD has been shown to be effective in treating children with refractory epilepsy16,17, in which onset often occurs in childhood and older adulthood18.
Epilepsy is a common neurological condition which affects 0.8%–1% of the human population, and ~30% of patients with epilepsy have drug-resistant epilepsy. Epileptic seizures are mostly due to a relative imbalance of excitatory and inhibitory neurotransmitters, including glutamate and gamma-aminobutyric acid (GABA)19, that leads to abnormal and excessively synchronized firing of neurons in the brain20. In the central nervous system, GABA is primarily produced by glutamate decarboxylase 1 (GAD1), which decarboxylates glutamate within GABAergic axon terminals21. The interictal epileptic activity in the human temporal lobe is critically dependent on the depolarizing action of GABA22. Disturbance of central glutamate and GABA homeostasis is consistently associated with neurological disorders, such as schizophrenia and epilepsy23,24. The proposed antiepileptic mechanisms include inhibiting the presynaptic release of glutamate by directly competing with Cl– for allosteric activation of vesicular glutamate transporters and increasing levels of GABA in cerebrospinal fluid25. BHB can be produced in astrocytes from oxidation of fatty acids or catabolism of amino acids, and is metabolized in the mitochondria of all brain cell types26. The open probability of KATP channels in the hippocampus in vitro is enhanced in the presence of BHB27.
Glutamate receives amines from most amino acids for deamination and is the most abundant excitatory neurotransmitter in the vertebrate nervous system28. Glutamate can be converted to α-ketoglutarate (α-KG) to enter the tricarboxylic acid cycle29 via glutamate dehydrogenase (GDH) or to GABA via GAD1. It has been suggested that SIRT4 represses enzymatic activity and limits the catabolism of glutamate and glutamine by ADP-ribosylation of GDH30. GDH-produced ammonia is converted to carbamoyl phosphate (CP), which spontaneously forms protein lysine carbamylation (CP-K) to activate mitochondrial proteins, including ornithine transcarbamylase (OTC) of the urea cycle; CP-K effects on OTC are reversed via mitochondrial sirtuin SIRT431. SIRT4 regulates GDH activity and GABA/glutamate ratio that determines neuronal excitation, it is possible that CP-K and SIRT4 can regulate glutamate and GABA homeostasis, and consequently determine GABA/glutamate ratio in epilepsy control.
Results
KD increased the GABA, glutamate, and GABA/glutamate ratio in the brains of mice
To investigate the seizure-preventing effects of the KD, 8-week-old male C57/BL6 mice were fed with either a KD or normal diet (ND) for 12 weeks before they were intraperitoneally injected with pentetrazol (PTZ), which induces acute epilepsy in mice32. Female mice were not used in this study to exclude the effects of hormone changes during puberty and hormone fluctuations over estrous cycle33. The development of both clonic and tonic-clonic seizures began with stereotypic behaviors, such as Straub tail, myoclonic jerks and hindlimb flexions, which progressed to forelimb clonus and head bobbing and ended with relaxed muscle tone. And an isoelectric period34 was monitored. The severity of epilepsy was calculated according to the Racine scale (Fig. 1a)35.

a Schematic diagram of the timeline of dietary exposure and PTZ treatment of mice and the Racine scale used to assess the intensity of epilepsy in mice. b–e PTZ-induced epilepsy in mice. KD- and ND-fed mice were intraperitoneally administered with a shot of PTZ before being monitored for epilepsy symptoms that were staged by the Racine scale. Latency time (b), the highest level (c), duration of (d) and frequency of seizures (e) were monitored. Data are means ± SD, n = 10 mice in each treatment. f The KD weakened neuronal activity in mice. The representative EEG recordings of WT mice fed the ND or KD with epileptiform activity are shown. g Pathway enrichment analysis showing metabolic pathways affected by the KD. The rich factors represent the ratio of differentially expressed protein in this pathway annotated by number. h The KD increased GABA and glutamate in mouse brains. Levels of GABA, glutamate, glutamine and aspartate were determined in the brains of C57 mice fed with either ND or KD. n = 6 mice. i The KD increased the GABA/glutamate ratio in mouse brains. The ratio of GABA/glutamate was compared between ND- and KD-fed mice. n = 6 mice.
The KD-fed mice had a longer seizure latency than the ND-fed mice (Fig. 1b). Moreover, the KD-fed mice also had less severe seizure symptoms. Among ten mice tested in each group, only three KD-fed mice reached grade III or above; in contrast, most ND-fed mice reached grade IV or V (Fig. 1c). Furthermore, while PTZ induced a similar epilepsy duration in both KD- and ND-fed mice (Fig. 1d), it induced lower seizure frequency in KD-fed mice than in ND-fed mice (Fig. 1e). The epilepsy-relieving effects of the KD were further confirmed via electroencephalography (EEG), which showed that PTZ-induced less electrical activity in KD-fed mice than in ND-fed mice (Fig. 1f).
Proteomic analysis revealed that metabolic pathways, including glutamatergic and GABA-like receptor-related pathways, were altered by the KD (Fig. 1g). These results were consistent with those of the targeted metabolomic analyses, which showed that KD-fed mice had higher levels of glutamate, GABA, and associated metabolites in the brains than those of ND-fed mice (Fig. 1h). Notably, KD feeding resulted in a higher GABA/glutamate ratio (Fig. 1i). These findings highlighted the possibility that the KD exerts seizure-relieving effects via augmenting GABA and GABA/glutamate ratio, both inhibit the excitation of neurons36.
KD inactivated GDH and activated GAD1
The KD altered glutamate homeostasis, prompting us to examine whether it affected the levels of enzymes involved in the Glu/GABA metabolism pathway (Fig. 2a) in vivo37. The KD had negligible impacts on other Glu/GABA metabolism pathway enzymes, except for increased mRNA and protein levels of GAD1 (Fig. 2b, c), consistent with the proteomics results (Figs. 1g, 2d; Supplementary Fig. S1a). As the KD caused neither GDH nor glutaminase (GLS) changes in mouse brains (Fig. 2b, c), it may increase glutamate (see Fig. 1h) by inactivating GDH, which oxidizes glutamate to α-KG, or by inactivating GAD1, which converts glutamate to GABA38. When GAD1 was overexpressed in U87MG (a human brain glioblastoma cell line) cells, it increased GABA levels; however, it failed to decrease glutamate levels (Fig. 2e), suggesting that GAD1 activity is more important for GABA/glutamate ratio than for glutamate levels. Collectively, these results suggest that the KD may decrease GDH-specific activity to increase glutamate in mouse brains.

a Diagram of the enzymes involved in glutamate/GABA metabolism pathways. Enzymes studied in the current study (GDH and GAD1) are highlighted. b, c The KD upregulated GAD1 expression. Protein (b) and mRNA (c) levels of glutamate/GABA metabolism enzymes were determined in the brains of mice fed ND or KD. n = 4 mice. d KD induced SIRT4 and GAD1 expression. The histogram reflects the protein levels of SIRT4 and GAD1. n = 3 mice. e GAD1 had a limited impact on glutamate levels. GAD1 was overexpressed in U87MG cells; GABA and glutamate levels were compared between overexpressed and control U87MG cells. f The KD induced SIRT4 expression. SIRT4 levels in mouse brains were compared between KD- and ND-fed mice. n = 4 mice. g–k SIRT4 was required for the KD to relieve epilepsy in mice. KD- and ND-fed WT and Sirt4–/– mice were each administered with an intraperitoneal PTZ injection before being monitored for epilepsy symptoms. Latency time (g), duration (h), the highest level (i), and frequency of seizures (j) and EEG (k) were monitored. Data were means ± SD, n = 10 mice in each treatment.
SIRT4 was involved in seizure relief via the KD
Notably, SIRT4, a known GDH regulator30, was also induced by the KD (Fig. 2d; Supplementary Fig. S1a). Upregulation of SIRT4 by the KD was confirmed via western blot (Fig. 2f) and real-time qPCR (Supplementary Fig. S1b) in the mice brains. The involvement of SIRT4 in the KD-mediated control of epilepsy was further investigated. 8-week-old wild-type (WT) and Sirt4 knockout male C57 mice (Sirt4–/– mice) were fed with ND or KD. The Sirt4–/– mice grew normally, similar to WT mice, upon chow feeding (Supplementary Fig. S1c, d). Moreover, indiscernible differences in the brain tissue structure were observed using Nissl (Supplementary Fig. S1e) and hematoxylin and eosin (HE) staining (Supplementary Fig. S1f) after 12 weeks of ingesting KD. PTZ-induced pronounced epilepsy symptoms in Sirt4–/– mice, as manifested by faster onset of symptoms such as limb tremors and side-to-side swaying (Fig. 2g), accompanied by longer seizures (Fig. 2h). Sirt4–/– mice exhibited more frequent and intense symptoms, including outstretched hind limbs and front paws tucked under the chin, which suggest death during an episode of status epilepticus (Fig. 2i, j) upon PTZ administration. These behavioral seizures and epileptiform interictal activity in PTZ-challenged Sirt4–/– mice were confirmed using video/EEG (Fig. 2k). Notably, the KD failed to relieve the epilepsy symptoms in PTZ-challenged Sirt4–/– mice (Fig. 2g, k). Collectively, these results suggest that the KD prevents epilepsy, which is dependent on SIRT4 function.
We injected Cre-adeno-associated virus (AAV) into the hippocampus of Sirt4flox/flox mice to delete Sirt4 from hippocampus (Sirt4-cKO, Supplementary Fig. S2a, b). We found that Sirt4-cKO mice phenocopied constitutive Sirt4 knockout mice in response to PTZ challenge and to KD feeding (Supplementary Fig. S2c–i). These results confirmed that Sirt4 in central nervous system is involved in seizure relief effects of the KD.
KD-produced BHB-activated SIRT4 and GAD1 transcription
We investigated how the KD induces SIRT4 and GAD1 expression. The KD has a low carbohydrate content. Therefore, we tested whether sugar deprivation could result in KD-induced SIRT4 expression. Culturing U87MG (Supplementary Fig. S3a) and mouse neuroblastoma (Neuro-2a) (Supplementary Fig. S3b) cells in low-glucose medium failed to induce changes in SIRT4 and GAD1 protein and mRNA levels, ruling out the possibility that the KD drives SIRT4 overexpression solely owing to its low carbohydrate content.
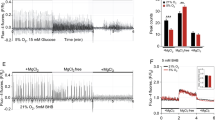
Ketone bodies, such as BHB, AcAc, and acetone, significantly (P < 0.05) increased in the serum (Fig. 3a) and brain (Fig. 3b) of KD-fed mice. However, at physiologic relevant higher levels, only BHB-induced protein and mRNA expression of SIRT4 and GAD1, when U87MG and Neuro-2a cells were treated with each of 5 mM sodium BHB, sodium AcAc, or sodium chloride, or with 0.5 mM acetone (Fig. 3c, d; Supplementary Fig. S3c). Moreover, the BHB-induced protein and mRNA expression of SIRT4 and GAD1 was dose-dependent in mouse hippocampal HT22 cells, and in U87MG (Fig. 3e, f; Supplementary Fig. S3d). To further determine the effect of BHB on SIRT4 and GAD1 expression, we overexpressed β-hydroxybutyrate dehydrogenase 1 (BDH1), which catalyzes the first reaction of BHB oxidation to convert BHB back into AcAc39, in HT22, U87MG, and Neuro-2a cells. We found BDH1 overexpression, while decreasing cellular BHB levels (Fig. 3g), decreased the expression of SIRT4 and GAD1 in HT22, U87MG, and Neuro-2a cells (Fig. 3h, i). Conversely, knockdown BDH1 with small interfering RNAs that increased intracellular BHB concentration (Fig. 3j), increased mRNA and protein levels of SIRT4 and GAD1 in U87MG and HT22 cells (Fig. 3k, l; Supplementary Fig. S3f). These results, together with the fact that SIRT4 and GAD1 degradation in protein synthesis-inhibited CHX-treated cell lysates were unaffected by BHB (Supplementary Fig. S3g), supported the hypothesis that the KD promotes SIRT4 and GAD1 transcription by upregulating BHB levels.

a, b The KD increased mice ketone body levels. Serum (a) and brain (b) levels of the BHB, AcAc, and acetone were detected in ND- and KD-fed mice. c, d BHB increased SIRT4 and GAD1 expression levels. U87MG cells were treated with ketone bodies and the protein (c) and mRNA (d) levels of SIRT4 and GAD1 were measured. e, f BHB induced SIRT4 and GAD1 expression in a dose-dependent manner. Protein (e) and mRNA (f) levels of SIRT4 and GAD1 were determined in HT22 cells that were treated with BHB at the indicated concentrations. g BDH1 overexpression decreased BHB levels. The BHB levels were measured in HT22, U87MG and Neuro-2a cells and in BDH1-overexpressing HT22 cells, U87MG and Neuro-2a cells. h, i BDH1 overexpression decreased SIRT4 and GAD1 expression. Protein (h) and mRNA (i) levels of SIRT4 and GAD1 were detected in HT22 cells, U87MG, and Neuro-2a cells and in BDH1-overexpressing HT22 cells, U87MG and Neuro-2a cells. j–l BDH1 knockdown increased BHB levels and increased SIRT4 and GAD1. BDH1 was silenced with independent siRNAs in HT22 and U87MG cells. BHB levels (j), protein (k), and mRNA(l) levels of SIRT4 and GAD1 were compared between siRNA-untreated and -treated cells.
BHB inhibited HDAC1/HDAC2, thus activating SIRT4 and GAD1 transcription
To investigate the mechanism by which BHB promotes SIRT4 and GAD1 expression in cells, we used actinomycin D to inhibit mRNA synthesis and measured the half-lives of SIRT4 and GAD1 mRNA in U87MG cells. No significant differences in SIRT4 and GAD1 mRNA half-lives were observed between the BHB-treated and control cells (Fig. 4a; Supplementary Fig. S4a), indicating that BHB promotes SIRT4 and GAD1 mRNA synthesis rather than inhibiting mRNA degradation. Given that BHB is an established inhibitor of histone deacetylases (HDACs)40, we analyzed the chromatin accessibility of the SIRT4 and GAD1 promoter regions and found that BHB treatment markedly enhanced chromatin accessibility of the SIRT4 and GAD1 promoters (Fig. 4b; Supplementary Fig. S4b). This result is consistent with the key function of BHB, which enables the transcriptional machinery to inhibit histone deacetylation40.

a mRNA degradation did not account for the BHB-promoted SIRT4 and GAD1 mRNA increase. SIRT4 mRNA in U87MG cells was detected at the indicated time points with 10 mM actinomycin D. b BHB increased chromatin accessibility of the SIRT4 promoter. Quantitative PCR was performed on DNase I-pretreated nuclei of U87MG cells that were treated with BHB with the indicated concentrations. c BHB specifically elevated H3K27Ac. Histone acetylation levels were assayed in U87MG cells and in BHB- or trichostatin A (TSA)-treated U87MG cells, employing site-specific and pan-anti-acetyl-lysine (Ac) antibodies. d BHB-treatment elevated H3K27Ac at the promoter regions of SIRT4 and GAD1. The H3K27Ac antibody was used for immunoprecipitation (IP) after BHB treatment, and qRT-PCR of SIRT4 and GAD1 promoter output was performed. e, f The HDAC1/HDAC2-specific inhibitor FK228 increased SIRT4 and GAD1 protein (e) and mRNA (f) levels. The ability of FK228 to upregulate SIRT4 and GAD1 expression was tested in U87MG cells. g, h BHB elevated SIRT4 and GAD1 expression, dependent on HDAC1 and HDAC2 acetylation. The effects of BHB (10 mM) treatment on SIRT4 and GAD1 expression were assayed in HeLa cells with HDAC1, HDAC2, or double knockout of both (g, quantified 4 replicates in right), and in U87MG cells overexpressing HDAC1, HDAC2, or both (h). i HDAC1 and HDAC2 knockout enhanced H3K27Ac at the SIRT4 promoter. ChIP assays were performed for the promoter region of SIRT4, using the H3K27Ac antibody for IP and qRT-PCR for the SIRT4 promoter output.
Tracing the acetylation levels at different sites on histone H3 via immunoblotting revealed that the acetylation levels at H3K27 increased with BHB (Fig. 4c). Moreover, chromatin immunoprecipitation (ChIP) assays showed that BHB enhanced H3K27Ac levels and histone interactions with SIRT4 and GAD1 promoters (Fig. 4d), suggesting that BHB-induced SIRT4 and GAD1 transcriptional activation may correlate with H3K27Ac, which is deacetylated by the HDAC family, including HDAC1–HDAC1041.
Treatment of U87MG cells with different HDAC inhibitors revealed that neither the HDAC3 inhibitor RGFP966 (Supplementary Fig. S4c, d) nor the HDAC 4/5/7/9 inhibitor TMP195 (Supplementary Fig. S4e, f) affected the expression of SIRT4 and GAD1, and only the HDAC1 and HDAC2 inhibitor FK228 upregulated protein (Fig. 4e) and mRNA (Fig. 4f) levels of SIRT4 and GAD1.
To determine whether HDAC1 and HDAC2 are involved in the transcriptional regulation of SIRT4 and GAD1 via BHB, we knocked out either HDAC1 or HDAC2 to induce SIRT4 and GAD1 transcription and protein expression; however, only knockout of both HDAC1 and HDAC2 caused pronounced increase of SIRT4 and GAD1 expression and prevented BHB from activating SIRT4 and GAD1 transcription in HeLa cells (Fig. 4g). In contrast, the overexpression of HDAC1 and HDAC2 in U87MG (Fig. 4h) and HeLa cells (Supplementary Fig. S4g) decreased SIRT4 and GAD1 expression. These results, consistent with those of ChIP assays, showed that HDAC1 and HDAC2 knockouts extensively increased the H3K27Ac of histones, which interacted with the SIRT4 and GAD1 promoters in HeLa cells (Fig. 4i). HDAC1/HDAC2-mediated histone deacetylation inhibits the transcription of GAD142 and this study confirmed that BHB activates SIRT4 and GAD1 transcription by inhibiting HDAC1- and HDAC2-induced H3K27 deacetylation.
Lysine carbamylation-activated GDH
Although GAD1 activity was unresponsive to SIRT4 variation (Supplementary Fig. S5a, b) in SIRT4-silenced and SIRT4-overexpressing HEK293T cells, GDH was activated (Fig. 5a) and inactivated (Fig. 5b), respectively, suggesting that GDH is inactivated by GDH mono-ADP-ribosylation43. However, the activity of the ADP-ribosylation-null mutant GDHC119G 44 was still responsive to SIRT4 variation (Fig. 5a, b), suggesting that a mechanism other than ADP-ribosylation regulates GDH enzyme activity to convert glutamate into α-KG.

a, b The activity of the ADP-ribosylation-null GDHC119G mutant was under regulation by SIRT4. Flag-tagged GDH or GDHC119G was ectopically expressed in HEK293T cells or SIRT4-silenced HEK293T cells (a). Flag-tagged GDH or GDHC119G was ectopically expressed in HEK293T cells or SIRT4-overexpressing HEK293T cells (b). The specific activities of GDH and the GDHC119G mutant were determined and compared. c GDH interacted with CPS1. Flag-tagged CPS1 or GDH was ectopically expressed in HEK293T cells together with individual HA-tagged GDH or CPS1 as indicated. Flag bead- or HA bead-purified proteins were subject to western blotting analysis. d Carbamylation of endogenous GDH relied on CPS1 to generate CP. Endogenous GDH of HeLa cells was purified via IP with Protein A beads conjugated with the GDH antibody, and carbamylation of the purified GDH was detected. e KD decreased mouse brain mitochondrial CP-K levels. Mitochondrial CP-K levels were detected in brains of mice fed with either ND or KD. f Carbamylation activated GDH. Flag-tagged GDH was ectopically expressed in HEK293T cells treated with CP (5 mM) and purified via IP with Flag beads. The specific activity of GDH was determined. g GDH K162 was carbamylated. MS/MS spectra from tryptic peptide libraries of HEK293T cells matching MS/MS spectra from synthetic peptide (composed of 12C, 1H, 14N, and 16O) identified GDH CP-K162. h Screening of the major regulatory carbamylation sites in GDH. Detected carbamylated residues were each mutated to arginine (R), and the specific activity of each mutant GDH was compared with that of WT GDH. i In vitro formation of CP-K162. The synthetic GDH K162-containing peptide was treated with 10 mM CP, followed by mass spectrometry analysis.
Glutamate oxidation via GDH produces α-KG and ammonia, which are converted to CP that spontaneously forms CP-K45,46, which is reversed by SIRT431. GDH interacted with CPS1, an enzyme that spontaneously produces CP to carbamylate lysine (Fig. 5c)45. Moreover, CPS1 knockout in HeLa cells reduced the level of GDH CP-K (Fig. 5d). Furthermore, in the mitochondria of mouse brains, the KD-induced lower CP-K levels (Fig. 5e). Thus, we investigated the possibility that the KD induces SIRT4, which inactivates GDH by decarbamylating it. Moreover, the incubation of recombinant GDH with 5 mM CP increased GDH CP-K levels and activated GDH (Fig. 5f), suggesting that carbamylation activates GDH.
Using a Thermo Q-Exactive HF (QE) Quadrupole-Obitrap mass spectrometer, we detected multiple CP-modified lysine sites, including K84, K162, K191, K200, K352, and K503 (Fig. 5g; Supplementary Fig. S6) in GDH purified from U87MG cells. Eliminating potential carbamylation sites in GDH by switching lysine to arginine (K/R) revealed that GDHK162R, GDHK352R, and GDHK503R inactivated GDH and diminished the ability of CP to activate GDH (Fig. 5h), suggesting that these were the key carbamylation sites that activated GDH. To confirm that CP spontaneously carbamylates GDH, we synthesized a GDH peptide containing K162, which is evolutionarily conserved from Trachemys scripta elegans to Homo sapiens (Supplementary Fig. S7a), and close to the active site of GDH, K183 (Supplementary Fig. S7b). Treatment with 5 mM CP induced carbamylation in the K162-containing peptide but not in the peptide in which K162 was replaced by arginine (K162/R) (Fig. 5i). Collectively, these results suggest that lysine carbamylation enhances GDH activity without affecting its expression.
SIRT4 decarbamylated and inactivated GDH to increase both glutamate and GABA but GABA/glutamate ratio
GDH and SIRT4 were co-immunoprecipitated when they were simultaneously expressed in HEK293T cells (Supplementary Fig. S8a). Moreover, the decarbamylase activity of SIRT4 was confirmed by detecting decarbamylated peptide formation from the K162R-containing GDH peptide using reverse-phase high-performance liquid chromatography (Fig. 6a). This suggests that SIRT4 is a decarbamylase of GDH, which was confirmed by the CP-K level and specific activity of ectopically expressed GDH in HEK293T cells (Fig. 6b). Endogenous GDH in U87MG cells (Supplementary Fig. S8b) cultured in CP-supplemented media was decreased by SIRT4 overexpression, and not by that of SIRT3 or SIRT5. Conversely, silencing SIRT4 in HEK293T cells increased CP-K levels and the specific activity of GDH (Fig. 6c). In contrast, SIRT4 had no effect on the specific activity of GDHK162R, which was diminished when it was co-expressed with GDHK162R (Fig. 6c, d). Affinity-purified GDH from WT mice brains were carbamylated less and were less active than that from Sirt4–/– mouse brains (Fig. 6e, f). Moreover, the KD, which upregulated SIRT4 in the mouse brains (Fig. 2d–f), decreased CP-K levels and inactivated endogenous GDH (Fig. 6e, f). By using 13C-glutamine as a tracer (Supplementary Fig. S8c), we detected the relative cytosolic and mitochondrial 13C-GABA and 13C-α-KG levels in both U87MG cells and SIRT4-silenced U87MG cells. SIRT4 knockdown decreased the GABA and glutamate levels, but increased α-KG levels (Fig. 6g). Moreover, the GABA and glutamate levels were decreased in Sirt4–/– mouse brains (Fig. 6h), and the GABA/glutamate ratio was not altered by SIRT4 removal (Fig. 6i). These results suggest that GAD1 is not the downstream of SIRT4, and collectively supports the idea that SIRT4 decarbamylates and inactivates GDH.

a SIRT4 decarbamylated GDH CP-K162. RP-HPLC analyses were used to detect decarbamylation of the synthetic GDH CP-K162 peptide catalyzed by SIRT4. b SIRT4 decarbamylated GDH. HA-tagged mitochondrial SIRT3, SIRT4, and SIRT5 were each individually expressed in untreated and CP (5 mM)-treated HEK293T cells. The GDH ectopically expressed in these cells were purified and subject to CP-K and specific activity analysis. c, d K162 was the major regulatory carbamylation site in GDH. Ectopically expressed GDH and GDHK162R mutant from HEK293T cells and SIRT4 knockdown (c) or SIRT4 overexpression (d) HEK293T cells assayed for CP-K and specific activities. e GDH in Sirt4–/– mouse brain was more carbamylated. The GDH protein was purified using Protein A beads conjugated with the GDH antibody from the brains of WT and Sirt4–/– mice that were fed either ND or KD. GDH carbamylation was detected for purified GDH. f GDH was inactivated by the KD only in WT mouse brains. The WT and Sirt4–/– mice were each fed the ND and KD from week 8. The mouse brain GDH purified at week 20 was assayed for specific activity. g SIRT4 knockdown decreased GABA and glutamate and increased α-KG. Culture media was supplemented with 2 mM 13C-glutamine. The M + 5 glutamate and α-KG (carbons fully labeled) and M + 4 GABA (carbons fully labeled) were determined in the mitochondria and cytoplasm of WT and SIRT4 knockdown U87MG cells after 6 h of chasing. h Sirt4 ablation caused more pronounced GABA elevation than glutamate elevation. The levels of GABA and glutamate were compared between WT and Sirt4–/– mouse brains. i Sirt4 ablation did not alter GABA/glutamate ratio in mouse brains. GABA/glutamate ratios were detected in WT and Sirt4–/– mouse brains.
BHB effectively relieved epilepsy
To determine whether BHB alone may have seizure-relieving effects, the ND was supplemented with a BHB ester via oral gavage, which increased serum BHB levels > 4-fold (Supplementary Fig. S9a); however, it did not affect body weight (Supplementary Fig. S9b) and food intake (Supplementary Fig. S9c).
BHB supplementation induced higher GABA/glutamate ratio in the brains (Fig. 7a), longer latency of seizures (Fig. 7b), less severe seizures (Fig. 7c), and lower frequency of them (Fig. 7d). However, it had no impact on seizure duration (Fig. 7e) in PTZ-induced C57 mice, reinstating the antiepileptic efficacy of the KD (see Fig. 1b–e). Notably, no seizure-relieving efficacy was observed in PTZ-treated Sirt4–/– mice (Fig. 7b, e). The lower potency of BHB in inhibiting epilepsy in Sirt4–/– mice than in WT mice was further confirmed by video/EEG (Supplementary Fig. S9d). The carbamylation (Supplementary Fig. S9e) and activity (Supplementary Fig. S9f) of mice GDH were decreased by BHB in WT but not in Sirt4–/– mouse brain. Moreover, H3K27Ac (Supplementary Fig. S9g) and SIRT4 and GAD1 protein levels (Supplementary Fig. S9e) were increased by BHB. These are all consistent with the findings that BHB elevates H3K27Ac to overexpress SIRT4 and GAD1 and shift GABA/glutamate ratio in the brains.

a BHB increased the GABA/glutamate ratio in WT and Sirt4–/– mouse brains. Data were represented as individual values. b–e BHB alone SIRT4-dependently relieved seizure phenotypes. The ND- and BHB-fed WT and Sirt4–/– mice were subject to measurements after they were administered with PTZ. Latency time (b), the highest level (c), the frequency (d) and the duration of seizures (e) were monitored. Data were means ± SD, n = 10 mice in each treatment. f–k GDH inhibition potentiated BHB to relieve seizures. BHB or the GDH inhibitor EGCG alone, or BHB together with EGCG were employed to treat Sirt4–/– mice. The seizure-relieving effects of these treatments were monitored after mice were administered with PTZ. Brain GABA and glutamate (f) and GABA/glutamate ratio (g), latency time (h), the highest level of seizures (i), the frequency of seizures (j) and the duration of seizures (k) were monitored. Data were means ± SD, n = 10 mice in each treatment. l Schematic illustration showing KD upregulates brain GABA levels and GABA/glutamate ratio. Compared to ND, KD promotes BHB, which inhibits HDAC1/HDAC2 to upregulate SIRT4 and GAD1 transcription, results in inactivated GDH and activated GAD1, and consequently higher GABA and GABA/glutamate ratio.
Inactivation of GDH through SIRT4-catalyzed decarbamylation may account for the inability to relieve PTZ-induced epilepsy in Sirt4–/– mice. We used epigallocatechin-3-gallate (EGCG), an inhibitor of GDH47, to reverse Sirt4 knockout effects, i.e., preserving and directing glutamate to GABA. Oral gavage of EGCG (100 mg/kg) and BHB (500 mg/kg) for 4 weeks48, which had no impact on body weight (Supplementary Fig. S10a) and food intake (Supplementary Fig. S10b) of Sirt4–/– mice, helped BHB to increase GABA and glutamate levels (Fig. 7f) and the GABA/glutamate ratio (Fig. 7g). It also led to remission of epileptic symptoms, which included a slower onset of the first seizure after PTZ administration (Fig. 7h), reduced intensity (Fig. 7i) and frequency of seizures (Fig. 7j), and shorter duration of seizures (Fig. 7k). These were consistent with the findings of the video/EEG for PTZ-induced Sirt4–/– mice (Supplementary Fig. S10c). Lastly, the seizure-relieving efficacy of BHB was not observed in PTZ-treated Sirt4-cKO mice (Supplementary Fig. S10d–j). These findings collectively support the notion that high GABA/glutamate ratio and GABA levels in central nerve systems induced by BHB are required to relieve epilepsy.
Discussion
Our results revealed that the KD-produced BHB accounts for antiepileptic efficacy of the KD. By inhibiting HDAC1/HDAC2, BHB increases H3K27Ac levels and facilitates the transcription of SIRT4 and GAD1, which function synergistically to inhibit neuronal activation. By upregulating SIRT4, the decarbamylase of GDH, BHB inactivates GDH and accumulates glutamate for GABA production. Moreover, upregulation of GAD1 by BHB favors the production of the inhibitory neurotransmitter GABA from glutamate and increases the GABA/glutamate ratio to inhibit epilepsy (Fig. 7l)49. Notably, the activation of both SIRT4 and GAD1 seems to be essential for BHB to exert antiepileptic effects, as BHB exhibits limited antiepileptic potency in Sirt4–/– mice although BHB increased brain GABA in both WT and Sirt4–/– mice brains. Inhibiting GDH alone to proportionally increase glutamate and GABA also has limited antiepileptic potency. However, the GDH inhibitor EGCG potentiates BHB to inhibit epilepsy in Sirt4–/– mice. This design suggests that a sufficiently high level of GABA is required in addition to high GABA/glutamate ratio to accomplish neuronal inhibition, given the array of other excitatory neurotransmitters besides glutamate that activate neurons50,51. It is worth to note that this mechanism may be related to mammal physiology. BHB and/or ketone bodies may be fat-sufficient signals, which inhibit neuronal excitation, required for hunting in mammals52, to keep energy expenditure in check and therefore exert antiepileptic efficacy. Therefore, it makes physiologic sense that KD and BHB inhibit epilepsy. KD induces cardiac fibrosis53 as high fat diet do54, supports that KD may generate fat-sufficient signals.
BHB is an endogenous and specific inhibitor of class I HDACs40. Moreover, GAD1 is down-regulated by HDAC1/HDAC2-mediated histone de-acetylation42. These results supported the upregulation of GAD1 by BHB or KD. The identification of BHB that upregulates SIRT4 in the mouse brains adds important clues to how BHB, whose production is upregulated by the KD, exerts antiepileptic effects. Therefore, in addition to its ADP-ribosyltransferase30, lipoamidase55 and deacylase56 activities, the newly identified de-carbamylase activity of SIRT431 expands its roles in glutamate and GABA metabolism and neuronal activity regulation.
In addition to many mechanisms that had been proposed to explain the use of KD to constrain epilepsy, the identification of BHB as a major antiepileptic metabolite in KD is clinically noteworthy. As KD has been used as a dietary treatment for refractory epilepsy since the 1920s, side effects include poor taste and low compliance57,58, and prolonged KD administration tends to cause gastrointestinal disturbances and vomiting59, malnutrition60, cardiovascular diseases61, kidney stones62, and poor growth63. However, these limitations may be overcome by employing BHB or its efficacy-optimized molecules to replace the KD, which have fewer limitations in terms of toxicity and high doses.
Overall, our study highlights that BHB may serve as fat-rich signals to inhibit neuronal excitation. Moreover, the identification of BHB as an antiepileptic molecule has translational potential for the development of low toxic and high tolerable antiepileptic drugs. We were unable to obtain samples from human patients, mechanistic studies were performed on cultured cells, and in vivo studies were performed on mice model. Given that we study the GABA and glutamate homeostasis on epilepsy, we used non-competitive GABA(A) receptor inhibitor PTZ to generate mouse epilepsy model, which simulates the generalized symptoms of clinical human myoclonic epilepsy and is used to screen anti-epilepsy drugs. However, PTZ model fails to find the anti-non-spasmodic seizure effect of lamotrigine and tiagabine, and other epilepsy models, especially genetic animal epilepsy models are necessary to be employed to confirm whether KD and/or BHB will exert general anti-epilepsy efficacies.
Materials and methods
Cell culture
HEK293T (human embryonic kidney), HeLa (human adenocarcinoma), U87MG (human glioblastoma), and Neuro-2a (mouse neuroblastoma) cells were cultured in normal DMEM (Gibco) supplemented with 10% fetal bovine serum (FBS) (Gibco), 100 U/mL penicillin (Invitrogen) and 100 μg/mL streptomycin (Invitrogen). All cells were cultured an incubator at 37 °C and 5% CO2. Cell treatments are carried out by adding chemicals to the culture media to the levels specified in legends. BHB treatment takes 12 h.
Animal experiments
Four-week-old C57BL/6 Sirt4–/– mice (constructed using CRISPR-mediated deletion of exons 3 and 4 of the mouse Sirt4 gene) were purchased from GemPharmatech Co., Ltd. (Shanghai, China). Genotypes were determined via PCR using genomic DNA obtained from tails and two primer pairs (sequences are provided in Supplementary Table S1). All mice were housed in a specific pathogen-free facility at 20–22 °C on a 12 h light/dark cycle with ad libitum access to food and water. All experiments were started when mice were 6 weeks old.
Sirt4-cKO mouse was generated based on C57BL/6 Sirt4flox/flox mice. 2–3-month-old C57BL/6 Sirt4flox/flox mice, purchased from Shanghai Research Center of the Southern model organisms, were anesthetized with a mixture of ketamine (100 mg/kg) and xylazine (10 mg/kg). Thirty three-gauge syringe needles (Hamilton, Reno, Nevada) were used to bilaterally infuse 1.0 μL of AAV virus into mouse dorsal hippocampus (AP = –2.0, ML = ±1.5, and DV = –2.0 from Bregma (mm)) or 0.5 μL of AAV virus into nucleus accumbens (NAc) (AP = +1.5, ML = + 1.6, and DV = –4.4 from Bregma (mm) at 10°) at a rate of 0.2 μL (0.1 μL for NAc) per min and the needle remained in place for 5 min before removal to prevent backflow. Intra-hippocampal microinjection of AAV vectors containing the gene for Sirt4 or GFP alone were prepared by Genechem, Shanghai, China. AAV-injected mice were allowed to recover for 21 days before behavioral testing.
Carbon dioxide asphyxiation was adapted to euthanize mice. The animals were brought from their home cages to the CO2 euthanasia apparatus in the procedure room. Remove the filter top from the cage, and cover the cage with the stainless-steel euthanasia lid. Turn on the flow of CO2 to the cage by using the flow meter attached to the pressure regulator. Expose the animals to CO2 until complete cessation of breathing for a minimum of 2 min after 5 to 10 min CO2 exposing. Continue the following experiments after visualizing all mice in the cage were euthanized.
Adult male mice were euthanized, and serum and brain tissues were collected according to standard procedure. Briefly, brains were perfused with saline to remove blood cells before isolation, and the tissues were quickly removed and snap frozen in liquid nitrogen for further analyses. All animal procedures were conducted in accordance with the animal care committee at Fudan University, Shanghai, China.
Mouse KD or BHB treatment
Eight-week-old WT and Sirt4–/– mice received an ND (Bio-Serv, F0761) or KD (Bio-Serv, F3666) with free access to drinking water for 12 weeks. BHB ester (HVMN, 200 µL by oral gavage daily, 80 mg per mouse) treatments to mice last 4 weeks before analysis.
PTZ-induced acute seizure mouse model
Twenty-week-old male mice received a single dose of intraperitoneal injection of PTZ (Sigma), with the dosage ranging between 40 and 45 mg/kg of body weight after being fed ND or KD. Mice were observed for 10 min after each injection.
The scores of seizure were staged as follows: 0 grade: no response; I grade: facial movements, including blinking, twitching, rhythmic chewing, etc; II grade: I grade plus rhythmic nodding; III grade: II grade plus forelimb clonic; IV grade: III grade plus standing on hind legs; V grade: IV grade plus falling.
Mouse EEG
Under Ketamine/Xylazine anesthesia, chronic EEG electrodes were surgically implanted using aseptic technique in mice. Micro-screws anchored into the cranium served as epidural electrodes and were connected to a microplug adhered to the cranium with dental acrylic. Recording electrodes were positioned bilaterally over each hemisphere (anterior-posterior, –2.0 mm from bregma; lateral, ±1.75 mm); a reference electrode was placed anterior to bregma (anterior-posterior, +0.5 mm) and a ground electrode was placed over the cerebellum. Mice were allowed to recover for 5–7 days before recording. The Telefactor Video/EEG Monitoring System was used to record EEGs and to videotape behavior simultaneously from un-anesthetized free-moving mice. The low-frequency filter setting was 0.3 Hz, and the high-frequency filter was set at 70 Hz.
CRISPR/Cas9-mediated deletion of HDAC1 and HDAC2
HDAC1-knockout and HDAC2-knockout HeLa were generated by a CRISPR/Cas9 plasmid, respectively targeting HDAC1 and HDAC2. The plasmids were constructed by cloning the annealed sgRNA into px459 vector. The sgRNAs were designed by CRISPR Design website (http://crispr.mit.edu) and the sequences are listed in Supplementary Table S1.
Plasmid construction
The cDNA encoding full-length human SIRT3, SIRT4, SIRT5, GDH, GAD1, BDH1, CPS1, HDAC1, and HDAC2 was cloned into Flag, Myc, HA-tagged vectors (pcDNA3.1-Flag; pcDNA3.1-Myc; pcDNA3.1-HA). Point mutations of GDH were generated by Mut Express II Fast Mutagenesis Kit V2 (Vazyme).
Metabolomics analysis
Metabolite extraction
Frozen tissues were homogenized in 80% methanol (methanol:water = 4:1) and centrifuged at 14,000× g for 15 min (4 °C). The supernatant was transferred to a new tube and lyophilized into a pellet. The sample was resuspended in 1500 μL of extract solution (acetonitrile:methanol:water = 2:2:1, with isotopically labelled internal standard mixture). After 30 s of vortexing, samples were sonicated for 10 min in an ice-water bath. Next, the samples were incubated for 1 h at –40 °C and centrifuged at 12,000× g for 15 min at 4 °C. The resulting supernatant was transferred to a fresh glass vial for analysis.
LC-MS/MS analysis
LC-MS/MS analyses were performed using a UHPLC system (Vanquish, ThermoFisher Scientific) with an UPLC BEH Amide column (2.1 mm × 100 mm, 1.7 μM) coupled to a QExactive HFX mass spectrometer (Orbitrap MS, ThermoFisher Scientific). The mobile phase consisted of 25 mmol/L ammonium acetate and 25 mmol/L ammonia hydroxide in water (pH = 9.75) (A) and acetonitrile (B). The analysis was carried out with an elution gradient as follows: 0–0.5 min, 95% B; 0.5–7.0 min, 95%–65% B; 7.0–8.0 min, 65%–40% B; 8.0–9.0 min, 40% B; 9.0–9.1 min, 40%–95% B; 9.1–12.0 min, 95% B. The column temperature was 30 °C. The temperature of the autosampler was 4 °C, and the injection volume was 2 μL.
The QE HFX mass spectrometer was used due to its ability to acquire MS/MS spectra in information-dependent acquisition mode under the control of acquisition software (Xcalibur, ThermoFisher Scientific). In this mode, the acquisition software continuously evaluated the full-scan MS spectrum. The ESI source conditions were set as follows: sheath gas flow rate of 30 Arb, Aux gas flow rate of 25 Arb, capillary temperature of 350 °C, full MS resolution of 60,000, MS/MS resolution of 7500, collision energy of 10/30/60 in NCE mode, and spray voltage of 3.6 kV (positive) or –3.2 kV (negative).
Data analysis
Raw data were converted to the mzXML format using ProteoWizard and processed using an in-house programme, which was developed using R and based on XCMS, for peak detection, extraction, alignment, and integration. An in-house MS2 database (BiotreeDB) was applied for metabolite annotation. The cut-off for annotation was set at 0.3.
13C labelling of metabolites
Cells were plated in 10-cm dishes for 24 h before labelling. The next day, the medium was changed for unlabelled medium without glutamine 1 h before adding the 13C5-glutamine medium. Cells were incubated in labelling media containing 2 mM 13C5-glutamine (Sigma) for 6 h before collection. At the time of collection, cells were washed with prechilled saline, lyzed with (40%) methanol:(40%) acetonitrile:(20%) water with 0.1 M formic acid, and processed as described above (Metabolomics). Metabolites were reconstituted in 100 μL of 0.1% formic acid in LC/MS-grade water, vortex-mixed, and centrifuged to remove debris. The supernatants were analyzed using LC-MS/MS.
Proteomics analysis
Protein digestion
The procedures for protein digestion were performed according to the filter-aided sample preparation (FASP) technique64 with slight modifications. Briefly, FASP digestion was adapted for the following procedures in Microcon PL-10 filters. Following a three-time buffer displacement with 4 M guanidine hydrochloride and 100 mM Tris-HCl, pH 8.0, proteins were reduced using 10 mM DTT at 37 °C for 30 min, followed by alkylation with 30 mM iodoacetamide at 25 °C for 45 min in the dark. Digestion was carried out with trypsin (enzyme:protein = 1:50) at 37 °C overnight after washing with 20% acetone nitrile and digestion buffer (30 mM Tris-HCl, pH 8.0). After digestion, the solution was filtrated out, the filter was washed with 15% ACN, and all filtrates were pooled, vacuum-dried, and kept at –80 °C until use.
Carbamyl peptide enrichment
Carbamyl peptide enrichment was performed using pan-anti-carbamyl lysine antibody (Abcam, Cat# ab175132) conjugated with protein A beads (Sigma, Cat# 16-156). Peptides were mixed with antibody-conjugated beads at 4 °C for 6–8 h under rotary shaking. The mixture was briefly centrifuged at 1000× g (< 10 s), and the supernatant was carefully removed. Antibody-conjugated beads were subsequently washed thrice with phosphate buffered saline (PBS) buffer. Finally, the bound carbamyl peptides were eluted with 30% ACN/0.1% TFA and concentrated in a SpeedVac. Before MS analysis, all carbamyl peptides were desalted.
LC-MS/MS analysis
LC-MS analysis was performed using a nanoflow EASYnLC 1200 system (ThermoFisher Scientific, Odense, Denmark) coupled to an Orbitrap Fusion Lumos mass spectrometer (ThermoFisher Scientific, Bremen, Germany). A one-column system was adopted for all analyses. Samples were analyzed on a C18 analytical column (75 µm i.d. ×25 cm, ReproSil-Pur 120 C18-AQ, 1.9 µm (Dr. Maisch GmbH, Germany)) constructed in-house. The mobile phases consisted of Solution A (0.1% formic acid) and Solution B (0.1% formic acid in 80% ACN). The derivative peptides were eluted using the following gradients: 2%–5% B in 2 min, 5%–35% B in 100 min, 35%–44% B in 6 min, 44%–100% B in 3 min, 100% B for 10 min, at a flow rate of 200 nL/min. Data-dependent analysis was employed in MS analysis: the time between master scan was 3 s, and fragmented in HCD mode, normalized collision energy was 30.
Data processing and analysis
Raw MS files were searched using Protein Discoverer (version 2.4, ThermoFisher Scientific) with Mascot (version 2.7.0, Matrix Science). Data were processed using the UniProt mouse protein database (55,462 entries). The mass tolerances were 10 ppm for precursor and fragment mass tolerance 0.05 Da. Up to two missed cleavages were allowed. The search engine set protein N-acetylation, pyroglutamate on peptide N termini, oxidation on methionine, and carbamylation on Lys as variable modifications and cysteine carbamidomethylation as a fixed modification.
Recombinant SIRT4 expression and purification
Recombinant SIRT4 expression was performed as previously described with slight modifications65. The NAD+-dependent deacetylase domain encompassing residues 27–314 of SIRT4 was subcloned into a pSUMOH10 vector (modified based on pET28b) containing an N-terminal 10× HIS-SUMO tag. SIRT427–314 were expressed in Escherichia coli BL21 (DE3) strain (Transgene, China). After induction with 0.2 mM isopropyl β-D-thiogalactoside at 16 °C in LB medium supplemented with 0.1 mM ZnCl2 overnight, cells were harvested by centrifugation at 4000× g for 20 min at 4 °C. The pellets were resuspended in buffer A containing 20 mM Tris-HCl, pH 7.5, 500 mM NaCl, 5% glycerol, and 20 mM imidazole and were then disrupted using an EmulsiFlex-C3 homogenizer (Avestin) at 4 °C. The lysate was further cleared by centrifugation at 14,000× g for 60 min at 4 °C, and the supernatant was loaded onto a His-trap affinity column (GE Healthcare). After washing with buffer A, bound proteins were subjected to on-column cleavage by ULP1 SUMO protease. The tag-free SIRT427–314 protein was collected as flow-through, centrifuge-concentrated, and then purified on an anion-exchange QHP column followed by size exclusion chromatography on a Superdex 75 column (GE Healthcare). Purified peak fractions were pooled, concentrated, aliquoted, and stored at −80 °C for future use.
MS-based in vitro decarbamylation assays
The ability of SIRT4 to hydrolyze carbamyl lysine was measured using MALDI TOF/TOF. Carbamyl peptide (200 µM) was incubated with 5 µM SIRT3, SIRT4, or SIRT5 in reaction buffer containing 50 mM Tris-HCl, pH 7.8, 1 mM DTT, and 1 mM MgCl2 without or with 2 mM NAD+ for 2 h at 37 °C. Before MS analysis, all carbamyl peptides were desalted and eluted using C18 tips. Samples were analyzed using a 4800 Plus MALDI TOF/TOF Analyzer (Applied Biosystems/MDSSCIEX) and operated in Reflector Positive mode using 4000 Series Explorer Software. The laser intensity was set at 3500 V.
SDS-PAGE and western blotting assay
SDS-PAGE and western blotting assay were performed following standard protocols. Briefly, cells were harvested with loading buffer containing 50 mM Tris-HCl pH 6.8, 10% glycerol (v/v), 2% SDS (w/v), 4% β-mercaptoethanol (v/v), and 0.0012% bromophenol blue (w/v). For western blotting analysis, each sample was subjected to SDS-PAGE and transferred to nitrocellulose membranes (GE Healthcare Life Science). The membranes were blocked in 5% (w/v) skim milk in Tris-buffered saline with 0.1% (v/v) Tween-20 (TBST) for 1 h at room temperature and were then probed with primary antibodies in antibody dilution buffer (QuickBlockTM, Beyotime) at 4 °C overnight. After incubation with horseradish peroxidase-conjugated secondary antibodies in TBST (containing 5% skim milk), membranes were developed using ECL-Plus (Thermo Fisher Scientific) and visualized using Typhoon (GE Healthcare Life Science).
Protein half-life assays
Cells were treated under the indicated conditions. For half-life studies, cycloheximide (100 mg/mL, Sigma) was added to the medium. Cells were collected at the indicated time points and protein concentrations were measured, then analyzed by immunoblot analysis.
ChIP assays
ChIP assays were conducted using the ChIP Assay Kit (Beyotime). First, U87MG and HeLa cells treated with or without BHB were crosslinked by 1% formaldehyde for 10 min. DNA was then sonicated into fragments with a mean length of 200–500 bp, and sheared chromatin was immunoprecipitated with antibodies against H3K27Ac (Abcam) or non-specific rabbit IgG (Santa Cruz) overnight at 4 °C. The precipitated DNA fragments were then identified by PCR and quantified by quantitative real-time PCR (qRT-PCR).
RNA isolation and qRT-PCR analyses
RNA was extracted from cells using TRIzol (Transgene, China) according to the manufacturer’s instructions, and the concentration/purity were determined using NanoDrop Spectrophotometer (ThermoFisher Scientific). cDNA was synthesized using the HiScript III 1st Strand cDNA Synthesis Kit (Vazyme) according to the manufacturer’s instructions. qRT-PCR was performed with SYBR qPCR Master Mix (Vazyme) on a Real-Time Thermocycler (BioRad). qPCR analyses in cells or tissues are relative to β-actin. The primer sequences are listed in Supplementary Table S1.
Mitochondrial isolation
Cells were washed with prechilled PBS, harvested with isolation buffer (20 mM HEPES, pH 7.4, 140 mM KCl, 10 mM EDTA, 5 mM MgCl2 with a protease and phosphatase inhibitor cocktail), and homogenized using a chilled glass-Teflon homogenizer with 20 strokes. The homogenate was centrifuged twice at 700× g for 10 min at 4 °C. The supernatant was transferred to a new tube and centrifuged at 9000× g for 30 min at 4 °C to obtain the mitochondrial pellet. The pellet was collected and washed thrice with washing buffer (20 mM HEPES, pH 7.4, 800 mM KCl, 10 mM EDTA, and 5 mM MgCl2).
Histone extraction
Cells were washed twice with PBS and resuspended in 200 μL of buffer A (10 mM HEPES (pH 7.9), 10 mM KCl, 1.5 mM MgCl2, 0.34 M sucrose, 10% glycerol, 1 mM DTT, 0.1% Triton X-100, and protease inhibitor mixture (Roche Molecular Biochemicals). After incubation for 5 min on ice, the nuclei were collected in the pellet by low-speed centrifugation (1500× g, 4 min, 4 °C). The nuclei were washed once with buffer A without 0.1% Triton X-100 and then lysed in 200 μL of buffer B (3 mM EDTA, 0.2 mM EGTA, 1 mM DTT, and protease inhibitor mixture). After 10 min incubation on ice, soluble histones were separated from chromatin by centrifugation (2000× g, 4 min).
RNA interference
Small RNA interference and stable knockdown were used in this study. In RNA interference, double-stranded siRNAs targeting GDH and BDH1, respectively, were purchased from GenePharma and transfected into cells via RNAiMax (Invitrogen) according to the manufacturer’s instructions. siRNA transfections were performed with Lipo2000 (Life technology) according to the manufacturer’s instruction. Cells were cultured in six-well plate (ThermoFisher Scientific) and siRNAs were used at a concentration of 20 nM. Cells were harvested at 48 h post-transfection.
Stable shRNA knockdown cells were generated by co-transfecting cells with pCMV-VSV-G, pCMV-Gag-Pol, and shRNA plasmids by the calcium phosphate method. DMEM containing 10% FBS was used to culture the cells for 6 h after the transfection. After 24 h of transfection, supernatant of the cultured medium was collected and used as retrovirus preparation to infect cells at a density of 10% confluence in 90 mm dishes. Cells were re-infected 48 h after the initial infection using 5 μg/mL puromycin (Amresco) for selecting cells. All the sequences of siRNA/shRNA are listed in Supplementary Table S1. The knock-down efficiency was verified by qPCR or western blotting assay.
Measurement of GDH enzyme activity
Fresh tissues were weighed and transferred to solubilization buffer (1% Triton X-100, 150 mM NaCl, 10 mM KCl, 5 mM EDTA, 50 mM Tris-HCl, pH 8.0) containing protease and phosphatase inhibitors in 5 µL of extraction buffer per mg tissue and homogenized using ceramic beads. The samples were centrifuged at 14,000× g for 15 min (4 °C). The supernatant was transferred to a new tube for immunoprecipitation and enzymatic activity measurements, and protein concentrations were determined using a BCA protein assay kit (Beyotime).
The enzyme activity of GDH were determined by using a Glutamate Dehydrogenase Activity Assay Kit (Sigma). In short, the GDH protein was mixed with the assay buffer and incubated for 2 min at 37 °C in darkness. The change in absorbance at 450 nm was measured every 30 s for 10 min at 37 °C in a SpectraMax M5 Microplate Reader (Molecular Devices). Each sample was analyzed in triplicate, and the enzyme activity of GDH was calculated following the manufacturer’s instruction.
In vitro GAD1 enzymatic assay
Pre-warmed (37 °C) assay buffer (50 mM KPO4 pH 8.0, 0.2 mM PLP, 1 mM AET, 100 μg/mL BSA) containing 20 mM sodium glutamate was added to test tube containing the different cell lysates in ratio 1:1. After 120 min of incubation at 37 °C, samples were immediately placed in ice to stop the reaction and GABA was immediately quantified. Each determination was performed in triplicates and the whole experiments were repeated twice.
BHB treatment and measurement
Mice were administered with BHB esters (HVMN), 200 µL by oral gavage daily, 80 mg per mouse.
Levels of BHB in serum and brain were determined by using a BHB Assay Kit (Sigma) according to the manufacturer’s instruction. In short, the samples were mixed with the assay buffer and incubated the reaction for 30 min at room temperature in darkness. Absorbance was measured at 450 nm (A450) in a SpectraMax M5 Microplate Reader (Molecular Devices). Each sample was analyzed in triplicate.
Measurements of the concentrations of AcAc and acetone
Serum and brain AcAc levels were measured using the Ketone Body Assay Kit (Abnova, KA1630) according to the manufacturer’s instructions. The acetone concentrations were measured using the Goat Acetone Elisa Kit (MyBioSource, MBS9310393) according to the manufacturer’s instructions.
Immunofluorescence analysis
Animals were deeply anesthetized with vaporized sevoflurane and perfused with 50 mL saline, followed by 50 mL 4% PFA. The brains were extracted and soaked in 4% PFA at 4 °C overnight, cryoprotected by transferring to 30% sucrose solution until the brains were saturated. Coronal brain sections (40 µm) were cut using a freezing microtome (CM1950, Leica). The slices undergoing immunohistochemical staining were washed in PBS three times (5 min each time) and pre-incubated in PBS containing 5% normal goat serum and 0.3% Triton X-100 for 1 h at room temperature, before incubating with the following primary antibodies at 4 °C overnight: rabbit anti-Cre recombinase (1:1000, 15036, Cell Signaling, USA). Then, the sections were washed in PBS three times (5 min each time) and incubated with a secondary antibody at 37 °C for 1 h. The slices were washed in PBS thrice (5 min each time). Finally, all of the slices were mounted onto glass slides, dried, and covered with Antifade Mounting Medium (Solarbio, S2100, China).
Quantification and statistical analysis
Statistical tests were performed using GraphPad Prism 8 software and two-tailed unpaired Student’s t-test was used to compare two groups. All data are presented as the means ± standard deviation (SD). P values were calculated assuming a normal distribution and were corrected for multiple hypotheses using the Benjamini–Hochberg procedure. P values are denoted in figures as: ns, P > 0.05; *P < 0.05; **P < 0.01; ***P < 0.001; ****P < 0.0001.
Change history
30 April 2024
A Correction to this paper has been published: https://doi.org/10.1038/s41421-024-00669-w
References
Luo, J., Wang, J.-K. & Song, B.-L. Lowering low-densitytein cholesterol: from mechanisms to therapies. Life Metab. 1, 25–38 (2022).
Dyńka, D., Kowalcze, K. & Paziewska, A. The role of ketogenic diet in the treatment of neurological diseases. Nutrients 14, 5003 (2022).
Ting, R., Dugre, N., Allan, G. M. & Lindblad, A. J. Ketogenic diet for weight loss. Can. Fam. Physician 64, 906–906 (2018).
Casanueva, F. F. et al. Ketogenic diets as treatment of obesity and type 2 diabetes mellitus. Rev. Endocr. Metab. Disord. 21, 381–397 (2020).
Yancy, W. S. Jr, Olsen, M. K., Guyton, J. R., Bakst, R. P. & Westman, E. C. A low-carbohydrate, ketogenic diet versus a low-fat diet to treat obesity and hyperlipidemia: a randomized, controlled trial. Ann. Intern. Med. 140, 769–777 (2004).
Puchalska, P. & Crawford, P. A. Multi-dimensional roles of ketone bodies in fuel metabolism, signaling, and therapeutics. Cell Metab. 25, 262–284 (2017).
Wang, X.-Y. et al. The neonatal ketone body is important for primordial follicle pool formation and regulates ovarian ageing in mice. Life Metab. 1, 149–160 (2022).
Evans, J. R. et al. The natural history of treated Parkinson’s disease in an incident, community based cohort. J. Neurol. Neurosurg. Psychiatry 82, 1112–1118 (2011).
Croteau, E. et al. Ketogenic medium chain triglycerides increase brain energy metabolism in Alzheimer’s disease. J. Alzheimers Dis. 64, 551–561 (2018).
Prabowo, A. S. et al. Fetal brain lesions in tuberous sclerosis complex: TORC1 activation and inflammation. Brain Pathol. 23, 45–59 (2013).
Bock, M., Karber, M. & Kuhn, H. Ketogenic diets attenuate cyclooxygenase and lipoxygenase gene expression in multiple sclerosis. EBioMedicine 36, 293–303 (2018).
Barrea, L. et al. Is there a relationship between the ketogenic diet and sleep disorders? Int. J. Food Sci. Nutr. 73, 285–295 (2022).
Castro, K., Baronio, D., Perry, I. S., Riesgo, R. D. S. & Gottfried, C. The effect of ketogenic diet in an animal model of autism induced by prenatal exposure to valproic acid. Nutr. Neurosci. 20, 343–350 (2017).
Stafford, P. et al. The ketogenic diet reverses gene expression patterns and reduces reactive oxygen species levels when used as an adjuvant therapy for glioma. Nutr. Metab. 7, 74 (2010).
Bigelow, M. D. & Kouzani, A. Z. Neural stimulation systems for the control of refractory epilepsy: a review. J. Neuroeng. Rehabil. 16, 126 (2019).
Wells, J., Swaminathan, A., Paseka, J. & Hanson, C. Efficacy and safety of a ketogenic diet in children and adolescents with refractory epilepsy—a review. Nutrients 12, 1809 (2020).
López González, F. J. et al. Drug-resistant epilepsy: definition and treatment alternatives. Neurologia 30, 439–446 (2015).
Matsushita, Y. et al. Hyperactive mTOR signals in the proopiomelanocortin-expressing hippocampal neurons cause age-dependent epilepsy and premature death in mice. Sci. Rep. 6, 22991 (2016).
Stein, V. & Nicoll, R. A. GABA generates excitement. Neuron 37, 375–378 (2003).
Akyuz, E. et al. Revisiting the role of neurotransmitters in epilepsy: an updated review. Life Sci. 265, 118826 (2021).
Yogeswara, I. B. A., Maneerat, S. & Haltrich, D. Glutamate decarboxylase from lactic acid bacteria—a key enzyme in GABA synthesis. Microorganisms 8, 1923 (2020).
Cohen, I., Navarro, V., Clemenceau, S., Baulac, M. & Miles, R. On the origin of interictal activity in human temporal lobe epilepsy in vitro. Science 298, 1418–1421 (2002).
Karaca, M. et al. GDH-dependent glutamate oxidation in the brain dictates peripheral energy substrate distribution. Cell Rep. 13, 365–375 (2015).
Gordon, J. A. Testing the glutamate hypothesis of schizophrenia. Nat. Neurosci. 13, 2–4 (2010).
Juge, N. et al. Metabolic control of vesicular glutamate transport and release. Neuron 68, 99–112 (2010).
Achanta, L. B. & Rae, C. D. β-hydroxybutyrate in the brain: one molecule, multiple mechanisms. Neurochem. Res. 42, 35–49 (2017).
Tanner, G. R., Lutas, A., Martínez-François, J. R. & Yellen, G. Single K. ATP channel opening in response to action potential firing in mouse dentate granule neurons. J. Neurosci. 31, 8689–8696 (2011).
Yu, X. et al. GABA and glutamate neurons in the VTA regulate sleep and wakefulness. Nat. Neurosci. 22, 106–119 (2019).
Hertz, L., Peng, L. & Dienel, G. A. Energy metabolism in astrocytes: high rate of oxidative metabolism and spatiotemporal dependence on glycolysis/glycogenolysis. J. Cereb. Blood Flow Metab. 27, 219–249 (2007).
Haigis, M. C. et al. SIRT4 inhibits glutamate dehydrogenase and opposes the effects of calorie restriction in pancreatic beta cells. Cell 126, 941–954 (2006).
Hu, S. H. et al. Amino acids downregulate SIRT4 to detoxify ammonia through the urea cycle. Nat. Metab. 5, 626–641 (2023).
Yu, L., Yang, J., Yu, W., Cao, J. & Li, X. Rhein attenuates PTZ‑induced epilepsy and exerts neuroprotective activity via inhibition of the TLR4-NFκB signaling pathway. Neurosci. Lett. 758, 136002 (2021).
Yuskaitis, C. J. et al. Factors influencing the acute pentylenetetrazole-induced seizure paradigm and a literature review. Ann. Clin. Transl. Neurol. 8, 1388–1397 (2021).
Yu, F. H. et al. Reduced sodium current in GABAergic interneurons in a mouse model of severe myoclonic epilepsy in infancy. Nat. Neurosci. 9, 1142–1149 (2006).
Lüttjohann, A., Fabene, P. F. & van Luijtelaar, G. A revised Racine’s scale for PTZ-induced seizures in rats. Physiol. Behav. 98, 579–586 (2009).
Bozzi, Y., Provenzano, G. & Casarosa, S. Neurobiological bases of autism-epilepsy comorbidity: a focus on excitation/inhibition imbalance. Eur. J. Neurosci. 47, 534–548 (2018).
Li, K. & Xu, E. The role and the mechanism of gamma-aminobutyric acid during central nervous system development. Neurosci. Bull. 24, 195–200 (2008).
Modi, J. P., Prentice, H. & Wu, J. Y. Regulation of GABA neurotransmission by glutamic acid decarboxylase (GAD). Curr. Pharm. Des. 21, 4939–4942 (2015).
Sandermann, H. Jr, McIntyre, J. O. & Fleischer, S. Site-site interaction in the phospholipid activation of D-beta-hydroxybutyratede hydrogenase. J. Biol. Chem. 261, 6201–6208 (1986).
Shimazu, T. et al. Suppression of oxidative stress by β-hydroxybutyrate, an endogenous histone deacetylase inhibitor. Science 339, 211–214 (2013).
Wang, M., Chen, Z. & Zhang, Y. CBP/p300 and HDAC activities regulate H3K27 acetylation dynamics and zygotic genome activation in mouse preimplantation embryos. EMBO J. 41, e112012 (2022).
Schnepp, P. M. et al. GAD1 upregulation programs aggressive features of cancer cell metabolism in the brain metastatic microenvironment. Cancer Res. 77, 2844–2856 (2017).
Fernandez-Marcos, P. J. & Serrano, M. Sirt4: the glutamine gatekeeper. Cancer Cell 23, 427–428 (2013).
Karaca, M., Frigerio, F. & Maechler, P. From pancreatic islets to central nervous system, the importance of glutamate dehydrogenase for the control of energy homeostasis. Neurochem. Int. 59, 510–517 (2011).
Ramponi, G. & Grisolia, S. Acylation of lysine and of arginine-rich histones with carbamyl phosphate and 1,3 diphosphoglycerate. Biochem. Biophys. Res. Commun. 38, 1056–1063 (1970).
Kollipara, L. & Zahedi, R. P. Protein carbamylation: In vivo modification or in vitro artefact? Proteomics 13, 941–944 (2013).
Li, C. et al. Green tea polyphenols modulate insulin secretion by inhibiting glutamate dehydrogenase. J. Biol. Chem. 281, 10214–10221 (2006).
Yin, X. et al. Targeting glutamine metabolism in hepatic stellate cells alleviates liver fibrosis. Cell Death Dis. 13, 955 (2022).
Sánchez Fernández, I. & Loddenkemper, T. Subunit composition of neurotransmitter receptors in the immature and in the epileptic brain. Biomed. Res. Int. 2014, 301950 (2014).
Bamford, N. S., Wightman, R. M. & Sulzer, D. Dopamine’s effects on corticostriatal synapses during reward-based behaviors. Neuron 97, 494–510 (2018).
Koon, A. C. et al. Autoregulatory and paracrine control of synaptic and behavioral plasticity by octopaminergic signaling. Nat. Neurosci. 14, 190–199 (2011).
Zhao, Z. D. et al. Zona incerta GABAergic neurons integrate prey-related sensory signals and induce an appetitive drive to promote hunting. Nat. Neurosci. 22, 921–932 (2019).
Xu, S. et al. Ketogenic diets inhibit mitochondrial biogenesis and induce cardiac fibrosis. Signal Transduct. Target. Ther. 6, 54 (2021).
Kruszewska, J., Cudnoch-Jedrzejewska, A. & Czarzasta, K. Remodeling and fibrosis of the cardiac muscle in the course of obesity-pathogenesis and involvement of the extracellular matrix. Int. J. Mol. Sci. 23, 4195 (2022).
Rowland, E. A. et al. Sirtuin lipoamidase activity is conserved in bacteria as a regulator of metabolic enzyme complexes. mBio 8, e01096–17 (2017).
Anderson, K. A. et al. SIRT4 is a lysine deacylase that controls leucine metabolism and insulin secretion. Cell Metab. 25, 838–855.e15 (2017).
Liu, H. et al. Ketogenic diet for treatment of intractable epilepsy in adults: a meta-analysis of observational studies. Epilepsia Open 3, 9–17 (2018).
Neal, E. G. et al. The ketogenic diet for the treatment of childhood epilepsy: a randomised controlled trial. Lancet Neurol. 7, 500–506 (2008).
Cai, Q. Y. et al. Safety and tolerability of the ketogenic diet used for the treatment of refractory childhood epilepsy: a systematic review of published prospective studies. World J. Pediatr. 13, 528–536 (2017).
van der Louw, E. et al. Ketogenic diet guidelines for infants with refractory epilepsy. Eur. J. Paediatr. Neurol. 20, 798–809 (2016).
Krauss, R. M. Lipoprotein subfractions and cardiovascular disease risk. Curr. Opin. Lipidol. 21, 305–311 (2010).
Sampath, A., Kossoff, E. H., Furth, S. L., Pyzik, P. L. & Vining, E. P. Kidney stones and the ketogenic diet: risk factors and prevention. J. Child Neurol. 22, 375–378 (2007).
Ferraris, C. et al. Impact of the ketogenic diet on linear growth in children: a single-center retrospective analysis of 34 cases. Nutrients 11, 1442 (2019).
Wiśniewski, J. R., Zougman, A., Nagaraj, N. & Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 6, 359–362 (2009).
Zhang, X. et al. Molecular basis for hierarchical histone de-β-hydroxybutyrylation by SIRT3. Cell Discov. 5, 35 (2019).
Acknowledgements
This work was supported by grants from the State Key Development Programs of China (2018YFA0800300 (S.M.Z.), 2018YFA0801300 (W.X.), 2020YFA0803601 (J.Y.Z.), 2019YFA0801900 (J.Y.Z.)), the National Science Foundation of China (92253305 (S.M.Z.), 31821002 (S.M.Z.), 32230054 (S.M.Z.), 31930062 (S.M.Z.), 91857000 (S.M.Z.), 92157001 (S.M.Z.), 32171298 (W.X.), 81971449 (Y.Y.Y.), 82171672 (Y.Y.Y.)), Program of Shanghai Academic Research Leader (21XD1423000 (W.X.)).
Author information
Authors and Affiliations
Contributions
S.M.Z. conceived the concept, designed and supervised the experiments; J.Y.Z., Y.Y.Y., W.X., J.C., and P.C.L. supervised experiments; Y.N.Q., S.H.H., Y.X.Y., Z.Z.M., and Y. Lin performed the biological experiments; L.L. performed mice behaviour experiments; L.H. and Y.P.A. performed proteomic and metabolic experiments; S.M.Z. and J.Y.Z. wrote the manuscript. All authors read and discussed the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Qiao, YN., Li, L., Hu, SH. et al. Ketogenic diet-produced β-hydroxybutyric acid accumulates brain GABA and increases GABA/glutamate ratio to inhibit epilepsy. Cell Discov 10, 17 (2024). https://doi.org/10.1038/s41421-023-00636-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41421-023-00636-x
