
Abstract
Mitochondrial Lon is a multi-function matrix protease with chaperone activity. However, little literature has been undertaken into detailed investigations on how Lon regulates apoptosis through its chaperone activity. Accumulating evidences indicate that various stresses induce transportation of p53 to mitochondria and activate apoptosis in a transcription-independent manner. Here we found that increased Lon interacts with p53 in mitochondrial matrix and restrains the apoptosis induced by p53 under oxidative stress by rescuing the loss of mitochondrial membrane potential (Δψm) and the release of cytochrome C and SMAC/Diablo. Increased chaperone Lon hampers the transcription-dependent apoptotic function of p53 by reducing the mRNA expression of p53 target genes. The ATPase mutant (K529R) of chaperone Lon decreases the interaction with p53 and fails to inhibit apoptosis. Furthermore, the chaperone activity of Lon is important for mitochondrial p53 accumulation in an mtHsp70-dependent manner, which is also important to prevent the cytosolic distribution of p53 from proteasome-dependent degradation. These results indicate that the chaperone activity of Lon is important to bind with mitochondrial p53 by which increased Lon suppresses the apoptotic function of p53 under oxidative stress. Furthermore, mitochondrial Lon-mtHsp70 increases the stability/level of p53 through trafficking and retaining p53 in mitochondrial matrix and preventing the pool of cytosolic p53 from proteasome-dependent degradation in vitro and in clinic.
Similar content being viewed by others

Introduction
The tumor-suppressor gene p53 is a key regulator of cell cycle arrest, senescence, and cell death including apoptosis and necrosis1,2,3. Thus p53 acts as one of the most important barriers against malignant development of cancer cells by linking many stress response pathways such as DNA damage, hypoxia, and oxidative stress4. A well-characterized function of p53 in the apoptosis regulation is its role as a transcriptional regulator. In addition to the functions as a transcription factor, p53 acts directly upon the outer membrane of mitochondria via a transcription-independent pathway. Upon onset of apoptosis following DNA damage stress, a part of p53 translocates to mitochondria, where it interacts with Bcl-2 or Bak, resulting in cytochrome C release and caspase-3 activation5. In addition, p53 accumulates in the mitochondrial matrix and triggers mitochondrial permeability transition pore (MPTP) opening and necrosis by interaction with the MPTP regulator cyclophilin D under oxidative stress2. However, mechanisms of p53-mediated transcription-independent apoptotic pathways in mitochondrial matrix are still lacking.
Mitochondria control cell death and survival by regulating intrinsic apoptosis, autophagy, necrosis, and ferroptosis2,6,7. Mitochondrial Lon protease is located in matrix and plays a crucial role in the maintenance of mitochondrial function, biogenesis, and homeostasis8,9. In addition to its ATP-dependent proteolytic activity, mitochondrial Lon has been found to show chaperone activity10,11,12,13. Mitochondrial Lon is a stress protein and induced by a number of stresses, such as hypoxia, oxidative, and unfolded protein stress10,12,14,15. Molecular chaperones including mitochondrial chaperones have been associated with enhanced cell survival under stress by inhibition of apoptotic cell death and increased stability of survival effectors that promote tumor growth16,17,18. Indeed, Lon downregulation causes loss of mitochondrial function, early embryonic lethality, reduced cell proliferation, and apoptosis12,19,20,21. Lon upregulation is required for cancer cell survival and tumorigenesis by regulating stress responses induced by oxidative condition12,20,22. However, the molecular mechanism of how Lon regulates apoptosis remains largely unclear. We recently identified heat-shock protein 60 (Hsp60) and mitochondrial Hsp70 (mtHsp70) as chaperone Lon clients by utilizing proteomic approach17. Interestingly, the ability of increased Lon-inhibited apoptosis is dependent on Hsp60 that binds p53 to inhibit apoptosis16,17. These findings allowed us to pursue the detailed mechanism of how chaperone Lon directly regulates apoptosis by interacting with p53.
To our knowledge, the present study for the first time demonstrates that p53 is bound by Lon in the mitochondrial matrix to control apoptosis. In this study, we demonstrated that Lon interacts with p53 in mitochondrial matrix and restrains the apoptosis induced by p53 under oxidative stress by reducing the mRNA expression of p53 target genes and rescuing the loss of mitochondrial membrane potential (Δψm) and the release of cytochrome C. The ATPase mutant (K529R) of mitochondrial Lon decreased the interaction with p53, reduced mitochondrial localization of p53, and failed to inhibit apoptosis, suggesting that the chaperone activity of Lon is important for the control of p53 protein level and apoptotic function by sequestering p53 in mitochondrial matrix. In addition, the level of cytoplasmic p53 significantly correlates that of mitochondrial Lon in oral cancer patients. Thus our findings suggest that targeting the chaperone activity of mitochondrial Lon will increase the efficacy of p53-induced apoptosis in cancer therapy.
Results
Overexpression of mitochondrial Lon increases the accumulation of mitochondrial p53 and restrains p53-dependent apoptosis under oxidative stress
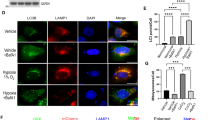
We previously showed that mitochondrial Lon physically interacts with Hsp60–mtHsp70 complex and regulates apoptosis through Hsp6017. Since Hsp60 binds p53 to restrain its apoptosis function in cytosol and mitochondria16, we asked whether Lon regulates p53-induced apoptosis under stress. We first found that the level of Lon and p53 are increased in cytosol and mitochondria after H2O2 and rotenone treatment (Fig. 1a, b, and Supplemental Figure S1). The level of cytosolic and mitochondrial p53 was further increased when Lon was overexpressed in cells (Fig. 1b) and only mitochondrial p53 was decreased when Lon was downregulated under oxidative stress (Fig. 1c), suggesting that the mitochondrial localization of p53 is regulated by Lon under oxidative stress. Since p53 protein can translocate to mitochondrial matrix in response to oxidative stress2, we tried to entangle the mitochondrial localization of p53 with Lon under oxidative stress. The immunofluorescence analysis showed that there was no significant colocalization of p53 (green) and Lon (red) in the control cells. Following exposure to H2O2, both p53 and Lon accumulated in the mitochondria, and they appeared colocalized with each other as yellow spots (Fig. 1d), which raised a possibility of the interaction between Lon and p53 in mitochondria under oxidative stress. Next we found that the signals of pro-apoptotic proteins, Bax, p53, cytochrome C release, and SMAC/Diablo release from mitochondria, are increased after H2O2 treatment. However, the activation of apoptosis was almost inhibited when Lon was overexpressed including the reduction of cytochrome C release (Fig. 1b). The results suggest that increased Lon restrains p53-induced apoptosis during oxidative stress. To confirm the role of Lon in p53-mediated apoptosis, terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay was performed. The TUNEL-positive cells were detected more when the cells were overexpressed by p53 and treated with H2O2, compared to the vector control cells (a, b, and d in Fig. 1e). However, the TUNEL-positive signals in p53-overexpressed cells and the H2O2-treated cells were decreased when Lon was also overexpressed (c and e in Fig. 1e). We further found that the positive signals repressed by Lon overexpression were reversed when p53 was overexpressed (f in Fig. 1e). Consistently, the TUNEL-positive signals induced by decreased Lon were rescued when p53 was knocked down under H2O2 treatment (g in Fig. 1e). The mitochondrial membrane potential (MMP) analysis showed that MMP was decreased in p53-overexpressed cells, compared to the vector control cells, and the MMP in p53-overexpressed cells was rescued when Lon was overexpressed (Fig. 1f). These data showed that increased Lon protein restrains p53-dependent apoptosis in cancer cells under oxidative stress, in which Lon is required for mitochondrial p53 accumulation.

a Immunoblotting analysis of the increase of p53 and Lon in the mitochondrial fraction under oxidative stress. HSC3 cells were treated with 200 μM H2O2 for 4 h. Immunoblotting were performed using the indicated antibodies. The purity of each cell fractions was monitored by immunoblotting for cytoplasmic (Actin) and mitochondrial (VDAC) markers. b Increased mitochondrial Lon restrains p53-dependent apoptosis in cancer cells under oxidative stress. HSC3 cells transfected with or without pcDNA3-Myc-Lon were treated with 200 μM H2O2 for 4 h, then recovered for 4 h. Whole-cell lysates were used to purify the mitochondrial and cytosolic fractions. Apoptosis-associated proteins were detected by western blot analysis using the indicated antibodies. The purity of mitochondrial fraction was monitored by immunoblotting for mitochondrial (VDAC) markers, and anti-Actin and anti-VDAC antibodies were used as loading control. c Mitochondrial Lon is required for mitochondrial accumulation of p53 under oxidative stress. HSC3 cells were transfected with the plasmid encoding Lon-shRNA. The transfected cells were treated with 200 μM H2O2 for 4 h. Immunoblotting were performed using the indicated antibodies. The purity of each cell fractions was monitored by immunoblotting for cytoplasmic (Actin), lysosome (LAMP2), and mitochondrial (VDAC and COX-4) markers. d The accumulation of mitochondrial Lon and p53 in mitochondria under oxidative stress was verified by immunofluorescence. 293T cells were untreated or treated with 50 μM H2O2 for 24 h. The cells were fixed and immunostained by anti-p53 (green) and anti-Lon (red) antibodies. DNA was stained with DAPI (blue). Scale bar, 10 μm. e Increased mitochondrial Lon restrains p53-dependent apoptosis under oxidative stress shown by TUNEL assay. The apoptosis in HSC3 cells was induced by p53 overexpression or by treatment with 200 μM H2O2 for 4 h. The treated HSC3 cells were transfected with pcDNA3-Myc-Lon, Lon-shRNA, p53-siRNA or not. Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL) assay was applied to examine the effect of Lon overexpression on p53-induced apoptosis. TUNEL-positive cells (green fluorescence) were counted in the cells expressing vector (a), p53 overexpression (b), or Lon plus p53 (c). Treatment with H2O2 acted as a positive control (d, e). Treatment with H2O2 plus Lon and p53 (f) and with H2O2 plus Lon-shRNA and p53-siRNA (g). DAPI was used for nuclear staining. Scale bar, 20 μm. The error bars shown in the bottom panel represent the standard deviation from three different experiments. *p < 0.05 and **p < 0.01. f Increased mitochondrial Lon restrains p53-dependent apoptosis shown by mitochondrial membrane potential assay. The apoptosis in HSC3 cells was induced by p53 overexpression. The HSC3 cells were transfected with pcDNA3-Myc-Lon plasmid or not. Mitochondrial membrane potential assay was applied to examine the effect of Lon overexpression on p53-induced apoptosis. The transfected cells were stained with 50 nM DiOC6(3) for 30 min and analyzed with flow cytometer. FCCP treatment acts as a control of mitochondrial membrane potential which shows that the stain is fundamentally dependent on mitochondrial membrane potential. The error bars shown in the bottom panel represent the standard deviation from three different experiments. *p < 0.05
Chaperone Lon interacts with p53 in mitochondrial fraction under oxidative stress
Since Lon physically interacts with Hsp60–mtHsp70 to restrain its apoptosis function in mitochondria, we asked whether p53 is a direct client protein of chaperone Lon. The association between Lon and p53 was first examined by co-immunoprecipitation (Co-IP) experiment (Fig. 2a, b), and endogenous p53 was able to be co-immunoprecipitated with Lon under oxidative stress (Fig. 2c), suggesting that mitochondrial Lon interacts with p53 in vivo. To further confirm the finding, we performed Co-IP experiment using the isolated mitochondrial fraction. Consistently, the expression of p53 is increased in mitochondrial fraction when Lon is overexpressed; overexpression of p53 triggers its accumulation in mitochondrial fraction (Fig. 2d, left panel). Lon was able to be co-immunoprecipitated with p53 in mitochondrial fraction and vice versa (Fig. 2d, right panel). Furthermore, the His-tag and GST-tag pull-down assays confirmed a direct interaction between Lon and p53 in vitro (Fig. 2e). These data demonstrate that mitochondrial chaperone Lon interacts with p53 under oxidative stress.

a, b Lon interacts with p53 shown by co-immunoprecipitation. 293T (a) cells were transiently transfected with the plasmids encoding Myc-Lon and p53 followed by co-immunoprecipitation with anti-Myc and anti-p53, respectively. Whole-cell lysates from HSC3 (b) cells transfected with the plasmids encoding Myc-Lon or p53 were immunoprecipitated with anti-p53 or anti-Lon antibodies. The immunoprecipitation complex was analyzed by western blotting using the indicated antibodies. IP immunoprecipitation. c Lon interacts with endogenous p53 under oxidative stress shown by co-immunoprecipitation. HSC3 cells were treated with 2 mM H2O2 for 6 h or 2 μM rotenone for 6 h. Whole-cell lysates were analyzed by western blotting using the indicated antibodies (left panel). Whole-cell lysates from HSC3 cells treated with H2O2 or rotenone were immunoprecipitated with anti-p53 or anti-Lon antibodies. The immunoprecipitation complex was analyzed by western blotting using the indicated antibodies. d Lon interacts with p53 in mitochondrial fraction shown by co-immunoprecipitation. HSC3 cells were transiently transfected with the plasmids encoding Myc-Lon or p53. Whole-cell lysates from HSC3 cells were used to purify the mitochondrial and cytosolic fractions followed by co-immunoprecipitation with anti-p53 and anti-Myc, respectively. The mitochondrial fraction and the immunoprecipitation complex from transfected HSC3 cells were subjected to immunoblotting using the indicated antibodies. The purity of mitochondrial fraction was monitored by immunoblotting for mitochondrial (VDAC) markers. e Mitochondrial Lon interacts with p53 shown by the pull-down assay. Direct interaction between mitochondrial Lon and p53 was verified by His-tag and GST-tag pull-down assay. The His-Lon fusion protein and GST-p53 were added, and the His-tag proteins were pulled down by anti-His antibody and the GST-tag proteins were pulled down by anti-GST antibody. The pulled down complex was subjected to immunoblotting using the indicated antibodies. The His-tag/GST-tag proteins were pulled down by anti-His/anti-GST antibody as a positive control. The His- and GST-fusion proteins added were shown on SDS/PAGE stained with Coomassie brilliant blue (right panel)
ATPase activity of Lon is required for the interaction with p53 by which attenuates apoptosis in mitochondria
To explore that the chaperone activity of Lon is required for the interaction with p53, we used the ATPase mutant of Lon, Lon-K529R23, to examine whether the ATPase activity is critical to the interaction with p53. The association between Lon and p53 was examined by Co-IP experiment. The result showed that the Lon-K529R mutant significantly abolished the interaction with p53 but not in the protease mutant, Lon-S855A (Fig. 3a). Consistently, endogenous p53 was able to be co-immunoprecipitated with Lon-WT under rotenone treatment but not the Lon-K529R mutant (Fig. 3b), suggesting that the ATPase activity of mitochondrial Lon is required for the binding with p53. To confirm the chaperone activity of Lon is important in p53-mediated apoptosis regulation, TUNEL assay was performed by using the overexpression of Lon-K529R mutant. The TUNEL-positive cells in p53-overexpressed cells were significantly decreased when Lon was also overexpressed. However, the Lon-K529R mutant failed to inhibit p53-induced apoptosis. Similarly, the Lon-K529R mutant failed to inhibit H2O2-induced apoptosis (Fig. 3c). However, the TUNEL-positive signals were reduced when p53 was knocked down under H2O2 treatment and the positive signals induced by the overexpression of Lon-K529R mutant were rescued when p53 was knocked down under H2O2 treatment (Fig. 3c). These data showed that mitochondrial Lon interacts with p53 through its chaperone activity that is required to restrain p53-dependent apoptosis in cancer cells under oxidative stress.

a The K529 residue of mitochondrial Lon is required for the interaction with p53. Whole-cell lysates from HSC3 cells transfected with the plasmids encoding p53 and wild-type pcDNA3-Myc-Lon (Lon-WT), an ATPase mutant (LonK529R), a proteolytic mutant (LonS855A), or an ATPase/proteolytic mutant (LonK529R/LonS855A) were immunoprecipitated with anti-Myc antibodies. The immunoprecipitation complex was analyzed by immunoblotting using the indicated antibodies. b The K529 residue of mitochondrial Lon is required for the interaction with endogenous p53 under oxidative stress. Whole-cell lysates from HSC3 cells transfected with the plasmids encoding Lon-WT and LonK529R mutant were analyzed by western blotting using the indicated antibodies (left panel). Whole-cell lysates from HSC3 cells transfected with the plasmids encoding Lon-WT and LonK529R mutant under rotenone treatment (10 μM for 8 h) were immunoprecipitated with anti-Myc-Lon antibodies. The immunoprecipitation complex was analyzed by western blotting using the indicated antibodies. c K529 residue of mitochondrial Lon is required for the inhibition toward p53-dependent apoptosis under oxidative stress shown by TUNEL assay. Apoptosis in HSC3 cells was induced by p53 overexpression. HSC3 cells were transfected with sip53 and/or the plasmids encoding p53, wild-type pcDNA3-Myc-Lon, or an ATPase mutant of Lon (LonK529R). TUNEL assay was applied to examine the effect of the K529 residue of mitochondrial Lon on p53-induced apoptosis under oxidative stress (200 μM H2O2 for 4 h). TUNEL-positive cells (green fluorescence) were counted in the transfected cells. DAPI was used for nuclear staining. Scale bar, 20 μm. The error bars shown in the right panel represent the standard deviation from three different experiments. **p < 0.01
Increased chaperone Lon hampers the transcription-dependent apoptotic function of p53 under oxidative stress by retaining p53 in the mitochondria
Since p53 is known for its ability to orchestrate cell cycle and apoptosis by a transcription-dependent mechanism24,25, we examined whether the suppression of p53-dependent apoptosis by mitochondrial Lon is through affecting the distribution between nuclear and mitochondrial p53. Thus we checked whether the transcription-dependent function of nuclear p53 is affected by increased chaperone Lon. The mRNA expression of Lon gene was significantly increased when Lon was overexpressed (Fig. 4a). The mRNA expression of p53 gene was significantly increased when the cells were overexpressed by p53 and treated with H2O2, but not in Lon-overexpressed and p53-knocked-down cells (Fig. 4b). This result indicates that Lon overexpression is not able to affect p53 gene expression. Then we checked the expression of several p53-targeted genes, such as Puma/Bim in the intrinsic apoptosis pathway24, Fas that is involved in the extrinsic pathway25, and p53R2 that is required for mitochondrial DNA stability and cell protection from oxidative stress26,27.

a, b The mRNA expression of Lon and p53 was analyzed by quantitative real-time PCR. c HSC3 cells were transfected with the plasmids encoding wild-type pcDNA3-Myc-Lon, a proteolytic mutant (LonS855A), or an ATPase mutant of Lon (LonK529R) under oxidative stress (200 μM H2O2 for 4 h). The mRNA expression of p53-targeted genes, Bim, Fas, and p53R2, was analyzed by quantitative real-time PCR. The results were presented as fold increase relative to vector-transfected cells (deliberately set to 1). Data are presented as mean ± SD of at least three independent experiments. The error bars shown in the panel represent the standard deviation from three different experiments. d HSC3 cells were transfected with the plasmids encoding p53 and/or wild-type pcDNA3-Myc-Lon or an ATPase mutant of Lon (LonK529R). The mRNA expression of p53-targeted genes, Puma, Bim, Fas, and p53R2, was analyzed by quantitative real-time PCR. The results were presented as fold increase relative to vector-transfected cells (deliberately set to 1). Data are presented as mean ± SD of at least three independent experiments. The error bars shown in the panel represent the standard deviation from three different experiments. *p < 0.05 and **p < 0.01.
The result showed that increased Lon-WT and the Lon-S855A mutant reduce the induction of p53-dependent apoptotic genes under oxidative stress. However, the overexpression of Lon-K529R mutant increased the expression of p53-dependent apoptotic genes under the same condition (Fig. 4c). Consistently, increased mitochondrial Lon reduced the induction of p53-dependent apoptotic genes when p53 was overexpressed. However, the Lon-K529R mutant failed to largely inhibit the expression of p53-targeted genes (Fig. 4d and Supplemental Figure S2). These results indicate that the ATPase activity of chaperone Lon indeed is important for p53-dependent apoptosis; the mechanism of apoptotic inhibition by increased Lon under oxidative stress is associated with sequestering some p53 in mitochondria that also lowers the nuclear distribution of p53.
Chaperone Lon-mtHsp70 is required for mitochondrial p53 accumulation that is important to prevent the cytosolic distribution of p53 from proteasome-dependent degradation
We found that the levels of p53 in cytosol and mitochondria were further increased when Lon was overexpressed in cells under oxidative stress (Fig. 1b), and the level of p53 in mitochondria was decreased when Lon was downregulated but not the level in cytosol under oxidative stress (Fig. 1c), suggesting that the mitochondrial accumulation and level of p53 is regulated by increased Lon under oxidative stress. Since previous findings showed that overexpression and knocked down of Lon both induce the production of ROS12,28, we tried to understand the mechanism of p53 accumulation induced by increased Lon without H2O2 treatment. Consistently, we observed that the protein level of p53 was increased when Lon was overexpressed in cancer cells (Fig. 5a), and knocked down of Lon caused a decrease in p53 level (Fig. 5b). We confirmed that p53 was accumulated in mitochondrial fraction when p53 was overexpressed in cells (Fig. 5c). p53 was increased in cytosol and mitochondrial fraction when Lon was overexpressed (Fig. 5d, left panel), and the level in mitochondria was decreased when Lon expression was inhibited by short hairpin RNA (shRNA; Fig. 5d, right panel). Intriguingly, we found that p53 still remained in the cytosol fraction when Lon was knocked down (Fig. 5d, right panel). These data suggest that Lon may be involved in the stability of p53 in the mitochondrial matrix, which may affect the distribution or transportation of p53 between cytosol and mitochondria. To test this idea, we knocked down mtHsp70 (mortalin) to examine the mechanism of p53 accumulation induced by increased Lon because mtHsp70 is involved in both protein import and folding process in mitochondrial protein homeostasis and is recently identified as a chaperone client of Lon17. We confirmed that p53 was increased in cytosol and mitochondrial fraction when Lon was overexpressed and the levels of p53 and mtHsp70 in mitochondria were decreased when Lon was knocked down by shRNA (Fig. 5e). However, the mitochondrial p53 accumulation induced by Lon overexpression was diminished when mtHsp70 was knocked down, suggesting that mtHsp70 is required for p53 accumulation in the mitochondria when Lon was overexpressed (Fig. 5e). Since we found that Lon-mtHsp70 is required for p53 accumulation in the mitochondria but not for the p53 level in the cytosol, we next asked whether the level of mitochondrial Lon regulates the distribution between cytosolic and mitochondrial p53. To answer this question, we first treated the Lon-knocked-down cells with a proteasome inhibitor, MG132, the level of p53 was gradually recovered, and increased in the cells (Supplemental Figure S3), indicating that p53 level regulated by Lon is mediated by the prevention of proteasome-dependent degradation in cytoplasm. Under MG132 treatment, p53 was increased in both cytosolic and mitochondrial fraction when Lon was overexpressed (revised Fig. 5f), suggesting that cytosolic and mitochondrial p53 are both affected by cytosolic proteasome-dependent degradation. Consistently, p53 was accumulated in the cytosol fraction under MG132 treatment when Lon, Lon-shRNA, or Lon-K529R mutant was overexpressed (Fig. 5f, left panel). However, p53 was decreased in the mitochondrial fraction when Lon was knocked down by shRNA or overexpression of the Lon-K529R mutant even under MG132 treatment (Fig. 5f, right panel). These results indicate that the chaperone activity of mitochondrial Lon and mtHsp70 are important to accumulate p53 in the mitochondria and to prevent the cytosolic distribution of p53 from proteasome-dependent degradation.

a, b Mitochondrial Lon is important for the stability/level of p53 protein in cancer cells. For overexpression experiment, oral cancer cells were transfected with the plasmid encoding Myc-tagged Lon. For knocking down experiment, Lon expression was inhibited by Lon-shRNA transfection. Immunoblotting were performed using the indicated antibodies. c Immunoblotting analysis of increased p53 in mitochondrial fraction. p53 was overexpressed in OEC-M1 cells by transfected with the plasmid encoding p53. Immunoblotting were performed using the indicated antibodies. The purity of each cell fractions was monitored by immunoblotting for cytoplasmic (Actin) and mitochondrial (VDAC) markers. d Chaperone Lon is required for mitochondrial p53 accumulation. OEC-M1 cells were transfected with the plasmid encoding Myc-tagged Lon or Lon-shRNA. Immunoblotting were performed using the indicated antibodies. The purity of each cell fractions was monitored by immunoblotting for cytoplasmic (Actin) and mitochondrial (VDAC and COX-4) markers. e Chaperone Lon-induced mitochondrial p53 accumulation is dependent on mtHsp70. OEC-M1 cells were transfected with the plasmid encoding Myc-tagged Lon, Lon-shRNA, and/or mtHsp70-shRNA. Immunoblotting were performed using the indicated antibodies. The purity of each cell fractions was monitored by immunoblotting for cytoplasmic (Actin) and mitochondrial (VDAC and COX-4) markers. f The chaperone activity of Lon is required for mitochondrial p53 accumulation and the prevention of the cytosolic distribution of p53 from proteasome-dependent degradation. OEC-M1 cells were transfected with the plasmid encoding WT-Lon, Lon-shRNA, or the ATPase mutant of Lon (LonK529R). The transfected cells were treated with 10 μM MG132 for 2 h. Immunoblotting were performed using the indicated antibodies. The purity of each cell fractions was monitored by immunoblotting for cytoplasmic (Actin, left) and mitochondrial (VDAC and COX-4, right) markers
To understand the interaction between mitochondrial Lon and p53, we tried to build a complex model structure by using protein–protein docking and the information-driven approach. Indeed, the most stable complex model showed that p53 core structure binds into a prominent valley formed between the ATPase domain and alpha-subdomain of mitochondrial Lon (Fig. 6a, b). This observation is consistent with a previous study which showed that the ATP-dependent helicase domain of SV40 Large T-antigen bind to the core structure (DNA-binding domain) of p5329. Since the K529R mutant of Lon abolished the ability to hydrolyze ATP (ATPase activity) but retained the ability to bind ATP23, the conformational change of the mutant will be inhibited when ATP accessed into the ATP pocket and occupied the pocket, which may explain the steric effect of the ATPase mutant of chaperone Lon on the binding with p53. In addition, surface-charge representations of the Lon–p53 complex model structure were built (Fig. 6b, c). The structure shows that the surface-charge property in the p53-binding valley of Lon is positively charged (Fig. 6b), and the charge property of p53 in the Lon-binding patches is negatively charged (Fig. 6c). The result implies that electrostatic forces may mediate the interactions between mitochondrial Lon and p53 core structure.

a Ribbon representation of the structure of mitochondrial Lon–p53 complex. The protein structure is labeled along the sequence. Each domain is shown in a different color: ATPase domain in yellow, protease domain in blue, and p53 core domain in purple. K529 in ATPase pocket domain is indicated by arrow. b Surface-charge representation of mitochondrial Lon–p53 complex structure is shown. Basic, acidic and uncharged surface regions are colored blue, red and white, respectively. p53 core domain is represented by Ribbon in purple. c. Structure representation of the mitochondrial Lon–p53 complex. The same cartoon as b after an 180o rotation around the vertical axis. Mitochondrial Lon is represented by Ribbon and p53 core domain is represented by surface-charge model. Basic, acidic, and uncharged surface regions are colored blue, red, and white, respectively
The level of cytoplasmic p53 protein correlates the level of mitochondrial Lon in oral cancer patients
We found that total p53 level is controlled by the level of mitochondrial Lon in several cancer cell lines (Fig. 5a, b) and the mitochondrial accumulation of p53 is regulated by Lon (Fig. 5e, f), suggesting that the cytoplasmic level (includes mitochondrial fraction) of p53 protein correlates the level of mitochondrial Lon. We then examined whether p53 stability/level regulated by mitochondrial Lon is clinically relevant. First, we found that the expression of Lon protein correlates with that of p53 in seven oral cancer cell lines (Fig. 7a). To link the clinical significance of increased Lon and p53 level, 123 samples of tumor tissues from oral squamous cell carcinoma (OSCC) patients were used to determine Lon and p53 expression pattern by immunohistochemical (IHC) analysis. The main clinicopathological characteristics of the 123 patients of this study are detailed in Table S1. Mitochondrial Lon was found in the cytoplasm of cancer cells (a and b in Fig. 7b); p53 was identified in nucleus only (c in Fig. 7b) or in both nucleus and cytoplasm of cancer cells (d in Fig. 7b), which are consistent with our previous reports12,30. The high expression (IHC level, median and strong) of Lon was observed in the majority of OSCC tumor tissues (85/123, 69.1%); the high expression of p53 showed a nearly half ratio in tumor tissues (64/123, 52.0%) (Table S2). We next examined the association between Lon and p53 expression in OSCC tissues using Fisher’s exact test and measured the correlation in contingency table. The results showed that p53 expression in either nucleus/cytoplasm or nucleus only shows no significant correlation with Lon expression (P = 0.879 and P = 0.264, respectively, Table S2 and S3). However, p53 simultaneous expression in cytoplasm showed a significant correlation with Lon expression (p = 0.047, Table 1). Consistently, when the expression pattern was categorized into three groups: strong, median, and weak expression, the correlation between Lon and p53 expression in nucleus/cytoplasm is statistically significant (p = 0.05) with a Cramer’s V coefficient of 0.44 (Table S4) but not in nucleus or cytoplasm or in nucleus only (Table S5 and S6). Taken together, these result indicated that cytoplasmic p53 protein is correlated with the level of Lon protein in OSCC patients.

a The protein level of Lon and p53 in oral cancer cell lines. The extracts of oral cancer cell lines were immunoblotted with the indicated antibodies and antibody to Tubulin as a loading control. b Immunohistochemical analysis of Lon and p53 expression in OSCC patients. Representative immunohistochemical analysis of Lon and p53 was performed by using paraffin-embedded sections of OSCC. The representative results shown here are positive staining of Lon (a, 200×; b, 400×), nuclear staining of p53 (c), and nuclear/cytoplasmic staining of p53 (d) in oral cancer tissues. The microscopic magnification of p53 staining was 400×. Scale bar, 50 μm. c Model of p53 accumulation in mitochondria and apoptosis inhibition by chaperone Lon in cancer cells. Upon Lon overexpression and/or oxidative stress, mitochondrial Lon binds with p53 and induces the accumulation of p53 in the matrix through its chaperone activity (residue K529) and inhibits p53-mediated apoptosis including transcription-independent and -dependent mechanisms. Mitochondrial Lon sequesters p53 to inhibit the opening of MPTP on the outer membrane and the MPTP–cyclophilin D complex on the inner membrane. Meanwhile, mitochondrial Lon retains p53 in the matrix to reduce the transcription-dependent function of nuclear distribution of p53. The stability/level of cytosolic p53 is increased by the prevention of proteasome-dependent degradation through sequestering p53 by Lon-mtHsp70 in mitochondrial matrix
Discussion
In this study, we have shown that mitochondria Lon interacts with p53 and retains it in mitochondrial matrix to restrain the apoptosis induced by oxidative stress. The ATPase mutant of Lon decreases the interaction with p53 and fails to inhibit apoptosis under oxidative stress. This study reveals that Lon overexpression inhibits apoptosis through the chaperone activity by interacting with and sequestering p53 in mitochondria.
Molecular chaperones of HSP family play important roles in promoting cell survival and tumor growth18,31. We previously identified a number of candidate client proteins of mitochondrial chaperone Lon by using a proteomic approach. We identified NDUFS8, Hsp60, and mtHsp70 are client proteins of mitochondrial Lon12,17. Mitochondrial Lon physically interacts with Hsp60–mtHsp70 complex and the protein stability/level of Hsp60 and mtHsp70 depends on the level of Lon under oxidative stress17. Hsp60 binds p53 to restrain its apoptosis function: the depletion of Hsp60 increases the level of p53 but does not affect MDM2 level16, suggesting that there is an unknown mechanism to regulate the p53 protein level beyond nucleus and cytoplasm. Mitochondrial Hsp60 inhibits apoptosis by antagonizing cyclophilin D-dependent mitochondrial permeability transition32, increasing the stabilization of survivin and restraining p53 function16. The scenarios for the mechanism underlying Lon-mtHsp70-inhibited apoptosis through binding with and stabilizing p53 are described (Fig. 7c). We found that increased mitochondrial Lon inhibits p53-mediated apoptosis including transcription-independent and -dependent mechanism. p53-mediated cell death is involved in transcription-dependent and -independent regulation24,33,34. Regarding transcription-independent mechanism, in response to oxidative stress, the cytoplasmic pool of p53 protein localizes to the outer membrane of mitochondria where it activate transcription-independent apoptosis by physically inhibiting antiapoptotic members (Bcl2, BclxL) as well as activating proapoptotic members (Bak, Bax) of mitochondrial outer membrane permeabilization (MOMP) regulators5,34. To our knowledge, the present study for the first time demonstrates that p53 is bound by Lon in the mitochondrial matrix to control MOMP and apoptosis. Indeed, we observed that increased mitochondrial Lon rescued Δψm and the release of cytochrome C induced by p53. This finding is consistent with the observation that p53 accumulates in the mitochondrial matrix and triggers MPTP opening and necrosis by interaction with cyclophilin D under oxidative stress2. MPTP is a regulated protein channel spanning the inner and outer mitochondrial membranes. Previous reports showed that matrix Lon interacts with proteins located at inner membrane, such as prohibitin, COX, and NDUFS812,17,35,36. We suggest that Lon may serve as a recruiter complex center to trap p53 to disrupt the interaction with cyclophilin D, by which it could prevent apoptosis by inhibiting the opening of MPTP, high levels of cytosolic Ca2+, and ROS accumulation during oxidative stress.
This study indicates that the chaperone activity of mitochondrial Lon is important to control p53 protein level by translocating and sequestering p53 in mitochondria under oxidative stress. Consistently, the mutations in the AAA+ domain of Lon caused the aggregation of mtDNA-encoded cytochrome C oxidase subunit II, which reduces the function of mitochondrial respiration in CODAS syndrome patients37. Thus the possible mechanism underlying Lon-mediated stabilization of p53 will be controlling p53 translocation into mitochondria. Accumulating studies have shown that at least three different mechanisms were described by which p53 translocates into the matrix of mitochondria, including the two classical import systems utilizing mitochondrial targeting sequences (MTS) or chaperone carriers and one mechanism that involves redox/respiration-dependent import system38,39. However, no MTS were found in the N- or C-terminal domain of p53 protein. Thus it is worth attracting more attention that p53 translocates into the matrix by the mechanism of chaperone carriers. Indeed, the mitochondrial trafficking of cytosolic p53 is mediated by mtHsp70/Tid-1 complex under DNA damage and hypoxia40,41. Our previous work showed that mtHsp70/Hsp60 complex act as Lon-associated proteins and their protein level and stability are dependent on Lon17. Therefore, increased mitochondrial Lon may promote the translocation of p53 into mitochondria by stabilizing the chaperone carriers, Hsp60/mtHsp70/Tid-1 complex. Consequently, mitochondrial Lon-mtHsp70 restrains p53-dependent apoptosis under stress, including the function of nuclear p53-dependent transcription and cytosolic p53-dependent mitochondria targeting.
In summary, we identified and validated p53 as a chaperone client of Lon along with Hsp60 and mtHsp70. This study for the first time reported that the function of p53 translocated into mitochondrial matrix in apoptosis regulation. Mitochondrial Lon retains p53 in the mitochondria matrix through its chaperone activity and inhibits p53-mediated apoptosis by transcription-independent and -dependent mechanisms. We have shown that mitochondria Lon interacts with p53 and retains it in mitochondrial matrix to restrain the apoptosis induced by oxidative stress. The ATPase mutant of Lon decreases the interaction with p53 and fails to inhibit apoptosis under oxidative stress. Our studies will provide new insights into the chaperone function of Lon in apoptotic cell death exerted by directly sequestering p53 in mitochondria and will allow us to understand that targeting the chaperone activity of mitochondrial Lon increases the efficacy of p53-induced apoptosis in cancer therapy.
Materials and methods
Patients and clinical sample
Tissue specimens of 123 patients with OSCC were chosen for IHC analysis based on availability of archival human oral tissue blocks from diagnostic resection specimens in the Departments of Pathology at Mackay Memorial Hospital, Taipei, Taiwan with approval from the Institutional Review Board (IRB number: 13MMHIS188). The main clinical characteristics of the 123 patients selected for this study are detailed in Table S1. All experiments were performed in accordance with relevant guidelines and regulations. The levels of Lon and p53 protein expression are categorized into no available, low, median, and strong based on the scores of IHC staining.
Cell culture and cell treatment
293, 293T, HSC3, FADU, and DOK (dysplastic oral keratinocyte) cells were cultured in medium containing Dulbecco’s modified Eagle’s essential medium (DMEM) (GIBCO, New York, NY, USA), supplemented with 5% fetal bovine serum (FBS) and 5% super calf serum and penicillin–streptomycin (50 U/ml, Sigma, St. Louis, MO, USA) in a 5% CO2/95% air atmosphere. SCC-9, SCC-25, and SAS cells were cultured in medium containing a 1:1 mixture of DMEM/F12 medium, supplemented with 10% FBS. OEC-M1 cells were cultured in medium containing RPMI 1640 medium, supplemented with 10% FBS. Stable cell lines expressing Lon-shRNA were generated by retroviral infection using pMKO vector. Briefly, the plasmids, Lon-Sh1/2-pMKO-puro for stable knockdown cells, along with packaging plasmid gag-pol and envelope plasmid VSV-G, were transfected into 293T cells by Lipofectamine™ 2000 (Invitrogen, Carlsbad, CA, USA). The cells were incubated for 24 h and the medium was changed to remove remaining transfection reagent. Retroviral supernatant was collected at 24 and 48 h post-transfection and used to infect the target cells 293T for 48 h. Polybrene (hexadimethrine bromide) was added to the medium for improving infection efficacy. Puromycin (Sigma-Aldrich) was used to select the successfully infected cells at a final concentration of 2 µg/ml and the survived cells were collected to check the expression of human Lon by western blotting. Lon-shRNA: 5′-GAAAGUUCGUCUCGCCCAGCC-3′ (sh-1) or 5′-AGGAGCAGCUAAAGAUCAUCA-3′ (sh-2)12,17. Cultured cells were treated with hydrogen peroxide (200 µM H2O2, Sigma-Aldrich) for 4 h at 37 °C.
Antibodies
Antibodies to human Lon was produced as described previously28. Antibodies used in this study were purchased as indicated: antibody to p53 and Flag from Sigma; Myc (9E10) from Millipore; α-tubulin (ab4074) and COX4 (ab16056) from Abcam (Cambridge, MA, USA); IkB alpha and VDAC from Cell Signaling Technology (Beverly, MA, USA); Bcl-2, Bax, and cytochrome C from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA); GAPDH and beta-actin from GenTex (Hsinchu, Taiwan).
Western blot analysis
Western blot analysis was performed as described previously12,17.
Isolation of mitochondria fraction
Mitochondrial fraction was isolated by the Mitochondria Isolation Kit for Cultured Cells (Thermo Fisher Scientific, 89874) according to the manufacturer’s instructions.After collecting the mitochondria fraction from cells, the mitochondrial pellets were resuspended with Lysis buffer, NETN buffer with protease inhibitors (100 mM PMSF, 5 µg/ml aprotinin, 5 µg/ml leupeptin) on ice for 30 min and centrifuged at 12,000 × g for 15 min. Then the supernatant was collected and applied for subsequent western blotting.
Co-immunoprecipitation
Cells were lysed in NETN (150 mM NaCl, 1 mM EDTA, 20 mM Tris-Cl (pH 8.0), 0.5% NP-40) containing protease and phosphatase inhibitors (1.0 mM sodium orthovanadate, 50 μM sodium fluoride). Immunoprecipitation was performed by incubating primary antibodies with cell lysates at 4 °C for overnight, followed by the addition of secondary antibody and protein A/G-agarose (Calbiochem) for two additional hour with slow agitation and centrifugation for 15 s. The pellets were washed three times with NETN containing protease inhibitor cocktail (Roche) buffer and examined for binding partners by western blotting.
Immunofluorescence
Cells were plated on glass coverslips placed in a 12-well culture dish. When cells had attached to the surface and spread well, they were washed with cold phosphate buffered saline (abbreviated PBS) and then fixed with pre-cold methanol/ acetone (1:1, v/v) mixture for 15 minutes at room temperature. Fixed cells were washed with PBS and permeabilized with 0.5 % (v/v) Triton X-100 in PBS for 15 min at room temperature. Cells on coverslips were incubated with indicated antibodies: anti-Lon (1:400) and anti-p53 (1:200) overnight at 4 °C. The following day, fixed cells were washed three times with 0.5% Triton X-100 in PBS and incubated with Alexa 488-conjugated and Alexa-594 conjugated anti-mouse or anti-rabbit secondary antibodies. Finally, coverslips were mounted by ProLong® Gold Antifade Reagent with DAPI (Invitrogen, Carlsbad, CA) for room temperature 10 min. Fluorescent images were acquired by by an Olympus BX51 fluorescence microscopy.
Apoptosis assay
Apoptosis was analyzed by TUNEL staining or MMP measurement. Cell apoptosis was detected by TUNEL assay according to the manufacturer’s instructions (TaKaRa BIO, Shiga, Japan) and was performed as described previously12,42. Apoptotic cells also were analyzed by flow cytometry after DiOC6(3) (AnaSpec, Inc) staining. Flow cytometry and data analysis were carried out on a FACSCalibur instrument (Becton Dickinson), excitation = 488 nm; emission = 530 nm (F1), using the program Lysis.
Reverse transcription-PCR (RT-PCR)
Total RNA was extracted using Trizol reagent (Invitrogen) and reverse-transcribed using Maxima First Strand cDNA Synthesis Kit for RT-qPCR (Thermo Fisher Scientific, USA). The resulting cDNA was used as the template for PCR reactions. Real-time PCR reactions were performed using the ABI Step One Plus Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) using SYBR Green PCR Master Mix (Thermo Fisher Scientific, USA) and relative quantification was performed using the comparative 2−ΔΔCT method. The sets of forward and reverse primers, the corresponding PCR conditions, and the lengths of PCR products were described as follows: Lon (5′-GTCATGGATGTTGTGGACGA-3′, 5′-GTAGTTGCGGGTGACATTGA-3′); p53 (5′-GGCTCTGACTGTACCACCATCCA-3′, 5′-GGCACAAACACGCACCTCAAAG-3′); Fas (5′-TCTGGTTCTTACGTCTGTTGC-3′, 5′-CTGTGCAGTCCCTAGCTTTCC-3′); Bax (5′-TGCTTCAGGGTTTCATCCAG-3′, 5′-GGCGGCAATCATCCTCTG-3′); Puma (5′-GAAGAGCAAATGAGCCAAACG-3′, 5′-GGAGCAACCGGCAAACG-3′); p53R2 (5′-GAGGCTCGCTGTTTCTATGG-3′, 5′-ATCTGCTATCCATCGCAAGG-3′); Bim (5′-TGGCAAAGCAACCTTCTGATG-3′, 5′-GCAGGCTGCAATTGTCTACCT-3′). All the PCR reactions were started at 94 °C for 5 min and terminated at 72 °C for 5 min. Finally, the data were analyzed using the StepOne Software v2.3. Differential RNA expressions between various samples were calculated using β-actin as an internal control.
Protein structure and modeling
Structural modeling of human Lon was built by using I-TASSER package43 by iterative fragment assembly simulation for protein 3D structure prediction and refinement. The best predicted structure was used for further application. Docking of p53 DNA-binding domain (PDB code: 1TSR) to the structure of human Lon ATPase domain from predicted model was initially carried out using the ZDOCK server44, which employs rigid-body docking and utilizes a scoring function based on pairwise shape complementarity, desolvation, and electrostatic energies. No residue constraints were supplied as inputs for docking calculation. The structure of human Lon ATPase domain was assigned as the receptor in the docking calculation. During rigid-body energy minimization, >500 structures were calculated and the 100 best structures based on the intermolecular energy were used for semiflexible simulated annealing. Docked structures corresponding to the 100 best structures with the lowest intermolecular energies were generated. Finally, the top 1 docking models predicted by ZDOCK data was selected for our biochemical elucidation.
Statistical methods
We examined the significance of the IHC staining association between Lon and p53 protein using Fisher’s exact test and considered a statistical significance if a p value was ≤0.05. We measured the correlation in contingency table using Cramer’s V coefficient, which was calculated by CramersV function in R-based lsr package. All data were analyzed using the R statistical software (version 3.1.1). Parametric Student’s t test was used to judge the significance of difference between conditions of interest. In general, a p value of <0.05 was considered as statistically significant (*p < 0.05, **p < 0.01, and ***p < 0.001).
References
Lane, D. & Levine, A. p53 Research: the past thirty years and the next thirty years. Cold Spring Harb. Perspect. Biol. 2, a000893 (2010).
Vaseva, A. V. et al. p53 opens the mitochondrial permeability transition pore to trigger necrosis. Cell 149, 1536–1548 (2012).
Ranjan, A. & Iwakuma, T. Non-canonical cell death induced by p53. Int. J. Mol. Sci. 17, https://doi.org/10.3390/ijms17122068 (2016).
Vousden, K. H. & Prives, C. Blinded by the light: the growing complexity of p53. Cell 137, 413–431 (2009).
Mihara, M. et al. p53 has a direct apoptogenic role at the mitochondria. Mol. Cell 11, 577–590 (2003).
Galluzzi, L., Bravo-San Pedro, J. M., Kepp, O. & Kroemer, G. Regulated cell death and adaptive stress responses. Cell. Mol. Life Sci. 73, 2405–2410 (2016).
Galluzzi, L., Kepp, O., Trojel-Hansen, C. & Kroemer, G. Mitochondrial control of cellular life, stress, and death. Circ. Res. 111, 1198–1207 (2012).
Van Dyck, L. & Langer, T. ATP-dependent proteases controlling mitochondrial function in the yeast Saccharomyces cerevisiae. Cell. Mol. Life Sci. 56, 825–842 (1999).
Pinti, M. et al. Emerging role of Lon protease as a master regulator of mitochondrial functions. Biochim. Biophys. Acta 1857, 1300–1306 (2016).
Hori, O. et al. Transmission of cell stress from endoplasmic reticulum to mitochondria: enhanced expression of Lon protease. J. Cell Biol. 157, 1151–1160 (2002).
Lee, A. Y., Hsu, C. H. & Wu, S. H. Functional domains of Brevibacillus thermoruber lon protease for oligomerization and DNA binding: role of N-terminal and sensor and substrate discrimination domains. J. Biol. Chem. 279, 34903–34912 (2004).
Cheng, C. W. et al. Overexpression of Lon contributes to survival and aggressive phenotype of cancer cells through mitochondrial complex I-mediated generation of reactive oxygen species. Cell Death Dis. 4, e681 (2013).
Rep, M. et al. Promotion of mitochondrial membrane complex assembly by a proteolytically inactive yeast Lon. Science 274, 103–106 (1996).
Ngo, J. K. & Davies, K. J. Mitochondrial Lon protease is a human stress protein. Free Radic. Biol. Med. 46, 1042–1048 (2009).
Fukuda, R. et al. HIF-1 regulates cytochrome oxidase subunits to optimize efficiency of respiration in hypoxic cells. Cell 129, 111–122 (2007).
Ghosh, J. C., Dohi, T., Kang, B. H. & Altieri, D. C. Hsp60 regulation of tumor cell apoptosis. J. Biol. Chem. 283, 5188–5194 (2008).
Kao, T. Y. et al. Mitochondrial Lon regulates apoptosis through the association with Hsp60-mtHsp70 complex. Cell Death Dis. 6, e1642 (2015).
Calderwood, S. K., Khaleque, M. A., Sawyer, D. B. & Ciocca, D. R. Heat shock proteins in cancer: chaperones of tumorigenesis. Trends Biochem. Sci. 31, 164–172 (2006).
Bota, D. A., Ngo, J. K. & Davies, K. J. Downregulation of the human Lon protease impairs mitochondrial structure and function and causes cell death. Free Radic. Biol. Med. 38, 665–677 (2005).
Quiros, P. M. et al. ATP-dependent Lon protease controls tumor bioenergetics by reprogramming mitochondrial activity. Cell Rep. 8, 542–556 (2014).
Gibellini, L. et al. Silencing of mitochondrial Lon protease deeply impairs mitochondrial proteome and function in colon cancer cells. FASEB J. https://doi.org/10.1096/fj.14-255869 (2014).
Bota, D. A. & Davies, K. J. Mitochondrial Lon protease in human disease and aging: Including an etiologic classification of Lon-related diseases and disorders. Free Radic. Biol. Med. 100, 188–198 (2016).
Ambro, L. et al. Mutations to a glycine loop in the catalytic site of human Lon changes its protease, peptidase and ATPase activities. FEBS J. 281, 1784–1797 (2014).
Galluzzi, L. et al. Mitochondrial liaisons of p53. Antioxid. Redox Signal. 15, 1691–1714 (2011).
Haupt, S., Berger, M., Goldberg, Z. & Haupt, Y. Apoptosis - the p53 network. J. Cell Sci. 116, 4077–4085 (2003).
Thelander, L. Ribonucleotide reductase and mitochondrial DNA synthesis. Nat. Genet. 39, 703–704 (2007).
Kuo, M. L. et al. RRM2B suppresses activation of the oxidative stress pathway and is up-regulated by p53 during senescence. Sci. Rep. 2, 822 (2012).
Wang, H. M. et al. Obtusilactone A and (-)-sesamin induce apoptosis in human lung cancer cells by inhibiting mitochondrial Lon protease and activating DNA damage checkpoints. Cancer Sci. 101, 2612–2620 (2010).
Lilyestrom, W., Klein, M. G., Zhang, R., Joachimiak, A. & Chen, X. S. Crystal structure of SV40 large T-antigen bound top53: interplay between a viral oncoprotein and a cellular tumor suppressor. Genes Dev. 20, 2373–2382 (2006).
Fan, C. C. et al. Expression of E-cadherin, Twist, and p53 and their prognostic value in patients with oral squamous cell carcinoma. J. Cancer Res. Clin. Oncol. 139, 1735–1744 (2013).
Jego, G., Hazoume, A., Seigneuric, R. & Garrido, C. Targeting heat shock proteins in cancer. Cancer Lett. https://doi.org/10.1016/j.canlet.2010.10.014 (2010).
Ghosh, J. C., Siegelin, M. D., Dohi, T. & Altieri, D. C. Heat shock protein 60 regulation of the mitochondrial permeability transition pore in tumor cells. Cancer Res. 70, 8988–8993 (2010).
Caelles, C., Helmberg, A. & Karin, M. p53-dependent apoptosis in the absence of transcriptional activation of p53-target genes. Nature 370, 220–223 (1994).
Marchenko, N. D. & Moll, U. M. Mitochondrial death functions of p53. Mol. Cell. Oncol. 1, e955995 (2014).
Kasashima, K., Sumitani, M., Satoh, M. & Endo, H. Human prohibitin 1 maintains the organization and stability of the mitochondrial nucleoids. Exp. Cell Res. 314, 988–996 (2008).
Bayot, A. et al. Identification of novel oxidized protein substrates and physiological partners of the mitochondrial ATP-dependent Lon-like protease Pim1. J. Biol. Chem. 285, 11445–11457 (2010).
Strauss, K. A. et al. CODAS syndrome is associated with mutations of LONP1, encoding mitochondrial AAA+Lon protease. Am. J. Human. Genet. 96, 121–135 (2015).
Park, J. H., Zhuang, J., Li, J. & Hwang, P. M. p53 as guardian of the mitochondrial genome. FEBS Lett. 590, 924–934 (2016).
Zhuang, J., Ma, W., Lago, C. U. & Hwang, P. M. Metabolic regulation of oxygen and redox homeostasis byp53: lessons from evolutionary biology? Free Radic. Biol. Med. 53, 1279–1285 (2012).
Ahn, B. Y. et al. Tid1 is a new regulator of p53 mitochondrial translocation and apoptosis in cancer. Oncogene 29, 1155–1166 (2010).
Marchenko, N. D., Zaika, A. & Moll, U. M. Death signal-induced localization of p53 protein to mitochondria. A potential role in apoptotic signaling. J. Biol. Chem. 275, 16202–16212 (2000).
Cheng, A. N. et al. Increased Cdc7 expression is a marker of oral squamous cell carcinoma and overexpression of Cdc7 contributes to the resistance to DNA-damaging agents. Cancer Lett. 337, 218–225 (2013).
Yang, J. & Zhang, Y. I-TASSER server: new development for protein structure and function predictions. Nucleic Acids Res. 43, W174–W181 (2015).
Pierce, B. G. et al. ZDOCK server: interactive docking prediction of protein-protein complexes and symmetric multimers. Bioinformatics 30, 1771–1773 (2014).
Acknowledgements
This work was supported by grants from the Ministry of Science and Technology (MOST 103-2320-B-400-022-MY3, MOST105-2628-B-400-003-MY3, and MOST106-2627-M-400-002), Ministry of Health and Welfare (MOHW106-TDU-B-212-122015), and National Health Research Institutes (106A1-CA-PP-07), Taiwan to A.Y.-L.L.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Edited by I. Amelio
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sung, YJ., Kao, TY., Kuo, CL. et al. Mitochondrial Lon sequesters and stabilizes p53 in the matrix to restrain apoptosis under oxidative stress via its chaperone activity. Cell Death Dis 9, 697 (2018). https://doi.org/10.1038/s41419-018-0730-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41419-018-0730-7
This article is cited by
-
Mitochondrial Lon-induced mitophagy benefits hypoxic resistance via Ca2+-dependent FUNDC1 phosphorylation at the ER-mitochondria interface
Cell Death & Disease (2023)
-
A Fc-VEGF chimeric fusion enhances PD-L1 immunotherapy via inducing immune reprogramming and infiltration in the immunosuppressive tumor microenvironment
Cancer Immunology, Immunotherapy (2023)
-
MiR-601-induced BMSCs senescence accelerates steroid-induced osteonecrosis of the femoral head progression by targeting SIRT1
Cellular and Molecular Life Sciences (2023)
-
Glucocorticoid-induced activation of NOX/ROS/NF-κB signaling in MSCs contributes to the development of GONFH
Apoptosis (2023)
-
Mitochondrial oxidative stress in the tumor microenvironment and cancer immunoescape: foe or friend?
Journal of Biomedical Science (2022)
