
Abstract
Decline in cognitive function is the most feared aspect of ageing. Poorer midlife cognitive function is associated with increased dementia and stroke risk. The mechanisms underlying variation in cognitive function are uncertain. Here, we assessed associations between 1160 proteins’ plasma levels and two measures of cognitive function, the digit symbol substitution test (DSST) and the Montreal Cognitive Assessment in 1198 PURE-MIND participants. We identified five DSST performance-associated proteins (NCAN, BCAN, CA14, MOG, CDCP1), with NCAN and CDCP1 showing replicated association in an independent cohort, GS (N = 1053). MRI-assessed structural brain phenotypes partially mediated (8–19%) associations between NCAN, BCAN, and MOG, and DSST performance. Mendelian randomisation analyses suggested higher CA14 levels might cause larger hippocampal volume and increased stroke risk, whilst higher CDCP1 levels might increase intracranial aneurysm risk. Our findings highlight candidates for further study and the potential for drug repurposing to reduce the risk of stroke and cognitive decline.
Similar content being viewed by others


Introduction
Decline in cognitive ability and dementia are the most feared aspects of ageing [1], providing a strong rationale for investigating the mechanisms underlying cognitive function. Poorer cognitive function is associated with a greater risk of Alzheimer’s dementia and stroke [2, 3]. This may be due to reduced “cognitive reserve”, which postulates that lower premorbid cognitive function leads to worse cognitive impairment for a given degree of neuropathology [4]. A better understanding of these mechanisms could inform strategies for the prevention and treatment of dementia and stroke.
Recent studies have highlighted the potential for investigating cognition and structural brain phenotypes through the study of plasma proteins [5,6,7,8,9]. These studies identified associations between cognitive function and proteins involved in biological functions previously implicated in dementia, including synaptic function, inflammation, immune function, and blood-brain barrier integrity [5,6,7, 9]. At the time of carrying out this study, previous studies were limited by a focus on a restricted number of proteins and/or purely observational analyses.
Here, we investigated associations between 1160 plasma proteins and cognitive function in the Prospective Urban and Rural Epidemiology (PURE)-MIND cohort [10], and we sought replication in the independent imaging subsample of the Generation Scotland cohort (henceforth, referred to as “GS”). The proteins assessed represent a wide range of biological processes, permitting a hypothesis-free approach to investigating cognitive function. Subsets of these proteins have been assessed in previous studies, allowing assessment of cross-study replication.
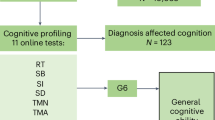
Using a simple measure of processing speed, the digit symbol substitution task (DSST), and a cognitive screening tool, the Montreal Cognitive Assessment (MoCA), we carried out a screen for cognition-associated proteins and then employed mediation analyses to assess the proportion of the protein expression-cognition relationship that could be explained by structural brain phenotypes, including measures of brain volume and white matter hyperintensity (WMH) volume. WMH is an MRI marker of white matter damage and is one of the manifestations of age-related cerebral small vessel disease. Two sample Mendelian randomisation (MR) analyses were performed to assess potentially causal effects of genetically predicted protein levels on genetically predicted cognitive function, brain structure, stroke subtypes, and Alzheimer’s disease (see Fig. 1 for an overview of the study design).

This study involved European (N = 3514), Latin (N = 4309), and Persian (N = 1332) PURE participants for whom genetic and plasma proteomic data were available. Observational analyses to detect plasma biomarkers of cognitive function were performed in the subset of these participants who were enrolled in the PURE-MIND sub-study (N = 1198), for whom plasma protein (N = 1060 proteins) and MRI measurements were available. Mediation analyses were performed to assess whether any observed associations between protein levels and cognitive function were mediated by structural brain phenotypes ascertained by MRI. Finally, two-sample Mendelian randomisation analyses were performed to assess potentially causal effects of genetically-predicted cognition-associated protein levels on genetically-predicted neurological outcomes. For these analyses, genetic instrumental variables for protein levels were identified in the European, Latin, and Persian PURE participants, and associations with neurological outcomes were assessed using external (non-PURE) datasets. Created with BioRender.com.
Results
Cohort characteristics
Key demographic, cognitive, brain MRI and health variables for the participants in PURE-MIND (N = 1198) and GS (N = 1053) are summarised in Table 1. Participants in PURE-MIND were significantly younger than participants in GS (PURE-MIND: mean = 54.5 years (SD = 8.05 years); GS: mean = 59.9 years (SD = 9.59 years); p < 2.2 × 10−16), and a small, but significant, between-cohort difference in DSST score was observed (PURE-MIND: mean = 69.5 (SD = 15.3); GS: mean = 68.1 (SD = 15.2); p = 0.0298). Differences in the levels/types of education received by the two cohorts were observed (p = 5 × 10−4). The two cohorts also differed significantly on several brain volume measurements: PURE-MIND participants have a smaller total brain volume (PURE-MIND: mean = 1058 cm3 (SD = 110 cm3); GS: mean = 1069 cm3 (SD = 109 cm3); p = 0.0212), cerebral white matter volume (PURE-MIND: mean = 447 cm3 (SD = 60.5 cm3); GS: mean = 455 cm3 (SD = 56.9 cm3); p = 1.87 × 10−3), and hippocampal volume (PURE-MIND: mean = 3.92 cm3 (SD = 0.433 cm3); GS: mean = 4.18 cm3 (SD = 0.437 cm3); p = < 2.2 × 10−16) than GS participants, whilst GS participants have a smaller intracranial volume (ICV) than PURE-MIND participants (PURE-MIND: mean = 1491 cm3 (SD = 159 cm3); GS: mean = 1400 cm3 (SD = 225 cm3); p = < 2.2 × 10−16). The two cohorts also differed significantly on other health-related variables that were not directly assessed in this study (Table 1).
Scores on the DSST were normally distributed in both PURE-MIND and GS (Supplementary Figure 1), but scores on the MoCA showed a leftward skew in PURE-MIND (Supplementary Figure 1).
Identification of protein biomarkers of cognitive function and enrichment analyses
Five proteins were associated with DSST performance in PURE-MIND (Fig. 2; Fig. 3; Supplementary Table 1; Supplementary Figure 2). Higher plasma levels of neurocan (NCAN; β = 2.03 (indicating a 2.03 higher DSST score per a standard deviation higher NCAN level), p = 9.11 × 10−8), brevican (BCAN; β = 1.91, p = 5.56 × 10−7), carbonic anhydrase 14 (CA14; β = 1.90, p = 5.90 × 10−7), and myelin-oligodendrocyte glycoprotein (MOG; β = 1.82, p = 2.29 × 10−6), and lower levels of CUB domain-containing protein 1 (CDCP1; β = −1.57, p = 3.97 × 10−5) were associated with significantly better DSST performance. Adjustment for educational attainment modestly attenuated the effect estimate for all five proteins (Supplementary Table 1). Levels of NCAN, BCAN and MOG were positively correlated (0.251 ≤ r ≥ 0.615; all p < 2.20 × 10−16), while CDCP1 and CA14 expression levels were negatively correlated (r = −0.101, p = 4.82 × 10−4; Supplementary Table 2). Three proteins (NCAN, BCAN and CDCP1) proteins were also measured in GS, of which two replicated their association with DSST performance: NCAN (β = 1.40, p = 1.07 × 10−3) and CDCP1 (β = −1.99, p = 9.21 × 10−6; Fig. 3). MoCA performance was not associated with the level of any protein (all p ≥ 2.34 × 10−4; Supplementary Table 3; Supplementary Figure 3). When considering the 129 proteins that were nominally significantly associated (p < 0.05) with DSST score in PURE-MIND and measured in GS (Supplementary Table 4), their effect estimates showed a strong, statistically significant between-cohort correlation (r = 0.625, p = 2.34 × 10−15).

Each protein is represented by a triangle with upwards-facing triangles indicating a positive association with DSST performance and downwards-facing triangles indicating a negative association with DSST performance. The position of each protein on the x-axis is determined by the genomic location of its corresponding gene and the position on the y-axis is determined by the –log10 p-value. The dashed horizontal line indicates the Bonferroni-corrected significance threshold (p = 4.31 × 10−5) required to maintain a 5% type I error rate.

For each protein, the difference in DSST score associated with a standard deviation higher level of protein is shown, together with the 95% confidence interval. Abbreviations: BCAN brevican, CA14 carbonic anhydrase 14, CDCP1 CUB-domain containing protein 1, CI confidence interval, GS Generation Scotland imaging subsample, MOG myelin oligodendrocyte glycoprotein, NCAN neurocan, PURE Prospective Urban and Rural Epidemiology study.
Proteins nominally associated (p < 0.05) with DSST performance (N = 184) were enriched for brain-expressed proteins, most significantly for proteins with hippocampal expression (FDR-corrected p = 0.0154; Supplementary Table 5). Better DSST performance was nominally associated with lower levels of 90 proteins. These proteins mapped to the following immune pathways “interleukin-10 signalling”, “glomerulonephritis”, “regulation of granulocyte chemotaxis”, “positive regulation of leukocyte chemotaxis”, “positive regulation of leukocyte migration”, and “inflammation” (FDR-corrected p ≤ 0.0337; Supplementary Table 6).
Structural brain phenotypes as mediators of protein biomarker-DSST performance associations
In PURE-MIND, better DSST performance was associated with greater cerebral white matter volume (β = 0.0615, p = 4.34 × 10−7), greater total brain volume (β = 0.0349, p = 9.64 × 10−6), greater hippocampal volume (β = 2.97, p = 4.79 × 10−3), and lower log-transformed WMH volume (β = −3.20, p = 1.18 × 10−6). These associations replicated in GS (Supplementary Table 7).
Assessment of the relationships between protein levels and DSST-associated structural brain phenotypes in PURE-MIND revealed systematic differences between those proteins for which higher levels were associated with better DSST performance (NCAN, BCAN, CA14, and MOG), and CDCP1, which was negatively associated with DSST performance (Fig. 4). Whilst NCAN, BCAN, CA14, and MOG showed a positive direction of association with total brain, cerebral white matter, and hippocampal volume measurements and a negative association with WMH volume, the converse was true for CDCP1. The associations between NCAN levels and total brain, cerebral white matter, and hippocampal volumes reached statistical significance (p ≤ 2.56 × 10−5) and were replicated in GS (p ≤ 6.70 × 10−3). BCAN levels were significantly associated with all four brain volumes (p ≤ 4.36 × 10−4), with the associations with total brain and cerebral white matter volumes replicating in GS (p ≤ 3.44 × 10−3). The associations between MOG levels and total brain and cerebral white matter volumes attained statistical significance (p ≤ 2.63 × 10−9), but could not be assessed in GS. We did not identify any significant associations with CA14 or CDCP1 levels after multiple testing correction.

For each protein, the effect estimate (change in brain volume (cm3 or %) per standard deviation increase in protein expression) is shown, together with the 95% confidence interval. Abbreviations: BCAN brevican; CA14 carbonic anhydrase 14, CDCP1 CUB-domain containing protein 1, CI confidence interval, GS Generation Scotland imaging subsample, MOG myelin oligodendrocyte glycoprotein, NCAN neurocan, PURE Prospective Urban and Rural Epidemiology study, WMH white matter hyperintensity.
In PURE-MIND, cerebral white matter volume explained a significant proportion of variance in the relationship between MOG (19.2%), BCAN (14.9%), and NCAN (12.7%) levels and DSST performance (all p < 2 × 10−16) (Supplementary Table 8). After controlling for cerebral white matter volume, the average effect estimates (based on 1000 bootstrap resamples) for these proteins were reduced from 1.81 to 1.47 (MOG), 1.91 to 1.62 (BCAN), and 2.03 to 1.77 (NCAN). Log-transformed WMH volume was a significant partial mediator of the association between BCAN levels and DSST performance (p = 0.002). Controlling for log-transformed WMH resulted in a reduction in the effect estimate from 1.91 to 1.75 (8% mediation).
Identification of potentially causal relationships between protein levels and cognitive function, structural brain phenotypes, and disease outcomes
Inverse variance weighted (IVW) MR analyses were performed to assess the effects of genetically predicted CA14, CDCP1, and MOG levels on cognitive function, structural brain phenotypes, and Alzheimer’s disease, and stroke. Protein quantitative trait loci (pQTLs) located in cis to the genes encoding the proteins-of-interest acted as instrumental variables (IVs) for plasma protein levels (Supplementary Table 9). An insufficient number of pQTLs precluded the assessment of BCAN and NCAN with any of the outcomes-of-interest. For MOG, limited overlap between the pQTLs and SNPs included in the outcome GWASs meant only a subset of the outcomes-of-interest could be assessed.
A one standard deviation higher level of genetically predicted plasma CA14 was associated with a larger hippocampal volume (β = 0.0971 [95% CI: 0.0300 to 0.164], p = 4.58 × 10−3), and a greater risk of all stroke (odds ratio (OR) = 1.08 [95% CI: 1.02 to 1.14], p = 6.97 × 10−3; Supplementary Table 10). A one standard deviation higher level of genetically predicted plasma CDCP1 was associated with an increased risk of intracranial aneurysm (OR = 1.22 [95% CI: 1.02 to 1.47], p = 0.0280). These associations were corroborated by similar effect estimates from the weighted median and MR-RAPS analyses. No evidence of directional or horizontal pleiotropy were observed, and the correct causal direction was assessed. No significant associations were observed between the genetically predicted levels of CA14 or CDCP1 and the risk of Alzheimer’s disease (p ≥ 0.125) (NB. A lack of significant pQTLs precluded the assessment of BCAN, MOG, or NCAN).
Sensitivity analyses were performed in which instrumental variables (IVs) were selected using a stricter threshold for independence. For genetically predicted CA14, these analyses supported the association with hippocampal volume (β = 0.144 [95% CI: 0.0435 to 0.244], p = 4.97 × 10−3), and produced a consistent, although non-significant, effect estimate for the association with risk for all stroke (Supplementary Table 10). For CDCP1, the sensitivity analyses identified a consistent, although non-significant, effect estimate for the association with risk for intracranial aneurysm.
To assess whether between-population heterogeneity in pQTL effects could have affected our findings, we sought to replication using random effects meta-analysis (Supplementary Table 10). For genetically predicted CA14, this approach supported the significant associations with hippocampal volume (β = 0.0953 [95% CI: 0.0586 to 0.132], p = 3.63 × 10−7), and risk for all stroke (OR = 1.06 [95% CI: 1.03 to 1.09], p = 2.56 × 10−4). For genetically predicted CDCP1, it was only possible to assess the association with risk for intracranial aneurysm using a single IV; this identified a significant association (OR = 1.32 [95% CI: 1.08 to 1.61], p = 6.93 × 10−3).
Pairwise Conditional Analysis and Co-localisation Analyses (PWCoCo) were performed to assess the presence of a shared variant for each of the five proteins-of-interest and the same outcomes as assessed by two-sample MR analyses. We were only adequately powered to assess co-localisation between SNPs associated with one pair of traits: MOG plasma level and cognitive function. We did not observe any evidence in support of co-localisation or conditional co-localisation (posterior probability (PP)4/PP3 ≤ 4.81 × 10−4).
Discussion
In this large-scale analysis of the associations between the plasma levels of 1160 proteins and cognitive function, we identify CA14 and CDCP1 as being associated with processing speed, as measured by the DSST, and having potentially causal effects on hippocampal volume and stroke (CA14) and intracranial aneurysm (CDCP1).
Other proteins (BCAN, NCAN, and MOG) were associated with DSST performance and important structural brain phenotypes, with cerebral white matter volume mediating a significant proportion (13-19%) of the relationship between the levels of all three proteins and DSST performance, and WMH volume mediating 8% of the relationship between BCAN levels and DSST performance. A lack of genetic instruments precluded the assessment of potentially causal effects of BCAN and NCAN with any outcome-of-interest, and MOG with several outcomes-of-interest. Enrichment analyses of proteins that were nominally significantly associated with DSST performance revealed a significant enrichment for brain-expressed proteins.
There were no significant associations between plasma protein levels and performance on the MoCA. This might reflect the fact that the MoCA is a screening tool for mild cognitive impairment [11], meaning its sensitivity to detect variation in cognitive function in non-clinical groups is likely to be limited. The maximum MoCA score is 30, and scores higher than 26 indicate normal function. A high mean score (26.5) and a left-skewed distribution indicate a ceiling effect, which likely limited the power to detect associations between protein levels and MoCA score in PURE-MIND.
CA14 is one of fifteen isoforms of the carbonic anhydrase family of zinc metalloprotease enzymes, which catalyse the reversible hydration of carbon dioxide [12]. CA14 is expressed by neurons [13] and involved in regulating extracellular pH following synaptic transmission [14, 15]. Consistent with our findings, acute inhibition of CA14 leads to impaired performance on cognitive tasks in mice [16]. Carbonic anhydrase activation may lead to beneficial cognitive effects in rodents [17]. In keeping with our MR results, there are neuroprotective effects of carbonic anhydrase inhibition in models of amyloidosis, Huntington’s disease, and ischaemic and haemorrhagic stroke [17]. The mechanisms by which carbonic anhydrase inhibition and activation exert their effects are uncertain [16, 17]. FDA-approved carbonic anhydrase inhibitors, and thus the majority of carbonic anhydrase inhibitors investigated to date, are pan-carbonic anhydrase inhibitors. Of the carbonic anhydrase family members measured in our study (CA1, 2, 3, 4, 5A, 6, 9, 12, 13, and 14), only CA14 levels were significantly associated with DSST performance. Further studies are required to determine the therapeutic potential for carbonic anhydrase modulation in the context of cognitive impairment, Alzheimer’s disease, and stroke.
The extracellular matrix (ECM) proteins NCAN and BCAN are brain-specific chondroitin sulphate proteoglycans, which are expressed by neurons and astrocytes (NCAN and BCAN), and oligodendrocytes (BCAN). They contribute to the formation of a specialised structure, the perineuronal net (PNN), which plays a key role in memory and neuronal plasticity, and which is disrupted in Alzheimer’s disease [18]. Our findings are consistent with those of Harris et al. [5], who found plasma levels of NCAN and BCAN were positively associated with brain volume. Plasma levels of both NCAN and BCAN have previously been shown to be positively associated with general cognitive function and DSST performance [9], whilst BCAN levels have been found to be positively associated with Mini Mental State Examination performance and reduced in patients with Alzheimer’s disease or mild cognitive impairment [7]. Mice that are lacking either NCAN or BCAN expression show normal development and memory function but reduced hippocampal long-term potentiation [19, 20], whilst quadruple knock-outs, which lack NCAN, BCAN, and two additional ECM proteins (tenascin-C and tenascin-R) show an altered ratio of excitatory to inhibitory synapses and a reduction in the number and complexity of hippocampal PNNs [21]. Genetic variation in the gene encoding A Disintegrin and Metalloproteinase with Thrombospondin Motifs 4 (ADAMTS4), which degrades the four members of the lectican family (including NCAN and BCAN), has been implicated in Alzheimer’s disease [22]. Taken together, the evidence suggests NCAN, BCAN and their regulators as molecules-of-interest in Alzheimer’s disease.
MOG is an oligodendrocyte-expressed membrane glycoprotein, the exact function of which is unknown [23].
CDCP1 is a widely expressed transmembrane glycoprotein that acts as a ligand for T cell-expressed Cluster of Differentiation 6 (CD6),and is implicated in autoimmune conditions [24]. CDCP1 is amenable to modulation by approved drug treatments: Itolizumab, which is used to treat psoriasis, disrupts CDCP1-CD6 binding and downregulates T-cell-mediated inflammation [25], whilst atomoxetine, a treatment for attention deficit hyperactivity disorder, which is being considered for the treatment of mild cognitive impairment, reduced cerebrospinal fluid (CSF) CDCP1 levels [26]. Intriguingly, findings in mice suggest a functional link between CDCP1 and MOG [6].
Our study has several strengths. We measured 1160 proteins, associated with a wide range of physiological processes, in a large, well-characterised cohort. Replication analyses, where possible, were performed in an independent cohort in which proteins were measured using an independent methodology. The availability of genetic and brain MRI data permitted an exploration of causality and putative causal pathways. The use of MR to identify potentially causal associations will have offered protection against some of the common confounders of observational analyses [27], with the use of multiple MR methods, which generally gave concordant estimates of effect, mitigating against the individual biases of different MR methodologies [28]. Moreover, by requiring instrumental variables to be located in cis to their target protein, we limited the chance of pleiotropic effects [29].
There are also several limitations to consider
First, the 1160 proteins measured represent a small subset of the circulating proteome [30]. Although these proteins are involved in a wide range of biological functions represented by all 13 Olink Target 96 panels, limitations to our understanding of the proteome mean that is not possible to assess the extent to which these proteins are representative of the rest of the proteome. Replication analyses were only performed for those proteins for which data were available in the GS cohort, meaning that we did not assess the replication of CA14 or MOG.
Second, the availability of suitable IVs mean that our primary MR analyses were only performed for CA14, CDCP1, and MOG. Whilst we required a minimum of three IVs for the primary MR analyses, our sensitivity analyses, in which a stricter threshold for independence was applied to the IVs necessitated the use of fewer than three IVs in each analysis. As such, the results of the sensitivity analyses should be interpreted with this caveat in mind.
Third, for all but one pair of traits, we were insufficiently powered to assess co-localisation between genetic variants associated with protein level and cognition, structural brain phenotypes, and disease outcomes. This means that it is possible that significant MR findings might reflect the presence of separate causal variants in linkage disequilibrium (LD) with one another [31]
Fourth, we measured protein levels in the plasma, rather than in the brain or CSF. It is, however, important to note the striking enrichment for brain-expressed proteins amongst the DSST-associated proteins. Previous analyses of the GS cohort, in which replication was sought in the present study, have identified the levels of several plasma proteins as being associated with multiple markers of brain health [8]. These findings support the use of the plasma to assess brain-related phenotypes and emphasise the need for additional research to explain the mechanisms controlling the efflux of brain-expressed proteins into the bloodstream in non-clinical populations. Moreover, the use of cis pQTLs, which are likely to be shared across tissues [32], as IVs in our MR analyses, supports the possibility that the MR-identified associations reflect the actions of the proteins-of-interest in the brain.
In summary, we identified protein biomarkers of cognitive function that may causally affect brain structure and risk for stroke and intracranial aneurysm. Notwithstanding the need for replication, our findings prompt several hypotheses that should be assessed by future studies. Our apparently paradoxical findings of higher CA14 levels being associated with both better cognitive function and increased stroke risk suggest that molecular findings can inform a more nuanced understanding of the relationship between premorbid cognitive function and neurological disease risk. It is possible that improved risk stratification may be achieved through the combination of cognitive assessment and biomarker measurement. The availability of approved drugs targeting our identified proteins raises the possibility of drug repurposing for novel therapeutic interventions to prevent cognitive decline, stroke, and intracranial aneurysm.
Methods
Sample information
This study used data from participants of self-reported European (N = 3514), Latin (N = 4309), or Persian (N = 1332) ancestry from the Population Urban Rural Epidemiology (PURE) biomarker sub-study [33] (Supplementary Information). African (N = 659), South Asian (N = 604), East Asian (N = 314), and Arab (N = 204) participants were excluded to align PURE genetic data with external genetic datasets, which are predominantly European. Participants of Latin and Persian ancestry were included due to their genetic overlap with European participants [33].
The PURE biomarker study also included European participants enrolled in PURE-MIND (N = 1198) [10] (Supplementary Information).
The European, Latin, and Persian PURE biomarker cohort participants were used to identify protein biomarker pQTLs [33], for use in MR analyses, while data from the European PURE-MIND biomarker participants were used for observational association analyses.
We sought replication of our observational findings in GS [34, 35], which was recruited through re-contact of the Generation Scotland: Scottish Family Health Study (GS:SFHS) [36, 37]. GS:SFHS is a population- and family-based cohort of >24,000 individuals from Scotland. GS:SFHS participants were recruited between 2006 and 2011. Upon recruitment, participants attended a clinic where detailed health, cognitive, and lifestyle information, and biological samples were collected. Between 2015 and 2018, a subset of the GS:SFHS participants completed additional health and cognitive assessments, brain MRI, and provided blood samples for proteomic analysis. Up to 1053 GS participants were available for replication analyses.
The GS:SFHS obtained ethical approval from the NHS Tayside Committee on Medical Research Ethics, on behalf of the National Health Service (reference: 05/S1401/89). All participants provided broad and enduring written informed consent for biomedical research. GS:SFHS has Research Tissue Bank Status (reference: 15/ES/0040), providing generic ethical approval for a wide range of uses within medical research. The imaging subsample of GS:SFHS (referred to as “GS” herein) received ethical approval from the NHS Tayside committee on research ethics (reference 14/SS/0039). All experimental methods were in accordance with the Helsinki Declaration.
Assessment of between-cohort differences
Between cohort differences in quantitative variables (age, DSST score, BMI, blood pressure, and brain MRI volumes) were assessed using two-sample t-tests. Categorical variables (sex, education type, and disease status) where the smallest count in any cell of the contingency table was greater than five were assessed using a chi-squared test; otherwise, a Fisher’s Exact Test was employed. Statistical significance was defined as p < 0.05.
Assessment of cognitive function
General cognitive ability was measured in PURE-MIND and GS by trained assessors using the DSST (Wechsler Adult Intelligence Scale, 3rd Edition) [38]. The DSST is a pencil and paper test in which participants must match symbols to numbers according to a key. Participants were scored according to the number of correct matches made within two minutes (maximum score: 133). The DSST measures several cognitive functions, including associative learning and executive function [39], and DSST performance is highly correlated with the general intelligence factor, g. PURE-MIND participants completed the Montreal Cognitive Assessment (MoCA) [11], a questionnaire-based test with scores 0 to 30. A score of 26 or higher is considered normal [11].
Measurement of plasma protein expression
In the PURE biomarker cohort, 1196 plasma protein levels were measured by proximity extension assay using the Olink Proseek Target 96 reagent kit (Olink, Uppsala, Sweden) in 12066 participants (including 3735 European, 4695 Latin, and 1436 Persian). Following pre-processing and quality control steps (Supplementary Information), measurements were available for 1160 biomarkers in 8369-9154 European, Latin, or Persian participants (depending on biomarker-specific missingness).
In GS, plasma protein levels were measured with the SOMAscan assay platform (SomaLogic Inc.), as described previously [40]. Following initial data processing and quality control steps, measures of 4058 proteins were available in 1095 participants. Prior to analysis, protein abundance measurements were log-transformed and rank-based inverse normalised.
Brain imaging
PURE-MIND participants enrolled in the PURE biomarker cohort were scanned at four sites in Canada (three at 1.5 T (two on General Electric (GE) scanners, one on a Phillips scanner), one at 3 T (GE)). The brain imaging phenotypes assessed in this study were total brain volume (excluding ventricles), total white matter volume, hippocampal volume, average cortical thickness, a multi-region composite thickness measure designed to differentiate Alzheimer’s disease patients from clinically normal participants [41], silent brain infarcts (SBI), cerebral microbleeds (CMB), and WMH volumes. These will henceforth be referred to as the “structural brain phenotypes”. Further information about the derivation of the structural brain phenotypes is available in the Supplementary Information.
Genotyping and imputation of PURE-MIND
PURE participant genotypes (Thermofisher Axiom Precision Medicine Research Array r.3) were called using Axiom Power Tools and in-house scripts. Quality control steps are described in the Supplementary Information.
Imputation was performed on the 749,783 genotyped variants following the TOPMed Imputation server pipeline (https://imputation.biodatacatalyst.nhlbi.nih.gov/). Further details are in the Supplementary Information.
Assessment of the association between protein biomarkers and cognitive function and structural imaging phenotypes
We assessed the association between standardised protein levels and cognitive and structural brain phenotypes using two-tailed linear (DSST, MoCA, total brain volume, white matter volume, hippocampal volume, WMH volume, cortical thickness) or logistic (CMB, SBI) regression. The cognitive or structural brain phenotype-of-interest was the dependent variable with the standardised protein expression level, age, age2, sex, and the first ten genetic principal components as independent variables. A sensitivity analysis was performed for DSST-associated proteins in which we further adjusted for education (a categorical variable with levels: (i) no education; (ii) high school or less; (iii) trade school; and (iv) college or university). We calculated Pearson’s correlation coefficient to assess the pairwise correlations between DSST-associated proteins. Within each analysis, we applied a Bonferroni correction to determine statistical significance, yielding the following significance thresholds: p < 4.31 ×10−5 when assessing associations with 1160 proteins; p < 2.5 × 10−3 when assessing associations with the five DSST-associated proteins across four DSST-associated structural brain phenotypes; and p < 5 × 10−3 when assessing 10 pairwise correlations between proteins.
We performed replication analyses in GS for the significant proteins identified in PURE-MIND. Two-tailed mixed-effects models were fitted using the lmekin function from the R package coxme v.2.2.17 [42] to assess the association of the outcome variable (DSST performance, total brain volume (excluding ventricles), cerebral white matter volume, hippocampal volume, and WMH volume) with standardised protein expression, covarying for age, age2, sex, study site (Dundee or Aberdeen), the delay between blood sampling and protein extraction, depression (a binary variable representing lifetime depression status), and a kinship matrix. When a brain volume phenotype was the outcome variable, additional covariates were included to account for ICV, the interaction between ICV and the study site (to account for a site-associated batch effect on ICV measurement), and whether there was manual intervention using tools within Freesurfer during the quality control process. Replication was defined as a concordant direction of effect, meeting a Bonferroni-corrected threshold of p < 1.67 × 10−2 (accounting for the assessment of three DSST-associated proteins) or p < 7.14 × 10−3 (accounting for the assessment of seven structural brain phenotype-protein combinations).
Assessment of the association of DSST performance with MRI-derived structural brain phenotypes
To identify mediators of the association between protein expression and DSST performance, we first established the structural brain phenotypes that satisfied the requirements of potential mediators (i.e. associated with both DSST performance and at least one DSST-associated protein), and then formally tested the meditation relationship by bootstrap mediation analyses.
We estimated the association between DSST performance and structural brain phenotypes in PURE-MIND using linear models. All brain volume measurements were normalised to ICV and the models included covariates for age, age2, sex, and the first ten genetic principal components. We defined statistical significance as p < 0.00625 (Bonferroni correction for eight phenotypes; two-tailed) and sought replication of significant associations (N = 5) in GS. In GS, brain volumes were residualised for ICV, scanner location, the interaction between ICV and scanner location, and whether there was manual intervention during the quality control process. The resultant residuals were included as the dependent variable in a mixed effects model with DSST score, age, age2, sex, depression, and a kinship matrix as independent variables. Statistical significance was defined as p < 0.0125 (two-tailed).
The DSST performance-associated brain MRI phenotypes (N = 4) were assessed as potential mediators of the protein level-DSST associations (N = 3, yielding a total of N = 9 mediations to assess) using bootstrap mediation analysis in PURE-MIND. Analyses were performed using the R package “mediation” [43] with 1000 bootstraps. We corrected for the nine potential mediation relationships assessed using a Bonferroni-corrected threshold of p < 5.56 × 10−3.
Functional and tissue-specific expression enrichment analyses
Proteins associated with DSST performance at p < 0.05 in PURE-MIND were included in functional and tissue-specific expression analyses in three groups: (i) all proteins; (ii) positively associated proteins; and (iii) negatively associated proteins. Enrichment was assessed relative to all proteins in our dataset that passed quality control (N = 1160). Functional enrichment analyses were performed using WebGestalt (http://www.webgestalt.org/) [44] using default parameter settings for the over-representation analysis method to assess enrichment for: (i) gene ontology categories (biological processes, molecular functions, and cellular compartments); (ii) Reactome pathways; and (iii) disease-associated genes (Disgenet). Tissue-specific enrichment analyses were performed using the “GTEx v8: 54 tissue types” and “GTEx v8: 30 general tissue types” gene expression datasets in FUnctional Mapping and Annotation (FUMA) [45]. For both the functional enrichment and tissue expression analyses, enrichment was assessed using a hypergeometric test and significant enrichment was defined as a Benjamini-Hochberg-adjusted p < 0.05, correcting for the number of tests performed within each analysis platform. Analyses were performed using web interfaces accessed on 18/04/2022 (WebGestalt and FUMA) and 14/01/2023 (FUMA).
Two-sample forward MR analyses
We performed two-sample forward MR analyses to identify potentially causal associations between genetically predicted plasma protein levels and: (i) cognitive function; (ii) structural brain phenotypes (total brain volume, cerebral white matter volume, hippocampal volume, WMH volume, and CMB); and (iii) disease outcomes (Alzheimer’s disease, all stroke, stroke subtypes (ischaemic, cardioembolic, large artery, and small vessel), and intracranial aneurysm).
Associations between single nucleotide polymorphisms (SNPs) and plasma protein expression levels were calculated in PURE (Supplemental Information). Following quality control, a set of pQTLs that was independent (r2 < 0.1) in all three populations was retained. Sensitivity analyses were performed in which the pruning threshold was adjusted to r2 < 0.01.
The independent set of pQTLs were assessed for their associations with cognitive function, structural brain phenotypes, and disease outcomes using summary statistics from published studies [46,47,48,49,50,51,52,53].
MR analyses were performed using the R packages MRBase for TwoSample MR v.0.5.6 [54], mr.raps v.0.4.1 [55], and MRPRESSO v.1.0 [56]. We employed several complementary MR approaches: IVW [57], weighted median [58], robust adjusted profile scores (RAPS) [55], MR-Egger [59], and MR-PRESSO [56]. We adopted the IVW approach as our primary methodology and defined statistical significance using a liberal within-outcome variable Bonferroni correction for the proteins (CA14, CDCP1, and MOG) that could be assessed, yielding a significance threshold of p < 0.0167 (or p < 0.025 or 0.05 when an outcome could only be assessed for two or one protein(s)). Further details of the MR analyses are included in the Supplementary Information.
Pairwise conditional analysis and co-localisation analysis (PWCoCo)
PWCoCo [31, 60] was performed to assess the existence of a shared causal variant between (i) pQTLs for each of the five proteins-of-interest and (ii) variants associated with the outcomes assessed in the two-sample MR analyses (Supplementary Information).
Software
Statistical analyses and plot generation were performed in R (versions 3.6.0, 4.1.1, 4.1.2, 4.2.0, 4.2.1).
Data availability
The terms of consent for PURE participants preclude the sharing of individual-level data. Individual level data is available through collaboration with PURE researchers (https://www.phri.ca/research/pure/). Summary-statistics for the analyses presented here are available in the supplementary materials. According to the terms of consent for GS participants, applications for individual-level data must be reviewed by the GS Access Committee (access@generationscotland.org). Complete summary statistics are available in the supplementary materials for the protein-DSST score associations assessed in this study.
Code availability
The code used to generate the results in this study is available on reasonable request from the corresponding author.
References
Martin GM. Defeating Dementia. Nature. 2004;431:247–8.
Rostamian S, Mahinrad S, Stijnen T, Sabayan B, de Craen AJ. Cognitive impairment and risk of stroke: a systematic review and meta-analysis of prospective cohort studies. Stroke. 2014;45:1342–8.
Valenzuela MJ, Sachdev P. Brain reserve and dementia: a systematic review. Psychol Med. 2006;36:441–54.
Pettigrew C, Soldan A. Defining Cognitive Reserve and Implications for Cognitive Aging. Curr Neurol Neurosci Rep. 2019;19:1.
Harris SE, Cox SR, Bell S, Marioni RE, Prins BP, Pattie A, et al. Neurology-related protein biomarkers are associated with cognitive ability and brain volume in older age. Nat Commun. 2020;11:800.
Lindbohm JV, Mars N, Walker KA, Singh-Manoux A, Livingston G, Brunner EJ, et al. Plasma proteins, cognitive decline, and 20-year risk of dementia in the Whitehall II and Atherosclerosis Risk in Communities studies. Alzheimers Dement. 2022;18:612–24.
Whelan CD, Mattsson N, Nagle MW, Vijayaraghavan S, Hyde C, Janelidze S, et al. Multiplex proteomics identifies novel CSF and plasma biomarkers of early Alzheimer’s disease. Acta Neuropathol Commun. 2019;7:169.
Gadd DA, Hillary RF, McCartney DL, Shi L, Stolicyn A, Robertson NA, et al. Integrated methylome and phenome study of the circulating proteome reveals markers pertinent to brain health. Nat Commun. 2022;13:4670.
Tin A, Fohner AE, Yang Q, Brody JA, Davies G, Yao J, et al. Identification of circulating proteins associated with general cognitive function among middle-aged and older adults. Communications Biology. 2023;6:1117.
Smith EE, O’Donnell M, Dagenais G, Lear SA, Wielgosz A, Sharma M, et al. Early cerebral small vessel disease and brain volume, cognition, and gait. Ann Neurol. 2015;77:251–61.
Nasreddine ZS, Phillips NA, Bedirian V, Charbonneau S, Whitehead V, Collin I, et al. The Montreal Cognitive Assessment, MoCA: a brief screening tool for mild cognitive impairment. J Am Geriatr Soc. 2005;53:695–9.
Lindskog S. Structure and mechanism of carbonic anhydrase. Pharmacol Ther. 1997;74:1–20.
Parkkila S, Parkkila AK, Rajaniemi H, Shah GN, Grubb JH, Waheed A, et al. Expression of membrane-associated carbonic anhydrase XIV on neurons and axons in mouse and human brain. Proc Natl Acad Sci USA. 2001;98:1918–23.
Chen JC, Chesler M. pH transients evoked by excitatory synaptic transmission are increased by inhibition of extracellular carbonic anhydrase. Proc Natl Acad Sci USA. 1992;89:7786–90.
Shah GN, Ulmasov B, Waheed A, Becker T, Makani S, Svichar N, et al. Carbonic anhydrase IV and XIV knockout mice: roles of the respective carbonic anhydrases in buffering the extracellular space in brain. Proc Natl Acad Sci USA. 2005;102:16771–6.
Provensi G, Carta F, Nocentini A, Supuran CT, Casamenti F, Passani MB, et al. A New Kid on the Block? Carbonic Anhydrases as Possible New Targets in Alzheimer’s Disease. Int J Mol Sci. 2019;20:4724.
Lemon N, Canepa E, Ilies MA, Fossati S. Carbonic Anhydrases as Potential Targets Against Neurovascular Unit Dysfunction in Alzheimer’s Disease and Stroke. Front Aging Neurosci. 2021;13:772278.
Sorg BA, Berretta S, Blacktop JM, Fawcett JW, Kitagawa H, Kwok JC, et al. Casting a Wide Net: Role of Perineuronal Nets in Neural Plasticity. J Neurosci. 2016;36:11459–68.
Brakebusch C, Seidenbecher CI, Asztely F, Rauch U, Matthies H, Meyer H, et al. Brevican-deficient mice display impaired hippocampal CA1 long-term potentiation but show no obvious deficits in learning and memory. Mol Cell Biol. 2002;22:7417–27.
Zhou XH, Brakebusch C, Matthies H, Oohashi T, Hirsch E, Moser M, et al. Neurocan is dispensable for brain development. Mol Cell Biol. 2001;21:5970–8.
Gottschling C, Wegrzyn D, Denecke B, Faissner A. Elimination of the four extracellular matrix molecules tenascin-C, tenascin-R, brevican and neurocan alters the ratio of excitatory and inhibitory synapses. Sci Rep. 2019;9:13939.
Marioni RE, Harris SE, Zhang Q, McRae AF, Hagenaars SP, Hill WD, et al. GWAS on family history of Alzheimer’s disease. Transl Psychiatry. 2018;8:99.
Peschl P, Bradl M, Hoftberger R, Berger T, Reindl M. Myelin Oligodendrocyte Glycoprotein: Deciphering a Target in Inflammatory Demyelinating Diseases. Front Immunol. 2017;8:529.
Enyindah-Asonye G, Li Y, Ruth JH, Spassov DS, Hebron KE, Zijlstra A, et al. CD318 is a ligand for CD6. Proc Natl Acad Sci USA. 2017;114:E6912–E6921.
Dogra S, Uprety S, Suresh SH. Itolizumab, a novel anti-CD6 monoclonal antibody: a safe and efficacious biologic agent for management of psoriasis. Expert Opin Biol Ther. 2017;17:395–402.
Levey AI, Qiu D, Zhao L, Hu WT, Duong DM, Higginbotham L, et al. A phase II study repurposing atomoxetine for neuroprotection in mild cognitive impairment. Brain. 2022;145:1924–38.
Lawlor DA, Harbord RM, Sterne JA, Timpson N, Davey Smith G. Mendelian randomization: using genes as instruments for making causal inferences in epidemiology. Stat Med. 2008;27:1133–63.
Slob EAW, Burgess S. A comparison of robust Mendelian randomization methods using summary data. Genet Epidemiol. 2020;44:313–29.
Swerdlow DI, Kuchenbaecker KB, Shah S, Sofat R, Holmes MV, White J, et al. Selecting instruments for Mendelian randomization in the wake of genome-wide association studies. Int J Epidemiol. 2016;45:1600–16.
Omenn GS, Lane L, Overall CM, Corrales FJ, Schwenk JM, Paik YK, et al. Progress on Identifying and Characterizing the Human Proteome: 2018 Metrics from the HUPO Human Proteome Project. J Proteome Res. 2018;17:4031–41.
Zheng J, Haberland V, Baird D, Walker V, Haycock PC, Hurle MR, et al. Phenome-wide Mendelian randomization mapping the influence of the plasma proteome on complex diseases. Nat Genet. 2020;52:1122–31.
Yang C, Farias FHG, Ibanez L, Suhy A, Sadler B, Fernandez MV, et al. Genomic atlas of the proteome from brain, CSF and plasma prioritizes proteins implicated in neurological disorders. Nat Neurosci. 2021;24:1302–12.
Narula S, Yusuf S, Chong M, Ramasundarahettige C, Rangarajan S, Bangdiwala SI, et al. Plasma ACE2 and risk of death or cardiometabolic diseases: a case-cohort analysis. Lancet. 2020;396:968–76.
Habota T, Sandu AL, Waiter GD, McNeil CJ, Steele JD, Macfarlane JA, et al. Cohort profile for the STratifying Resilience and Depression Longitudinally (STRADL) study: A depression-focused investigation of Generation Scotland, using detailed clinical, cognitive, and neuroimaging assessments. Wellcome Open Res. 2021;4:185.
Navrady LB, Wolters MK, MacIntyre DJ, Clarke TK, Campbell AI, Murray AD, et al. Cohort Profile: Stratifying Resilience and Depression Longitudinally (STRADL): a questionnaire follow-up of Generation Scotland: Scottish Family Health Study (GS:SFHS). Int J Epidemiol. 2018;47:13–14g.
Smith BH, Campbell A, Linksted P, Fitzpatrick B, Jackson C, Kerr SM, et al. Cohort Profile: Generation Scotland: Scottish Family Health Study (GS:SFHS). The study, its participants and their potential for genetic research on health and illness. Int J Epidemiol. 2013;42:689–700.
Smith BH, Campbell H, Blackwood D, Connell J, Connor M, Deary IJ, et al. Generation Scotland: the Scottish Family Health Study; a new resource for researching genes and heritability. BMC Med Genet. 2006;7:74.
Wechsler, D, Wechsler Adult Intelligence Scale-Third Edition (WAIS-III). 1997, San Antonio: Harcourt Assessment Inc.
Jaeger J. Digit Symbol Substitution Test: The Case for Sensitivity Over Specificity in Neuropsychological Testing. J Clin Psychopharmacol. 2018;38:513–9.
Shi L, Buckley NJ, Bos I, Engelborghs S, Sleegers K, Frisoni GB, et al. Plasma Proteomic Biomarkers Relating to Alzheimer’s Disease: A Meta-Analysis Based on Our Own Studies. Front Aging Neurosci. 2021;13:712545.
Schwarz CG, Gunter JL, Wiste HJ, Przybelski SA, Weigand SD, Ward CP, et al. A large-scale comparison of cortical thickness and volume methods for measuring Alzheimer’s disease severity. Neuroimage Clin. 2016;11:802–12.
Therneau, TM, coxme: mixed effects Cox models. 2012.
Tingley D, Yamamoto T, Hirose K, Keele L, Imai K. mediation: R Package for Causal Mediation Analysis. Journal of Statistical Software. 2014;59:e9034.
Liao Y, Wang J, Jaehnig EJ, Shi Z, Zhang B. WebGestalt 2019: gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019;47:W199–W205.
Watanabe K, Taskesen E, van Bochoven A, Posthuma D. Functional mapping and annotation of genetic associations with FUMA. Nat Commun. 2017;8:1826.
Savage JE, Jansen PR, Stringer S, Watanabe K, Bryois J, de Leeuw CA, et al. Genome-wide association meta-analysis in 269,867 individuals identifies new genetic and functional links to intelligence. Nat Genet. 2018;50:912–9.
Smith SM, Douaud G, Chen W, Hanayik T, Alfaro-Almagro F, Sharp K, et al. An expanded set of genome-wide association studies of brain imaging phenotypes in UK Biobank. Nat Neurosci. 2021;24:737–45.
Hibar DP, Adams HHH, Jahanshad N, Chauhan G, Stein JL, Hofer E, et al. Novel genetic loci associated with hippocampal volume. Nat Commun. 2017;8:13624.
Persyn E, Hanscombe KB, Howson JMM, Lewis CM, Traylor M, Markus HS. Genome-wide association study of MRI markers of cerebral small vessel disease in 42,310 participants. Nat Commun. 2020;11:2175.
Knol MJ, Lu D, Traylor M, Adams HHH, Romero JRJ, Smith AV, et al. Association of common genetic variants with brain microbleeds: A genome-wide association study. Neurology. 2020;95:e3331–e3343.
Bellenguez C, Kucukali F, Jansen IE, Kleineidam L, Moreno-Grau S, Amin N, et al. New insights into the genetic etiology of Alzheimer’s disease and related dementias. Nat Genet. 2022;54:412–36.
Mishra A, Malik R, Hachiya T, Jurgenson T, Namba S, Posner DC, et al. Stroke genetics informs drug discovery and risk prediction across ancestries. Nature. 2022;611:115–23.
Bakker MK, van der Spek RAA, van Rheenen W, Morel S, Bourcier R, Hostettler IC, et al. Genome-wide association study of intracranial aneurysms identifies 17 risk loci and genetic overlap with clinical risk factors. Nat Genet. 2020;52:1303–13.
Hemani G, Zheng J, Elsworth B, Wade KH, Haberland V, Baird D, et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife. 2018;7:e34408.
Zhao Q, Wang J, Hemani G, Bowden J, Small DS. Statistical inference in two-sample summary-data Mendelian randomization using robust adjusted profile score. The. Annals of Statistics. 2020;48:1742–69.
Verbanck M, Chen CY, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat Genet. 2018;50:693–8.
Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genet Epidemiol. 2013;37:658–65.
Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent Estimation in Mendelian Randomization with Some Invalid Instruments Using a Weighted Median Estimator. Genet Epidemiol. 2016;40:304–14.
Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017;32:377–89.
Robinson, JW, G Hemani, MS Babaei, Y Huang, DA Baird, EA Tsai, et al. An efficient and robust tool for colocalisation: Pair-wise Conditional and Colocalisation (PWCoCo). bioRxiv, 2022: 2022.08.08.503158.
Acknowledgements
We are grateful to all the families who took part in Generation Scotland, the general practitioners and the Scottish School of Primary Care for their help in recruiting them, and the whole Generation Scotland team, which includes interviewers, computer and laboratory technicians, clerical workers, research scientists, volunteers, managers, receptionists, healthcare assistants and nurses. We thank Dr Alison Offer for assistance in producing the forest plots.
Funding
PURE study: The PURE study is an investigator-initiated study that is funded by the Population Health Research Institute, the Canadian Institutes of Health Research (CIHR), Heart and Stroke Foundation of Ontario, support from CIHR’s Strategy for Patient Oriented Research, through the Ontario SPOR Support Unit, as well as the Ontario Ministry of Health and Long-Term Care and through unrestricted grants from several pharmaceutical companies (with major contributions from AstraZeneca [Canada], Sanofi-Aventis [France and Canada], Boehringer Ingelheim [Germany and Canada], Servier, and GlaxoSmithKline), and additional contributions from Novartis and King Pharma and from various national or local organisations in participating countries as follows: Argentina—Fundacion ECLA; Bangladesh—Independent University, Bangladesh and Mitra and Associates; Brazil—Unilever Health Institute, Brazil; Canada—Public Health Agency of Canada and Champlain Cardiovascular Disease Prevention Network; Chile—Universidad de la Frontera; Colombia—Colciencias (grant number 6566–04–18062); South Africa—The North-West University, SANPAD (SA and Netherlands Programme for Alternative Development), National Research Foundation, Medical Research Council of South Africa, The South Africa Sugar Association, Faculty of Community and Health Sciences; Sweden—grants from the Swedish State under the Agreement concerning research and education of doctors, the Swedish Heart and Lung Foundation, the Swedish Research Council, the Swedish Council for Health, Working Life and Welfare, King Gustaf V’s and Queen Victoria Freemasons Foundation, AFA Insurance, Swedish Council for Working Life and Social Research, Swedish Research Council for Environment, Agricultural Sciences and Spatial Planning, grant from the Swedish State under (LäkarUtbildningsAvtalet) Agreement, and grant from the Västra Götaland Region; and United Arab Emirates—Sheikh Hamdan Bin Rashid Al Maktoum Award for Medical Sciences, Dubai Health Authority, Dubai. The PURE biomarker project was supported by Bayer and the CIHR. The biomarker project was led by PURE investigators at the Population Health Research Institute (Hamilton, Canada) in collaboration with Bayer scientists. Bayer directly compensated the Population Health Research Institute for measurement of the biomarker panels, scientific, methodological, and statistical work. Genetic analyses were supported by CIHR (G-18–0022359) and Heart and Stroke Foundation of Canada (application number 399497) in the form of funding to GP. GS: This work was supported by the Wellcome Trust [104036/Z/14/Z, 220857/Z/20/Z, and 216767/Z/19/Z] and an MRC Mental Health Data Pathfinder Grant [MC_PC_17209] to AMM. DAG is funded by the Wellcome Trust Translational Neuroscience PhD Programme at the University of Edinburgh [108890/Z/15/Z]. LS and ANH are supported by Medical Research Council [MR/L023784/2]: Dementias Platform UK. LS is also supported by a Medical Research Council Award to the University of Oxford [MC_PC_17215]. AS is supported through the Wellcome-University of Edinburgh Institutional Strategic Support Fund (Reference 204804/Z/16/Z), and indirectly through the Lister Institute of Preventive Medicine award with reference 173096. Generation Scotland received core support from the Chief Scientist Office of the Scottish Government Health Directorates [CZD/16/6] and the Scottish Funding Council [HR03006]. Genotyping of the GS:SFHS samples was carried out by the Genetics Core Laboratory at the Clinical Research Facility, Edinburgh, Scotland, and was funded by the UK’s Medical Research Council and the Wellcome Trust [104036/Z/14/Z].
Author information
Authors and Affiliations
Contributions
Study conception and design: RMW, WNW, GP; data analysis: RMW, MC, NP; drafting the article: RMW, WNW, GP; data preparation: MC, NP, MP, DAG, AC, HCW, AS, LS, XS, EE, MOD; data collection: ANH, AMM, EE, SH, SR, MOD, SY, WNW, GP; revision of the article: RMW, MC, NP, MP; WNW, GP; all authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
MC is supported by a Canadian Institute of Health Research doctoral award and has received consulting fees from Bayer AG. MP is supported by the EJ Moran Campbell Internal Career Research Award from McMaster University. DAG is a part-time employee of Optima partners, a health data consultancy based at the Bayes centre, The University of Edinburgh. SH is an employee of Bayer AG. AMM has previously received speaker’s fees from Illumina and Janssen and research grant funding from The Sackler Trust. SY is supported by the Heart and Stroke Foundation/Marion W Burke Chair in Cardiovascular Disease. GP is supported by the CISCO Professorship in Integrated Health Systems. The other authors declare no competing interests.
Ethical approval
PURE study: All centres contributing to PURE were required to obtain approval from their respective ethics committees (Institutional Review Boards). Participant data is confidential and only authorized individuals can access study-related documents. The participants’ identities are protected in documents transmitted to the Coordinating Office, as well as biomarker and genetic data. Participants provided informed consent to obtain baseline information, and to collect and store genetic and other biological specimens.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Walker, R.M., Chong, M., Perrot, N. et al. The circulating proteome and brain health: Mendelian randomisation and cross-sectional analyses. Transl Psychiatry 14, 204 (2024). https://doi.org/10.1038/s41398-024-02915-x
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41398-024-02915-x
