
Abstract
In the past two decades, purinergic signaling has emerged as a key regulator of hematopoiesis in physiological and pathological conditions. ADP receptor P2y12 is a crucial component of this signaling, but whether it is involved in primitive hematopoiesis remains unknown. To elucidate the function of P2y12 and provide new insights for drug development, we established a zebrafish P2y12 mutant by CRISPR/Cas 9-based genetic modification system, and investigated whether P2y12 acted as an important regulator for primitive hematopoiesis. By using mass spectrometry (MS) combined with RNA sequencing, we showed that absence of P2y12 induced excessive erythropoiesis, evidenced by significantly increased expression of mature erythrocytes marker α-globin (Hbae1 and Hbae3), β-globin (Hbbe1 and Hbbe3). Expression pattern analysis showed that P2y12 was mainly expressed in red blood cells and endothelial cells of early zebrafish embryos. Further studies revealed that primitive erythroid progenitor marker Gata1 was markedly up-regulated. Remarkably, inhibition of Gata1 by injection of Gata1 morpholino could rescue the erythroid abnormality in P2y12 mutants. The present study demonstrates the essential role of purinergic signaling in differentiation of proerythrocytes during primitive hematopoiesis, and provides potential targets for treatment of blood-related disease and drug development.
Similar content being viewed by others
Introduction
All vertebrate organisms experience waves of hematopoiesis during their lifetime [1]. Hematopoiesis occurs in two successive waves during vertebrate development: the primitive wave generates erythrocytes expressing genes encoding embryonic hemoglobin and macrophages, and the definitive wave produces hematopoietic stem cells (HSCs), which can give rise to all blood lineages [1, 2]. Although the occurrence sites vary from those of mammals, zebrafish are an ideal model to study vertebrate hematopoiesis due to the high conservation in the developmental processes and genetic programs of hematopoiesis [1, 3, 4]. Hematopoiesis is a dynamic process that requires the regulation of a sophisticated network. Previous studies have addressed the core role of Gata1 in erythropoiesis. Mice with Gata1 knockout died between E10.5 and E11.5, since progenitor erythrocytes failed to differentiate into mature erythrocytes [5]. Consistent with previous studies in mice, the zebrafish Gata1 mutant also had a complete block in primitive hematopoiesis [6]. However, the upstream regulatory mechanisms of Gata1 are still unclear.
The purine signaling pathway serves as an unconventional modulator of hematopoiesis [7] and includes ATP and its products ADP, AMP, and adenosine [7]. In the receptor family, the ADP receptor P2y12, which participates in platelet aggregation and thrombus formation [8, 9], is a well-known therapeutic target for antithrombotic and antiplatelet treatments [10, 11] and cardiovascular disease [12]. Patients with congenital P2y12 deficiency experience mild to severe bleeding, and whether these patients have other disorders is not known [13,14,15,16,17]. Intriguingly, P2y12 could be detected on the surface of human hematopoietic cells in addition to platelets [18, 19]. Increasing evidence has demonstrated the comprehensive function of P2y12 in macrophages [20], microglia [21, 22], leukocytes [23] and nonhematopoietic cells [24,25,26]. However, whether P2y12 is involved in primitive hematopoiesis remains unknown.
In the present study, we established a zebrafish model with a P2y12 mutant phenotype and confirmed the essential role of P2y12 in the differentiation of progenitor erythrocytes during primitive hematopoiesis. Further, we found that the hematopoietic transcription factor Gata1 is required for the P2y12 mutant phenotype. These results suggest that P2y12 controls erythrocyte development by regulating Gata1 and provides a novel perspective for the application of the therapeutic target P2y12.
Materials and methods
Zebrafish care and lines
The zebrafish lines Tubingen, Tg(Gata1:egfp), and Tg(Fli1:egfp)y1were raised, mated and staged as described previously [27,28,29]. The fish maintenance was in accordance with the guidelines of the Institutional Review Board of the Institute of Health Sciences, Shanghai Institutes of Biological Sciences, Chinese Academy of Sciences (Shanghai, China).
Morpholino (MO) injections
All morpholinos were purchased from GeneTools (USA) and dissolved in RNase-free water (without treatment with DEPC). A scramble MO was used as a control. Then, 1 nL of MO was injected into one-cell stage embryos, and the concentrations were as follows unless specified otherwise: Gata1 MO [30], 0.25 mmol/L; control MO, 0.25 mmol/L.
Self-rescue experiment
The coding sequence of P2y12 was cloned into the vector PCS2+. Capped P2y12 mRNA was synthesized with a Sp6 kit (Promega, USA), and ~200 pg of P2y12 mRNA was injected into each embryo (sibling or P2y12 mutant) at the one-cell stage. Then, we extracted total RNA from embryos at 20 h postfertilization (hpf) and quantified the RNA level of Hbae1 by RT-PCR (see below for details).
Genome editing and genotyping
Cas9 mRNA was synthesized by in vitro transcription using T7 RNA polymerase (Roche, Germany). The target site of gRNA was designed manually. Cas9 mRNA and gRNA were coinjected into embryos at the one-cell stage. Each embryo was injected with 1 nL of solution containing Cas9 mRNA (200 ng/μL) and gRNA (100 ng/μL). We designed primers to amplify the gRNA target region and screened the founders by sequencing. The primers for genotype identification are shown in the Supplementary Table.
Mass spectrometry (MS)
The quantitative mass spectrometry (MS) analysis was performed by the National Center for Protein Science Shanghai. Whole embryos (siblings and P2y12 mutants) were deyolked at 28 hpf and then lysed in 8 M urea. Each group contained 80 embryos. TCEP (final concentration is 5 mM) (Thermo Scientific, USA) and iodoacetamide (final concentration is 10 mM) (Sigma, USA) for reduction and alkylation were added to the solution and incubated at room temperature for 20 min and 15 min, respectively, and then digested with trypsin overnight. The peptide mixture was analyzed by a homemade 15 cm-long pulled-tip analytical column (75 µm i.d.) packed with 3 µm reversed-phase beads. The column was then placed in-line with an Easy-nLC 1000 nano HPLC (Thermo Scientific, USA) for MS analysis with a flow rate of 300 nL/min. Data-dependent tandem mass spectrometry (MS/MS) analysis was performed with a Q Exactive Orbitrap mass spectrometer (Thermo Scientific, USA). Peptides eluted from the LC column were directly electrosprayed into the mass spectrometer with the application of a distal 1.8-kV spray voltage. A cycle of one full-scan MS spectrometer (m/z 300–1800) was acquired, followed by the top 20 MS/MS events. Acquired MS/MS data were analyzed against aUniProtKB zebrafish database (database released on Nov. 15, 2016) by the Andromeda search engine integrated in MaxQuant software. Mass tolerances for precursor ions were set at 20 ppm, and those for MS/MS were set at 0.02 Da. Trypsin was defined as the cleavage enzyme; cysteine alkylation by iodoacetamide was specified as a fixed modification with a mass shift of 57.02146. To accurately estimate peptide probabilities and false discovery rates, we used a decoy database containing the reverse sequences of all the proteins appended to the target database, and the FDR was set as 0.01. Label-free quantification was conducted by the LFQ algorithm by MaxQuant.
Volcano map and GO enrichment analysis were performed with online tools (https://www.omicsolution.org/wkomics/Volcano/) and DAVID Functional Annotation Tools, respectively.
RNA sequencing
Whole embryos (siblings and P2y12 mutants) were collected at 24 hpf, and each group had at least 40 embryos. Total RNA was extracted from the embryos using TRIzol reagent (Invitrogen) according to the protocol. RNA sequencing was performed by WuXiNextCODE at Shanghai.
Quantitative reverse transcriptase-polymerase chain reaction (RT-PCR)
Total RNA was extracted from whole embryos as described for RNA sequencing. The total RNA extracted was used to generate cDNA by using Super Script II reverse transcriptase with random primers for mRNA. RT-PCR was performed using SYBR Green (Toyobo, Japan). The relative RNA amount was calculated with the ΔΔCt method and normalized to the internal control β-actin.
Western blotting
Whole embryos (siblings and P2y12 mutants) were deyolked at the indicated time points, lysed in RIPA buffer containing proteinase inhibitor (Roche, Germany), and denatured in SDS loading buffer. The antibodies used in this study were as follows: tubulin antibody (Yeasen, China, 1:1000); HBB antibody (Abclonal, China, 1:1000); and Gata1 antibody (Abcam,UK, 1:1000).
Whole-mount in situ hybridization (WISH)
T7 polymerase (Roche, Germany) and digoxigenin (Roche, Germany) were used for probe synthesis. Then, whole-mount in situ hybridization was conducted in wild-type embryos as previously described [31]. The sequences of the probes for Lmo2 [32], Runx1, C-myb [33], Gata1, and Kdrl [34] were described previously.
Double fluorescence in situ hybridization
Double fluorescence in situ hybridization with a digoxigenin (Roche, Germany)-labeled probe (P2y12) and fluorescein (Roche, Germany)-labeled probe (Kdrl and Hbae1) was performed as previously described [31]. In contrast to WISH, the embryos were blocked with 2% Blocking Reagent (Roche, Germany) for 1 h at room temperature and incubated in a solution of anti-digoxigenin-POD Fab Fragments (Roche, Germany, 1:1000) on the second day. The samples were stained with a buffer containing Cy3 (PerkinElmer, UK, 1:50) for 40 min, and then, immunofluorescence was performed. Then, the staining reaction was stopped with 4% PFA, and the embryos were washed with PBST. The embryos were reblocked with 2% Blocking Reagent (Roche, Germany) and incubated in a solution of anti-fluorescein-POD Fab Fragments (Roche, Germany, 1:1000). The embryos were stained with a buffer containing FITC (PerkinElmer, UK, 1:50) for 20 min and then washed with PBST.
Confocal imaging and data analysis
Confocal images were obtained as previously described [35, 36]. The destination regions were scanned with a 1 μm step size, and the format was 1024 × 1024 pixels at 400 Hz. All the confocal images are lateral views, dorsal is up, and anterior is to the left unless specifically noted.
Dual-luciferase reporter assay
The Gata1 promoter sequences (−2000 bp to transcription start site) were inserted into the pGL3-Basic vector (Promega, USA). In brief, 293T cells were seeded in 12-well plates. Then, 50 ng Renilla with (500 ng PCS2+ and 500 ng PGL3-Basic), (500 ng PCS2+ and 500 ng PGL3-Basic-Gata1 promoter), (500 ng PCS2+-P2y12 and 500 ng PGL3-Basic) or (500 ng PCS2+-P2y12 and 500 ng PGL3-Basic-Gata1 promoter) was transfected into 293T cells. After 48 h, cells were collected, and the firefly and Renilla luciferase activities were measured by the Dual-Luciferase Reporter Assay System (DL101, Vazyme, China) according to the manufacturer’s instructions.
Statistical analysis
At least three independent experiments were performed. All the data are presented as the mean ± SD. Statistical analyses were performed using GraphPad. Analysis of differences between two groups was conducted with unpaired Student’s two-tailed t test. When more than two groups were compared, significant differences were determined with one-way ANOVA with Tukey’s post hoc test. Differences were considered significant when P < 0.05. Probability values are indicated by *(P < 0.05), **(P < 0.01), or ***(P < 0.001).
Results
P2y12 is involved in early embryonic development, especially in hematopoiesis
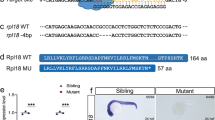
To explore the in vivo function of P2y12, we constructed P2y12 knockout lines using the CRISPR/Cas9 system (Fig. 1a–c). This mutation results in a frame shift and a premature stop codon. The altered transcript is predicted to encode a truncated P2y12 peptide with 117 amino acids (Fig. 1c). The survival rate of the P2y12 mutants was significantly lower than that of the siblings (Fig. S1a). The embryos that survived developed normally to adult hood (Fig. S1b). However, the mutants were prone to lepidorthosis and lived for ~1 year (data not shown). To further assess the ADP receptor P2y12, we conducted MS and obtained 31 genes affected by P2y12 deficiency. The result showed that there were 8 upregulated genes and 23 downregulated genes using a fold change of two as a cutoff in MS (Fig. 1d). Gene ontology (GO) analysis revealed that the upregulated genes of the P2y12 mutant were highly enriched in hemoglobin structure and function (Fig. 1e), which suggested that the globin structure and function were affected in the P2y12 mutants. For the downregulated genes, the categories included protease, hydrolase and hydrolase activity (Fig. 1e). Integration of the altered genes in MS and RNA-seq data showed that Alox5b, Vtg1, and Acad9 had changes in the opposite directions. Notably, hemoglobin-related proteins (Hbae1, Hbae3, Hbbe1 and Hbbe3) were upregulated at both the protein and mRNA levels (Fig. 1f, g). All these results demonstrated that P2y12 is involved in early embryonic development, especially in hematopoiesis.

a Partial structure and sequence of the P2y12 gene showing the target site of gRNA. PAM, protospacer adjacent motifs; S and AS, a pair of primers used for amplifying the mutation site +1, the translation initiation site; +1038, the stop codon site of P2y12; +308, the site of the gRNA. b Detection of mutations in the F0 embryos by Alw21I digestion. Fragments with potential indels at the Alw21I site are indicated by red arrow (the uncut band), and fragments from the wild type are indicated by black arrows (cut bands). c The sequence of the P2y12 cDNA from mutant embryos shows a 7 bp deletion (MU) compared with that of the WT embryos. d Mass spectrometry (MS) volcano map of the siblings and the P2y12 mutants at 28 hpf. Fold change > 2 and <0.5, P value < 0.05. Downregulated genes are shown in blue dots, and upregulated genes are shown in red dots. e Bubble chart of the differentially expressed genes (DEGs) in MS. f Heat map of the upregulated genes in MS and the corresponding changes in RNA sequencing. Sc8*:si:ch211–154a22.8; Sd9*:si:dkey-90l23.1. g Heat map of the downregulated genes in MS and the corresponding changes in RNA sequencing.
P2y12 is specifically enriched in the red blood cells and the vascular endothelial cells of zebrafish embryos
We analyzed the P2y12 protein sequence, which contains 345 amino acids and displays 53.6% identity (81.7% similarity) to the human P2Y12 protein and 51.9% identity (81.4% similarity) to the mouse P2y12 protein (Fig. 2a). The results indicated that P2y12 is highly evolutionarily conserved in vertebrates. Then, we analyzed P2y12 expression by WISH. The results showed that P2y12 was expressed at the one-cell stage, which indicated that it is a maternal gene in zebrafish (Fig. 2b). Furthermore, we observed ubiquitous expression of P2y12 at the four-cell stage, 6 hpf and 12 hpf (Fig. 2b), along with a spotty distribution in the head at 72 hpf (Fig. 2b). Intriguingly, P2y12 was enriched in the intermediate cell mass (ICM) and the posterior blood island at 24 hpf (Fig. 2b). To further examine the expression pattern, we performed double fluorescence in situ hybridization (DFISH) for P2y12 and the endothelial cell marker Kdrl. The results showed that P2y12 colocalized with Kdrl in the dorsal aorta and the posterior cardinal vein at 24 hpf (Fig. 2c). In particular, we found that P2y12 was expressed in blood cells (Fig. 2c). To investigate the specific type of blood cells expressing P2y12, we performed DFISH for P2y12 and the red blood cell marker Hbae1, which indicated that P2y12 is mainly expressed in red blood cells (Fig. 2d). Together, these observations demonstrate that P2y12 is expressed specifically in red blood cells and vascular endothelial cells in early embryonic development.

a Alignment of the zebrafish P2y12 protein and its closest human and mouse homologs. Identical and similar amino acids are shaded in gray. b Expression pattern of P2y12 analyzed by WISH from the one-cell stage to 72 hpf, and the developmental time points are indicated in the panels. The intermediate cell mass (ICM) and the posterior blood island (PBI) are indicated by black brackets. The dorsal aorta (DA) and the posterior cardinal vein (PCV) are indicated by red brackets and blue brackets, respectively. c Double fluorescence in situ hybridization (DFISH) for P2y12 and the endothelial cell marker Kdrl in 24 hpf embryos. The dorsal aorta (DA) and the posterior cardinal vein (PCV) are indicated by red brackets and blue brackets, respectively. d DFISH for P2y12 and the red blood cell marker Hbae1 in 24 hpf embryos. Scale bar = 100 μm (b); 25 μm (c, d).
Primitive hematopoiesis was disturbed in zebrafish with the P2y12 mutation
Similar to mammalian hematopoiesis, zebrafish hematopoiesis proceeds through two successive stages: primitive hematopoiesis and definitive hematopoiesis [1, 2]. To assess the function of P2y12 in hematopoiesis, we detected primitive hematopoiesis markers and found that the expression of the mature erythrocyte markers α-globin (Hbae1 and Hbae3) and β-globin (Hbbe1 and Hbbe3) was dramatically increased in the P2y12 mutants, as shown by whole-mount in situ hybridization (WISH) at 22 hpf (Fig. 3a). Further RT-PCR analysis also showed that the mRNA levels of α-globin and β-globinin in the P2y12 mutants were dramatically higher than those of the sibling embryos (Fig. 3b, c). As predicted, the trend of the β-globin protein level in the P2y12 mutants was consistent with that of the mRNA level (Fig. 3d–g). However, the expression of globin was restored to normal levels with further embryonic development (Fig. S2). To assess the aberrant primitive hematopoiesis resulting from the absence of P2y12, we injected P2y12 mRNA into one-cell stage siblings or P2y12 mutants to observe the rescue effects. The results showed that Hbae1 could be partially rescued by P2y12 mRNA in the P2y12 mutants (Fig. 3h). We further examined the function of P2y12 in definitive hematopoiesis, and the WISH results demonstrated that the HSC marker C-myb was not affected in embryos with P2y12 mutations (Fig. S3). Thus, P2y12 is responsible for primitive hematopoiesis in zebrafish.

a Detection of theHbae1, Hbae3, Hbbe1, and Hbbe3 mRNA levels by whole-mount in situ hybridization (WISH) in the siblings and the P2y12 mutants at 22 hpf. The number in the bottom left-hand corner indicates the representative embryos/total embryos. b–e Quantification of Hbae1, Hbae3, Hbbe1, and Hbbe3 expression at 22 hpf by real-time PCR. Statistical analysis was conducted using unpaired Student’s two-tailed t test. f Detection of the HBB protein levels by western blotting in the siblings and the P2y12 mutants at 28 hpf. Each group had more than 30 embryos, and this assay was performed three times. g Quantitative analysis of three western blotting results. Statistical analysis was conducted using unpaired Student’s two-tailed t test. h Quantitative analysis of Hbae1 in the siblings, the siblings injected with P2y12 mRNA, the P2y12 mutants, and the P2y12 mutants injected with P2y12 mRNA. Statistical differences were analyzed by one-way ANOVA with Tukey’s post-hoc test. Scale bar = 100 μm (a). *P < 0.05, **P < 0.01 and ***P < 0.001 compared with the siblings; ##P < 0.05 compared with the P2y12 mutants. All results are presented as the mean ± SD and are representative of three independent experiments.
P2y12 does not participate in vascular and lymphatic development
Based on the expression of P2y12 in endothelial cells, we traced vascular development using the endothelial cell-labeled zebrafish line Tg(Fli1:egfp)y1. In angiogenesis, endothelial tip cells sense the extracellular guidance cues and lead the way for other endothelial cells [37]. To test whether P2y12 was involved in vascular development, we selected key events during this process, and we found no obvious difference. In both siblings and mutants, endothelial tip cells migrated to the horizontal myoseptum at 24 hpf (Fig. 4a, b), and intersegmental vessels reached the dorsal roof and formed dorsal longitudinal anastomotic vessels (Fig. 4a, c). The venous endothelium is one of the origins of lymphatic vessels [38, 39]. In the P2y12 mutants, the formation of parachordal lymphangioblasts and thoracic ducts was normal compared with that in the siblings (Fig. 4d–f). Thus, P2y12 is dispensable for vascular and lymphatic development.

a Confocal stack micrographs of trunk vessels in a lateral view of the siblings and the P2y12 mutants with the Tg(Fli1:egfp)y1 background at 24 hpf (upper panels) and 30 hpf (bottom panels). Arrows and arrowheads indicate ISV and DLAV, respectively. b Percentage of embryos with normal ISVs at 24 hpf. n = 450 (siblings) and 154 (P2y12 mutants). c Percentage of embryos with normal DLAVs at 30 hpf. n = 449 (siblings) and 140 (P2y12 mutants). d Confocal stack micrographs of the parachordal lymphangioblast (PL) (upper panels, at 72 hpf) and the thoracic duct (TD) (bottom panels, at 120 hpf) in a lateral view of the siblings and the P2y12 mutants with the Tg(Fli1:egfp)y1 background. Arrowheads indicate PLs, and white dotted lines indicate TDs. e Percentage of embryos with normal PLsat 72 hpf. n = 448 (siblings) and 139 (P2y12 mutants). f Percentage of embryos with normal TDs at 120 hpf. n = 139 (siblings) and 130 (P2y12 mutants). Statistical analysis was conducted using unpaired Student’s two-tailed t test. Scale bar = 100 μm (a and upper panels in d) and 50 μm (bottom panels in d). All results are presented as the mean ± SD and are representatives of three independent experiments.
P2y12 participates in the differentiation of proerythrocytes by upregulation of Gata1 at the transcriptional level
To investigate the molecular mechanism underlying P2y12-mediated primitive hematopoiesis, we examined potential genes by real-time quantitative PCR of the siblings and the P2y12 mutants. The results demonstrated that the hemangioblast marker Lmo2 was slightly increased and the primitive erythroid progenitor marker Gata1 was apparently upregulated inthe P2y12 mutants. The hemangioblast marker Scl, primitive myeloid progenitor marker Pu.1, primitive macrophage marker l-plastin, and neutrophil marker Mpo were comparable to those in the siblings at 18 hpf (Fig. 5a). In addition, the RNA sequencing results showed that Gata1 was still upregulated in the P2y12 mutants at 24 hpf, and Lmo2 was unchanged (Fig. 5b). Consistent with the RT-PCR results, we confirmed the upregulation of Gata1 in the P2y12 mutants at 18 hpf (Fig. 5c), while Lmo2 expression remained unchanged, as shown by WISH (Fig. S4). Importantly, the protein level of Gata1 was also increased in the P2y12 mutants (Fig. 5d, e).

a Quantification of Lmo2, Scl, Gata1, Pu.1, l-plastin, and Mpo expression at 18 hpf by real-time PCR. b FPKM value of Lmo2 and Gata1 in RNA sequencing (siblings and P2y12 mutants at 24 hpf). c Detection of Gata1 by WISH in the 18 hpf siblings and P2y12 mutants. The number in the bottom left-hand corner indicates the representative embryos/total embryos. d Detection of Gata1 in the siblings and the P2y12 mutants by Western blotting at 24 hpf. e Quantitative analysis of three Western blotting results. Statistical analysis was conducted using unpaired Student’s two-tailed t test. f Fluorescence images of the siblings and the P2y12 mutants with theTg(Gata1:egfp) background. g Detection of Hbae1 by WISH in the siblings and the P2y12 mutants injected with control MO or Gata1 MO. The number in the bottom left-hand corner indicates the representative embryos/total embryos. h Quantification of Hbae1 in the 22 hpf siblings and P2y12 mutants injected with the control MO or the Gata1 MO by real-time PCR. Statistical analysis was conducted using unpaired Student’s two-tailed t test. Scale bar = 100 μm (c); 200 μm (g). *P < 0.05, **P < 0.01, and ***P < 0.001 compared with the siblings; ###P < 0.01 compared with the P2y12 mutants. All results are presented as the mean ± SD and are representative of three independent experiments.
To investigate whether P2y12 directly regulates Gata1, we crossed the P2y12 mutant line with the Tg(Gata1:egfp) transgenic line, and further observation showed that the fluorescence intensity of the P2y12 mutants was stronger than that of their siblings with the Tg(Gata1:egfp) background (Fig. 5f). The upregulation of fluorescence intensity in the P2y12 mutants indicated that P2y12 may regulate Gata1 via its promoter. In addition, we performed an in vitro luciferase assay, and the results showed that P2y12 inhibited the transcriptional activity of the Gata1 promoter, which indicated that P2y12 directly regulated the promoter of Gata1 (Fig. S5).
Combined with the above results, we presumed that Gata1 was a functional downstream effector of P2y12. Then, we blocked the translation of Gata1 by a Gata1 morpholino, and the results showed that the expression of Hbae1 was dramatically inhibited in the siblings injected with the Gata1 MO (Fig. 5g-a′, c′ and h), which is consistent with previous reports [30]. In the P2y12 mutants injected with the Gata1 MO, the expression level of Hbae1 was significantly downregulated (Fig. 5g-b′, d′, h) and was comparable to that in the siblings (Fig. 5g-a′, d′, h). All of the above results revealed that the upregulation of Hbae1 in the P2y12 mutants could be rescued by inhibiting Gata1. All these results demonstrated that the upregulation of Hbae1 in the P2y12 mutants could be counteracted by Gata1 inhibition, which indicated that P2y12 controlled proerythrocyte differentiation by altering the expression of Gata1.
Discussion
In this study, we demonstrated that the ADP receptor P2y12 is an essential component for primitive hematopoiesis in zebrafish and controls the differentiation of progenitor erythrocytes by regulating Gata1 at the transcriptional level.
P2y12 is expressed and functions in various hematopoietic cells [18,19,20, 22, 23]. We detected P2y12 expression in the red blood cells of zebrafish, and the localization of P2y12 in the microglia at 72 hpf is consistent with a previous study [21, 22]. The roles of P2y12 in platelet activation/aggregation [8], thrombus formation [9], and inflammation [9, 21, 22] have been verified, but it is unclear whether P2y12 is involved in primitive hematopoiesis. For the first time, we constructed Cas9 mutants of P2y12 and identified the relationship between purinergic signaling and primitive hematopoiesis. Previous studies of zebrafish utilized MO or a selective inhibitor (MRS2395) of P2y12 [21, 22]. Our work provides a better model to elucidate the function of P2y12 in physiological and pathological conditions.
It has already been confirmed that the ADP receptor P2y12 participates in platelet aggregation and thrombus formation [8, 9]. Our work showed that P2y12 is involved in embryonic early development. Of note, although vasculature development was normal in the P2y12 mutants, we cannot exclude the function of P2y12 in endothelial cells. The first potential possibility is genetic compensation, and another possibility is that this function may be present under specific stress conditions. With regard to the excessive mature erythrocytes in the P2y12 mutants, P2y12 mRNA can rescue the phenotype of the P2y12 mutants, and the results suggested that aberrant primitive hematopoiesis results from P2y12 deficiency. At present, we have not found obvious developmental abnormalities caused by a deficiency in primitive hematopoiesis in the P2y12 mutant embryos under normal conditions. However, due to the expression of P2y12 in the microglia, which is sensitive to neuroinflammatory diseases [40], it remains to be determined whether there are abnormalities in the P2y12 mutants under stress conditions. Globin could return to normal levels in subsequent embryonic development. This phenomenon may be due to definitive hematopoiesis or compensation of other signaling pathways.
As described previously, the central role of Gata1 in primitive erythroidlineage development has been identified [5, 6, 30]. The upstream regulatory mechanism of Gata1 has not been systematically studied. In mice, the enhancers mHS-3.5 and mHS-25/6 collectively regulate erythroid Gata1 expression in cis [41]. In zebrafish, regulators modulate Gata1 by an epigenetic mechanism or posttranscriptional control [42, 43]. Our work presented herein demonstrates that the ADP receptor P2y12 controls the expression of Gata1 at the transcriptional level during primitive hematopoiesis and functions upstream of Gata1, as inhibition of Gata1 can rescue erythroid differentiation in the P2y12 mutants.
Several potential mechanisms may underlie the regulation of P2y12 towards Gata1. One possibility is that P2y12 may directly regulate the transcription of Gata1. Our results both in vitro (Fig. S5) and in vivo (Fig. 5f) indicated that P2y12 directly regulated the transcriptional activity of the Gata1 promoter. In addition, the diverse distribution of P2y12 within cells, i.e., in the mitochondria and cell membrane [44, 45], indicated that P2y12 may localize in the nucleus to directly regulate the expression of Gata1. Alternatively, membrane P2y12 signaling-mediated posttranslational modification of an unknown factor may confer modulatory effects on Gata1. Furthermore, our RNA-seq data suggested that the G-protein coupled receptor signaling pathway was downregulated in the P2y12 mutants (Fig. S6), which may contribute to the modulation of Gata1 or be involved in regulating primitive hematopoiesis independently of Gata1.
P2y12 is a well-known drug target [10, 11], but current drugs targeting the receptor are still insufficient. For example, clopidogrel has a very long half-life and ticagrelor has characteristic adverse effects [46, 47]. Our work presented here provides a novel evaluation standard for drug safety, and P2y12 may be a potential target for the treatment of blood-related diseases.
References
Paik EJ, Zon LI. Hematopoietic development in the zebrafish. Int J Dev Biol. 2010;54:1127–37.
Jing L, Zon LI. Zebrafish as a model for normal and malignant hematopoiesis. Dis Model Mech. 2011;4:433–8.
Orkin SH, Zon LI. Hematopoiesis: an evolving paradigm for stem cell biology. Cell. 2008;132:631–44.
Galloway JL, Zon LI. Ontogeny of hematopoiesis: examining the emergence of hematopoietic cells in the vertebrate embryo. Curr Top Dev Biol. 2003;53:139–58.
Fujiwara Y, Browne CP, Cunniff K, Goff SC, Orkin SH. Arrested development of embryonic red cell precursors in mouse embryos lacking transcription factor GATA-1. Proc Natl Acad Sci USA. 1996;93:12355–8.
Belele CL, English MA, Chahal J, Burnetti A, Finckbeiner SM, Gibney G, et al. Differential requirement for Gata1 DNA binding and transactivation between primitive and definitive stages of hematopoiesis in zebrafish. Blood. 2009;114:5162–72.
Burnstock G. Blood cells: an historical account of the roles of purinergic signalling. Purinergic Signal. 2015;11:411–34.
Born GV. Adenosine diphosphate as a mediator of platelet aggregation in vivo: an editorial view. Circulation. 1985;72:741–6.
Li D, Wang Y, Zhang L, Luo X, Li J, Chen X, et al. Roles of purinergic receptor P2Y, G protein-coupled 12 in the development of atherosclerosis in apolipoprotein E-deficient mice. Arterioscler Thromb Vasc Biol. 2012;32:e81–9.
Nylander S, Mattsson C, Ramstrom S, Lindahl TL. The relative importance of the ADP receptors, P2Y12 and P2Y1, in thrombin-induced platelet activation. Thromb Res. 2003;111:65–73.
Andre P, Delaney SM, LaRocca T, Vincent D, DeGuzman F, Jurek M, et al. P2Y12 regulates platelet adhesion/activation, thrombus growth, and thrombus stability in injured arteries. J Clin Investig. 2003;112:398–406.
Storey RF. The P2Y12 receptor as a therapeutic target in cardiovascular disease. Platelets. 2001;12:197–209.
Cattaneo M, Lecchi A, Randi AM, McGregor JL, Mannucci PM. Identification of a new congenital defect of platelet function characterized by severe impairment of platelet responses to adenosine diphosphate. Blood. 1992;80:2787–96.
Nurden P, Savi P, Heilmann E, Bihour C, Herbert JM, Maffrand JP, et al. An inherited bleeding disorder linked to a defective interaction between ADP and its receptor on platelets. Its influence on glycoprotein IIb-IIIa complex function. J Clin Investig. 1995;95:1612–22.
Shiraga M, Miyata S, Kato H, Kashiwagi H, Honda S, Kurata Y, et al. Impaired platelet function in a patient with P2Y12 deficiency caused by a mutation in the translation initiation codon. J Thromb Haemost. 2005;3:2315–23.
Cunningham MR, Nisar SP, Cooke AE, Emery ED, Mundell SJ. Differential endosomal sorting of a novel P2Y12 purinoreceptor mutant. Traffic. 2013;14:585–98.
Daly ME, Dawood BB, Lester WA, Peake IR, Rodeghiero F, Goodeve AC, et al. Identification and characterization of a novel P2Y 12 variant in a patient diagnosed with type 1 von Willebrand disease in the European MCMDM-1VWD study. Blood. 2009;113:4110–3.
Rossi L, Salvestrini V, Ferrari D, Di Virgilio F, Lemoli RM. The sixth sense: hematopoietic stem cells detect danger through purinergic signaling. Blood. 2012;120:2365–75.
Bernhard MK, Ulrich K. Rt-PCR study of purinergic P2 receptors in hematopoietic cell lines. Biochemistry. 2006;71:607–11.
Kronlage M, Song J, Sorokin L, Isfort K, Schwerdtle T, Leipziger J, et al. Autocrine purinergic receptor signaling is essential for macrophage chemotaxis. Sci Signal. 2010;3:ra55.
Sieger D, Moritz C, Ziegenhals T, Prykhozhij S, Peri F. Long-range Ca2+ waves transmit brain-damage signals to microglia. Dev Cell. 2012;22:1138–48.
Liu C, Wu C, Yang Q, Gao J, Li L, Yang D, et al. Macrophages mediate the repair of brain vascular rupture through direct physical adhesion and mechanical traction. Immunity. 2016;44:1162–76.
Diehl P, Olivier C, Halscheid C, Helbing T, Bode C, Moser M. Clopidogrel affects leukocyte dependent platelet aggregation by P2Y12 expressing leukocytes. Basic Res Cardiol. 2010;105:379–87.
Ben Addi A, Cammarata D, Conley PB, Boeynaems JM, Robaye B. Role of the P2Y12 receptor in the modulation of murine dendritic cell function by ADP. J Immunol. 2010;185:5900–6.
Hogberg C, Svensson H, Gustafsson R, Eyjolfsson A, Erlinge D. The reversible oral P2Y12 antagonist AZD6140 inhibits ADP-induced contractions in murine and human vasculature. Int J Cardiol. 2010;142:187–92.
Wihlborg AK, Wang L, Braun OO, Eyjolfsson A, Gustafsson R, Gudbjartsson T, et al. ADP receptor P2Y12 is expressed in vascular smooth muscle cells and stimulates contraction in human blood vessels. Arterioscler Thromb Vasc Biol. 2004;24:1810–5.
Siekmann AF, Lawson ND. Notch signalling limits angiogenic cell behaviour in developing zebrafish arteries. Nature. 2007;445:781–4.
Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF. Stages of embryonic development of the zebrafish. Dev Dyn. 1995;203:253–310.
Chi NC, Shaw RM, De Val S, Kang G, Jan LY, Black BL, et al. Foxn4 directly regulates tbx2b expression and atrioventricular canal formation. Genes Dev. 2008;22:734–9.
Galloway JL, Wingert RA, Thisse C, Thisse B, Zon LI. Loss of gata1 but not gata2 converts erythropoiesis to myelopoiesis in zebrafish embryos. Dev Cell. 2005;8:109–16.
Thisse C, Thisse B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat Protoc. 2008;3:59–69.
Wang L, Zhang Y, Zhou T, Fu YF, Du TT, Jin Y, et al. Functional characterization of Lmo2-Cre transgenic zebrafish. Dev Dyn. 2008;237:2139–46.
North TE, Goessling W, Walkley CR, Lengerke C, Kopani KR, Lord AM, et al. Prostaglandin E2 regulates vertebrate haematopoietic stem cell homeostasis. Nature. 2007;447:1007–11.
Jing CB, Chen Y, Dong M, Peng XL, Jia XE, Gao L, et al. Phospholipase C gamma-1 is required for granulocyte maturation in zebrafish. Dev Biol. 2013;374:24–31.
Chen J, Zhu RF, Li FF, Liang YL, Wang C, Qin YW, et al. MicroRNA-126a directs lymphangiogenesis through interacting with chemokine and Flt4 signaling in zebrafish. Arterioscler Thromb Vasc Biol. 2016;36:2381–93.
Zou J, Li WQ, Li Q, Li XQ, Zhang JT, Liu GQ, et al. Two functional microRNA-126s repress a novel target gene p21-activated kinase 1 to regulate vascular integrity in zebrafish. Circ Res. 2011;108:201–9.
Gerhardt H, Golding M, Fruttiger M, Ruhrberg C, Lundkvist A, Abramsson A, et al. VEGF guides angiogenic sprouting utilizing endothelial tip cell filopodia. J Cell Biol. 2003;161:1163–77.
Nicenboim J, Malkinson G, Lupo T, Asaf L, Sela Y, Mayseless O, et al. Lymphatic vessels arise from specialized angioblasts within a venous niche. Nature. 2015;522:56–61.
Martinez-Corral I, Ulvmar MH, Stanczuk L, Tatin F, Kizhatil K, John SW, et al. Nonvenous origin of dermal lymphatic vasculature. Circ Res. 2015;116:1649–54.
Mildner A, Huang H, Radke J, Stenzel W, Priller J. P2Y12 receptor is expressed on human microglia under physiological conditions throughout development and is sensitive to neuroinflammatory diseases. Glia. 2017;65:375–87.
Drissen R, Guyot B, Zhang L, Atzberger A, Sloane-Stanley J, Wood B, et al. Lineage-specific combinatorial action of enhancers regulates mouse erythroid Gata1 expression. Blood. 2010;115:3463–71.
Li X, Lu YC, Dai K, Torregroza I, Hla T, Evans T. Elavl1a regulates zebrafish erythropoiesis via posttranscriptional control of gata1. Blood. 2014;123:1384–92.
Yang F, Hu H, Liu Y, Shao M, Shao C, Gong Y. Cul4a promotes zebrafish primitive erythropoiesis via upregulating scl and gata1 expression. Cell Death Dis. 2019;10:388.
Krzeminski P, Misiewicz I, Pomorski P, Kasprzycka-Guttman T, Baranska J. Mitochondrial localization of P2Y1, P2Y2 and P2Y12 receptors in rat astrocytes and glioma C6 cells. Brain Res Bull. 2007;71:587–92.
Cattaneo M. New P2Y(12) inhibitors. Circulation. 2010;121:171–9.
Wallentin L, Becker RC, Budaj A, Cannon CP, Emanuelsson H, Held C, et al. Ticagrelor versus clopidogrel in patients with acute coronary syndromes. N Engl J Med. 2009;361:1045–57.
Husted S, van Giezen JJ. Ticagrelor: the first reversibly binding oral P2Y12 receptor antagonist. Cardiovasc Ther. 2009;27:259–74.
Acknowledgements
We thank Dr Nathan Lawson (University of Massachusetts Medical School, Worcester, USA) for kindly providing the Tg(Fli1:egfp) y1 line. We thank Dr Jiu-lin Du (Institute of Neuroscience, CAS, Shanghai, China) for kindly providing the Tg(Gata1:egfp) transgenic lineand Dr Jing-wei Xiong (Peking University, Beijing, China) for providing the plasmids of the CRISPR/Cas9 system. We thank all members of Dr Jing’s lab for helpful discussions and comments on this article. We thank Min Deng and Mei Dong for technical assistance and Sheng-rong Yang for zebrafish husbandry. This work was supported in part by the National Key Research and Development Program of China (2019YFA0802700, 2017YFA0103700), the National Natural Science Foundation of China (91739301, 91339205, 81130005).
Author information
Authors and Affiliations
Contributions
QJ and FFL conceived and designed the project. FFL carried out the experiments and analyzed the data. YLL contributed to confirming and maintaining the knockout line and helped to confirm the phenotype and analyze the data. JC and XSH directed the FISH analysis and drew the heatmap. YNG and PW carried out bioinformatics analysis for MS. XXZ supportedthe zebrafish platform. FFL drafted the manuscript. QJ supervised the study and edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Supplementary information
Rights and permissions
About this article

Cite this article
Li, Ff., Liang, Yl., Han, Xs. et al. ADP receptor P2y12 prevents excessive primitive hematopoiesis in zebrafish by inhibiting Gata1. Acta Pharmacol Sin 42, 414–421 (2021). https://doi.org/10.1038/s41401-020-0431-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41401-020-0431-5
Keywords
This article is cited by
-
Zebrafish: a convenient tool for myelopoiesis research
Cell Regeneration (2023)
