
Abstract
The serotonin 5-HT1A receptor has attracted wide attention as a target for treatment of psychiatric disorders. Although this receptor is important in the pharmacological mechanisms of action of new-generation antipsychotics, its characterization remains incomplete. Studies based on in vitro molecular imaging on brain tissue by autoradiography, and more recently in vivo PET imaging, have not yielded clear results, in particular due to the limitations of current 5-HT1A radiotracers, which lack specificity and/or bind to all 5-HT1A receptors, regardless of their functional status. The new concept of PET neuroimaging of functionally active G-protein-coupled receptors makes it possible to revisit PET brain exploration by enabling new research paradigms. For the 5-HT1A receptor it is now possible to use [18F]-F13640, a 5-HT1A receptor radioligand with high efficacy agonist properties, to specifically visualize and quantify functionally active receptors, and to relate this information to subjects’ pathophysiological or pharmacological state. We therefore propose imaging protocols to follow changes in the pattern of functional 5-HT1A receptors in relation to mood deficits or cognitive processes. This could allow improved discrimination of different schizophrenia phenotypes and greater understanding of the basis of therapeutic responses to antipsychotic drugs. Finally, as well as targeting functionally active receptors to gain insights into the role of 5-HT1A receptors, the concept can also be extended to the study of other receptors involved in the pathophysiology or therapy of psychiatric disorders.
Similar content being viewed by others

Introduction
Serotonin (5-hydroxytryptamine, 5-HT) receptors constitute a large family of seven receptor classes (5-HT1 to 5-HT7) with 14 different subtypes. Five subgroups are distinguished within the 5-HT1 receptor group, i.e., 5-HT1A, 5-HT1B, 5-HT1D, 5-HT1E and 5-HT1F receptors. The 5-HT1A subtype was the first of the serotonin receptors to be sequenced and is nowadays one of the most thoroughly studied. Its distribution within the brain is classically divided into two functional groups: presynaptic autoreceptors which are present at high densities in the midbrain raphe nuclei, and postsynaptic heteroreceptors in other brain regions, notably the hippocampus, the cingulate cortex, the lateral septum and other brain regions that are important for control of mood and cognition. Presynaptic 5-HT1A autoreceptors on serotonergic neurons mediate an important negative feedback influence on neuronal firing, thus eliciting inhibition of 5-HT release. Postsynaptic 5-HT1A receptors are located on different cell types, including GABAergic interneurons and excitatory pyramidal and granule cells. Due to its modulating influence on the whole serotonergic system and its large distribution throughout the brain, the 5-HT1A receptor is considered as one of the most important 5-HT receptor subtypes, and is of primary interest when investigating serotonergic neurotransmission in brain1,2.
The 5-HT1A receptor is strongly associated with alterations in mood and emotion, as demonstrated by numerous preclinical and clinical studies1. Many of these are focused on depression and anxiety, but compelling evidence also points to an important role of 5-HT1A receptor dysfunction in schizophrenia3. This chronic psychiatric disease affects around 20 million people worldwide4, creating a substantial global burden of care, with significant morbidity and functional impact4,5. Notably, life expectancy in patients with schizophrenia has been estimated to be 10–20 years shorter than in the general population6. Although schizophrenia is associated with an imbalance in dopamine transmission, with an hyperactivity in the mesolimbic system and an hypoactivity in the prefrontal cortex, other neurotransmitters, including glutamate, GABA and serotonin, are also involved in its physiopathological process7,8. The main symptoms of schizophrenia comprise delusions, hallucinations, disorganization and negative symptoms, i.e., blunted affect or aboulia. Diagnosis is based on the presence of at least two of these main symptoms for 6 months or more, leading to significant functional impairment. Cognitive impairment is a core feature of schizophrenia and may be present at any stage of the disease9. Both negative and cognitive symptoms are associated with poor functional outcomes10 and are currently not well-managed by available pharmacological treatments. A better understanding of the physiopathology of schizophrenia is thus required to identify and validate pharmacological targets and, ideally, lead to novel therapeutic strategies for patients.
The involvement of serotonin 5-HT1A receptors in the pharmacology of antipsychotics
Early classical (i.e., typical) antipsychotics (e.g., phenothiazine, butyrophenone, and benzamide derivatives) possess dopamine D2 receptor blocking activity and show efficacy for control of positive symptoms. These agents, however, are not very effective for treatment of negative or cognitive symptoms and frequently induce extra-pyramidal side effects. Subsequently, a variety of atypical antipsychotics became available as first-line treatments. On a pharmacological level, these atypical antipsychotics commonly act as antagonists on both the dopaminergic D2 receptor and the serotoninergic 5-HT2A receptor, as well as interacting with multiple other receptors11,12,13. For example, antipsychotics such as risperidone or olanzapine display the desired two-fold action (D2 and 5-HT2A) and exhibit efficacy against positive symptoms with improved activity on negative and cognitive symptoms and reduced side effects, such as extra-pyramidal symptoms (EPS)11,12,13. Nonetheless, there are still unmet needs in the treatment of schizophrenia, including patients who are resistant to treatment with current antipsychotics14,15, the need to more efficaciously improve cognitive deficits, alleviate affective disorders (e.g., anxiety and depression) and reduce antipsychotic‐induced EPS, metabolic and endocrine side effects16.
It is notable that some atypical antipsychotic drugs that primarily target D2 and 5-HT2A receptors, such as risperidone and olanzapine, also indirectly activate 5-HT1A receptors17. Other antipsychotics, including clozapine, ziprasidone and quetiapine, act directly as partial agonists at 5-HT1A receptors18. Moreover, ‘third generation’ antipsychotics, such as aripiprazole, cariprazine and brexpiprazole, exhibit prominent direct-acting agonist properties at 5-HT1A receptors19,20,21. Indeed, 5‐HT1A receptors are now an important therapeutic target for selection of novel antipsychotics based on a range of observations12,22. For example, 5-HT1A receptor activation has been proposed as a target for the development of cognitive enhancers for treatment of cognitive dysfunction in schizophrenia, notably via stimulation of dopamine release in key regions such as the frontal cortex11,23. Thus, some authors proposed that verbal memory and executive functioning were improved in schizophrenia patients that received the 5-HT1A partial agonist, tandospirone, in addition to neuroleptic treatment24,25. Based on molecular, neurochemical, behavioural and clinical data, it has been suggested that agonism of the 5-HT1A receptor may ameliorate the therapy of negative symptoms and comorbid mood deficits, attenuate cognitive deficits and oppose EPS elicited by D2 receptor blockade. Future antipsychotics may therefore be identified that possess an optimized dual D2/5-HT1A mechanism which retains efficacy against positive symptoms via D2 antagonism, whilst improving cognitive impairment and negative symptoms associated with schizophrenia and functional outcome by activation of 5-HT1A receptors26,27.
In addition to pharmacological factors such as those described above, genetic factors could also influence the role of 5-HT1A receptors in schizophrenia28. Indeed, accumulating evidence indicates that a common 5-HT1A gene promoter single-nucleotide polymorphism (SNP rs6295, C-1019G), which occurs in about 5–10% in Oriental populations and 40% in Caucasian population29, leads to changes in 5-HT1A receptor function that markedly affect vulnerability to psychiatric disorders, including schizophrenia30. Thus, several studies highlighted the correlation between 5-HT1A receptor polymorphisms and the presence of schizophrenia symptoms and their response to treatments31,32,33. This single-nucleotide polymorphism is proposed to have a relevant impact on 5-HT1A receptor density and/or activity. In raphe nuclei, the C-G change linked to the polymorphism impairs the binding of nuclear proteins (e.g., Deaf1) to a palindromic DNA sequence located at this polymorphism. As a result, the rs6295 G allele is associated with a reduced efficiency of Deaf-1 binding, expressed in an increase of 5-HT1A autoreceptor density in the raphe nucleus. In contrast, the 5-HT1A receptor expression is proposed to be decreased on the postsynaptic part, such as in the prefrontal cortex. This lowered postsynaptic 5-HT1A receptor density is linked to a reduced serotonergic signal transduction, which may in turn reduce dopamine release in the prefrontal cortex34. As a consequence, this serotonin imbalance may have implications regarding response to antipsychotics (notably as concerns negative symptoms) and treatment options for patients with schizophrenia29. Taken together, these findings reinforce the assertion that the 5-HT1A receptor is a relevant target for treatment of schizophrenia and other psychiatric disorders.
The exploration of 5-HT1A receptors in schizophrenia by molecular imaging and its contradictory results
The development of molecular imaging techniques with selective radioligands allows direct measurement of serotonin receptors in vitro on brain tissue by autoradiography, or in vivo, in the living human brain by positron emission tomography (PET). Several compounds have been radiolabelled for studies of the 5-HT1A receptor. The most commonly used radioligand for in vitro studies is [3H]-8-OH-DPAT and the most frequent PET radiopharmaceuticals for quantification of 5-HT1A receptors in the living human brain are [11C]-WAY100635 and [18F]-MPPF.
Post-mortem studies and their shortcomings
The first exploration of the 5-HT1A receptor densities in the brain of schizophrenic patients was carried out by in vitro autoradiography on post-mortem brain tissues. The aim was to investigate the role of the 5-HT1A receptor in the pathophysiology of schizophrenia. Other studies followed and, in total, there have been eleven post-mortem studies exploring the G-protein-coupled serotonin 5-HT1A receptor in patients with schizophrenia compared to healthy controls by homogenate or in situ autoradiographic binding techniques (for review, see ref. 35). Although the majority of these post-mortem studies described an increase of 5-HT1A receptor densities in various cortical regions, several articles reported contradictory results. Six studies showed an increase in 5-HT1A receptor density in the prefrontal cortex (PFC) of patients with schizophrenia compared with healthy controls36,37,38,39,40,41 whereas three did not42,43,44; one study found a significant increase in the temporal cortex38 and two did not36,44. Other contradictory results mainly concerned the anterior cingulate cortex and the hippocampus36,37,38,40,44,45. One study showed an increase in 5-HT1A receptor density in the cerebellum of patients with schizophrenia compared with controls46. No studies found any decrease of the 5-HT1A receptors in any brain region in patients with schizophrenia compared to controls. Overall, meta-analysis from Selvaraj and colleagues35 found that the pooled results were in favour of a significant increase in 5-HT1A receptors in the prefrontal cortex of patients with schizophrenia compared to healthy controls, with an effect size of 0.60 (CI: 0.17–1.03, p = 0.007)35. For detailed results, see Table 1.
PET studies and their shortcomings
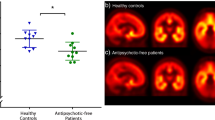
There have been few in vivo 5-HT1A positron emission tomography (PET) imaging studies in schizophrenia patients. PET imaging theoretically has great advantages over the in vitro approach. Indeed, in vivo imaging allows the exploration of live patients, allowing for possible longitudinal follow-up. Moreover, PET imaging permits exploration of the entire brain, potentially revealing regions of interest not examined in the brain tissue sections available in brain banks. However, results from PET studies on the 5-HT1A receptor in schizophrenia were mainly non-significant47,48 or contradictory49,50. For example, a PET study focused on 5-HT1A receptor availability in schizophrenia patients treated with ziprasidone and found no significant difference in their distribution before or after receiving medication but found a significant association between 5-HT1A binding and improvement in negative symptoms51. For detailed results, see Table 2.
It is striking that although molecular imaging studies of the 5-HT1A receptor in schizophrenia report disparate results, none of them have yet confirmed post-mortem findings. The contrast between in vitro and in vivo findings may be attributable to several factors, such as variations in the sample populations (age, duration of illness, pharmacological treatments…) but also to the methodological approaches. Thus, there are significant pharmacological differences between the radiotracers used for quantification of in vitro and in vivo studies. The 5-HT1A receptor can exist in a high (G-protein-coupled, functionally active) or low (G-protein-decoupled, non-functionally active) affinity state with respect to endogenous binding. As mentioned before, most post-mortem studies have used the agonist [3H]-8-OH-DPAT, which binds with high affinity only to receptors in high-affinity state and it also has limited specificity, binding to both 5-HT1A and 5-HT7 receptors52. On the contrary, PET studies used antagonist radiotracers, [11C]-WAY100635 or [18F]-MPPF, that are more specific (although WAY100635 also has nanomolar range binding affinity for dopamine D4 receptors, rarely taken into account53), and that bind with comparable affinity to 5-HT1A receptors in both high- and low-affinity states54. While these characteristics are well known to neuropharmacologists, they are seldom given consideration in the nuclear medicine and neuroscience communities.
The concept of functionally active 5-HT1A receptor imaging
Principle: antagonist versus agonist
The vast majority of PET radiopharmaceuticals currently used for receptor neuroimaging are antagonists. However, about 30 agonists have been used for PET imaging of various G-protein-coupled receptors: adrenergic, dopaminergic, serotonergic, muscarinic, cannabinoid, and opioid receptors55,56,57. Among them, a dozen of PET agonist radiotracers were developed and translated to humans for these receptors, but very few studies58,59,60 directly compared the in vivo density of receptors targeted by an agonist versus an antagonist radiotracer in the same subject (in these cases, animal models). Several interpretations have been given to in vivo binding differences based on the pharmacological agonist/antagonist properties of the ligands. The internalization of certain receptors has been evoked, as well as the modification of their allosteric conformations, or the influence of the extracellular concentration of the endogenous neurotransmitter, all of which could influence differently the binding of an antagonist and an agonist for the same family of receptors (see ref. 56 for a review on this subject).
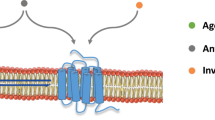
We have proposed a different interpretation, largely based on pharmacological facts and supported by experimental results. Thus it was shown several decades ago that 5-HT1A receptors can exist in a high or a low-affinity state, depending on whether they are coupled or not to their G proteins61,62. This property implies that whereas 5-HT1A receptor antagonists bind to the total pool of receptors, 5-HT1A receptor agonists preferentially bind to a subpopulation of receptors in their high-affinity state63. Consequently, the radiolabelled antagonists, i.e. the current PET radiopharmaceuticals, allow PET neuroimaging of the total pool of receptors, regardless of their functional state. In contrast, and according to this pharmacological mechanism, agonists bind preferentially to the functionally active state of receptors (see Fig. 1).

The antagonist radiotracer binds to all states of the receptors, i.e. functional (bound to G protein) and non-functional (uncoupled form) whereas the agonist only binds to functional receptors (bound to G protein). As a result, the binding pattern at 5-HT1A receptors differs significantly between the antagonist [18F]-MPPF (binding in hippocampus, cingulate cortex and raphe nuclei) and the agonist [18F]-F13640 (binding mainly in cortical and cerebellar regions).
[18F]-F13640, a new radiopharmaceutical validated in human subjects
In order to specifically target 5-HT1A receptors in their G-protein-coupled functionally active state, we have developed a highly selective 5-HT1A receptor agonist, [18F]-F1364064. F13640, also known as befiradol or NLX-112, is a potent ligand with nanomolar affinity at 5-HT1A receptors, and exceptional selectivity over a wide range of more than fifty receptor and transporter subtypes65,66. Its pharmacology has been extensively investigated, showing that it efficaciously activates 5-HT1A receptors both in vitro and in vivo. Its specificity for 5-HT1A receptors in vivo is supported by the observation that its effects in various experimental models are abolished by co-administration of the 5-HT1A receptor antagonist, WAY100635.
The use of [18F]-labelled F13640 as an agonist PET tracer for functionally active 5-HT1A receptors was recently investigated and validated in animal models (rat, cat and non-human primate). The results showed that the in vivo distribution pattern of [18F]-F13640 is very different from that of an antagonist radiotracer, e.g. [18F]-MPPF. There were also striking distribution differences for [18F]-F13640 between in vitro and in vivo results in rat brain. A probable explanation for the distinct binding patterns between 5-HT1A agonist and antagonist radiotracers could be that the ratio of G-protein-coupled versus uncoupled receptors might be much lower in vivo compared to in vitro findings. In several animal species, although the blockade of labelling by the antagonist WAY-100635 was incomplete, [18F]-F13640 binding was almost completely blocked by the agonist 8-OH-DPAT. This may be due to a more effective competition between agonists as they both compete for receptors in the high-affinity state only. Notably, in vitro [18F]-F13640 binding was markedly reduced by Gpp(NH)p, an agent that switches receptors into an uncoupled state, demonstrating that [18F]-F13640 binds preferentially to G-protein-coupled 5-HT1A receptors, a property that is consistent with its agonist activity64. In contrast, uncoupling of 5-HT1A receptors from G proteins tends to increase the binding of the antagonist [18F]-MPPF, as previously reported67,68. These results highlight the fact that although both [18F]-F13640 and [18F]-MPPF are selective 5-HT1A receptor radioligands, they bind to different pools of receptors depending on their coupling state. This proof of concept of the specific binding of [18F]-F13640 to functionally active receptors was supported by a post-mortem study in Alzheimer subjects68. This study showed that the in vitro binding of [18F]-F13640 differs from that of [18F]-MPPF, the prototypical radiopharmaceutical. [18F]-F13640 is therefore a promising PET tracer for in vivo imaging and quantification of functionally active 5-HT1A receptors in the human brain, and a first-in-man study with this radiopharmaceutical is currently underway (study registration: EudraCT 2017-002722-21). The first human in vivo brain images with [18F]-F13640 showed a binding pattern different from that seen with conventional antagonist 5-HT1A radiopharmaceuticals69. Although the binding regions were well correlated with the density of 5-HT1A receptors, the regions of interest were mainly cortical, indicating that these brain regions are enriched in functionally active receptors. The [18F]-F13640 binding in cerebellum is less frequent, since this region is known to be poor in 5-HT1A receptors, although PET studies in volunteers have revealed the presence of 5-HT1A receptors in the cerebellar vermis of the cerebellum70.
PET imaging of functionally active 5-HT1A receptors: how to revisit this receptor in schizophrenia?
The development of [18F]-F13640 as an agonist PET radiopharmaceutical opens new opportunities for nuclear medicine. Until now, in vivo exploration of 5-HT1A receptor in schizophrenia has mostly focused on the total pool of receptors with the default use of antagonist PET radiopharmaceuticals. We propose that the specific exploration of “functional” 5-HT1A receptors, coupled to their G proteins, opens the way to revisiting the in vivo exploration of this receptor. Comparison between agonist and antagonist PET radiotracer binding could provide a promising and unprecedented receptor mapping strategy, potentially revealing new pathophysiological mechanisms. The availability of a radiopharmaceutical able to measure 5-HT1A agonist sites, enables the undertaking of different protocols to explore 5-HT1A receptors in schizophrenia: either using [18F]-F13640 alone, or by comparing the labelling of [18F]-F13640 with that of [18F]-MPPF or [11C]-WAY1006365. By opening the possibility to specifically explore functionally active receptors instead of total receptor densities, use of [18F]-F13640 could help to investigate issues that have remained unresolved by previous neuroimaging protocols.
Revisiting pathophysiological processes
Protocols based on the concept of functionally active receptors may help to better define how variations in the expression of 5-HT1A receptors could represent a biological trait marker of schizophrenia. First, comparing subjects with schizophrenia with healthy controls matched for sex and age could bring greater precision in the mapping of receptor densities. This could help to clarify the contradictory results previously found in the limbic and frontal cortices in post-mortem and in vivo studies, notably because the pronounced cortical distribution of [18F]-F13640 binding is favourable to the exploration of these brain areas. As a next step, the identification of new functionally active receptor patterns could be associated to cognitive status and illness state. The question of the presence of depression in schizophrenia, considered by some authors as more than a comorbidity71 could also be explored, leading to more subtle phenotyping of different populations of schizophrenic patients. In particular, these patterns could also be linked to suicidal behaviour for which 5-HT1A receptor status is a key player72, life-time prevalence of suicide being around 5% in patients with schizophrenia73. Other notable clinical presentations such as catatonia that, according to the DSM-5, is not linked as a subtype to schizophrenia anymore, could be explored through the prism of the functionally active receptor pattern. As gender differences have been little studied so far or only provided preliminary results that deserved further investigation39,40,42, additional analysis could also be carried out concerning this question. Such protocols should be extended to schizophrenia patients’ relatives, since this disorder is defined by a high heritability and genetic vulnerability28. Finally, genetic polymorphisms of the 5-HT1A receptor, as described earlier, could be at the origin of variation in the density of functionally active receptors and in treatment response to antipsychotics. All these neuroimaging protocols and the extension of this functionally active receptor approach to other neurotransmitters, will undoubtedly strengthen the concept of Research Domain Criteria (RDoC) proposed as an alternative method of classification in psychiatry with more emphasis on neurobiology74.
Revisiting pharmacological mechanisms
The use of protocols based on the concept of functionally active receptor would also provide new insights into the understanding of pharmacological responses. In a previous PET study performed with the radiolabelled antagonist [11C]WAY100635, no difference in total 5-HT1A receptor densities was found in schizophrenia patients before and after treatment with the antipsychotic ziprasidone, but there was a significant association between 5-HT1A binding and improvement in negative symptoms51. Longitudinal studies based on the examination of functionally active receptor densities before and after treatment with antipsychotic drugs could revisit these previous results and show some changes in receptor patterns. Do drug-resistant subjects have a specific pattern of functionally active receptors? Do these receptors change differently over the course of treatment, depending on its efficacy? These protocols will be demanding because of the possible 5-HT1A receptor occupancy rate of some antipsychotics having affinity for this receptor. To avoid such potential confounding factors related to drug occupancy, it may therefore be advisable to initially focus investigation on the effects of antipsychotics having a weak affinity for 5-HT1A receptors. These results could then be correlated to the treatment response rate and may define new profiles between the responders and the non-responders. Moreover, there may be distinct patterns of functionally active receptor densities in drug-resistant patients.
Integrating new multimodal imaging approaches
The literature in biological psychiatry is rich in clinical and research studies that have been performed using different imaging modalities on both separate positron emission tomography (PET) and magnetic resonance (MR) scanners. However, the current emergence of PET/MR cameras could broaden this approach75. While neurologists already make extensive use of this hybrid imaging modality, there is little research currently published in psychiatry using PET/MR. Although PET is able to quantify receptor densities, receptor occupancy by antipsychotics, and, with the development of agonist PET tracers such as [18F]-F13640, also quantify receptors specifically coupled to their G proteins, this approach cannot measure dynamically functional information. On the contrary, functional MRI offers the opportunity to map changes in cerebral activity occurring at resting state or during cognitive tasks and after administration of drugs; but it lacks mechanistic specificity as it provides no information on the receptors primarily targeted76,77. Combination of regional patterns of 5-HT1A functionally active receptors, functional connectivity, and grey matter volume, derived from the disease characteristic networks, can potentially discriminate individual patients with different phenotypes. By analysing the functional connectivity of resting-state fMRI data in schizophrenia, the cortico–cerebellar–striatal–thalamic loop abnormalities, classically described could be correlated with the cortical binding of [18F]-F13640, which in turn could be associated with the cognitive performance of the subjects. Here again, the possibility of longitudinal follow-up, and thus intra-individual comparisons, would allow a distinction to be made between those subjects who respond and those who do not respond to pharmacological or psychotherapeutic strategies.
Conclusions
The exploration of 5-HT1A receptors in psychiatry has given rise to numerous studies. Although this receptor is associated with the pathophysiological mechanisms of schizophrenia and the pharmacological mechanisms of new-generation antipsychotics, its characterization remains incomplete. The contributions of molecular imaging, first in vitro on brain tissue, then in vivo, thanks to PET imaging, have not permitted significant advances but the recent concept of PET neuroimaging of G-protein-coupled receptors makes it possible to revisit PET brain exploration by enabling totally new research paradigms. These will make it possible to follow changes in the pattern of functionally active 5-HT1A receptors in relation to disease symptomatology, such as cognitive dysfunction or mood deficits, both to better distinguish phenotypes and also to understand the basis of the therapeutic response to antipsychotic drugs. It is likely that the strategy of targeting functionally active receptors, as described above for the 5-HT1A receptor, could also be usefully extended to other crucial receptors in pathophysiology or therapy of schizophrenia.
References
Popova, N. K. & Naumenko, V. S. 5-HT1A receptor as a key player in the brain 5-HT system. Rev. Neurosci. 24, 191–204 (2013).
Barnes, N. M. & Sharp, T. A review of central 5-HT receptors and their function. Neuropharmacology 38, 1083–1152 (1999).
Haleem, D. J. 5-HT1A receptor-dependent control of nigrostriatal dopamine neurotransmission in the pharmacotherapy of Parkinson’s disease and schizophrenia. Behav. Pharm. 26, 45–58 (2015).
James, S. L. et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet 392, 1789–1858 (2018).
Rössler, W., Joachim Salize, H., van Os, J. & Riecher-Rössler, A. Size of burden of schizophrenia and psychotic disorders. Eur. Neuropsychopharmacol. 15, 399–409 (2005).
Laursen, T. M., Nordentoft, M. & Mortensen, P. B. Excess early mortality in schizophrenia. Annu. Rev. Clin. Psychol. 10, 425–448 (2014).
Brisch, R. et al. The role of dopamine in schizophrenia from a neurobiological and evolutionary perspective: old fashioned, but still in vogue. Front. Psychiatry 5, 47 (2014).
Meltzer, H. Y., Li, Z., Kaneda, Y. & Ichikawa, J. Serotonin receptors: their key role in drugs to treat schizophrenia. Prog. Neuropsychopharmacol. Biol. Psychiatry 27, 1159–1172 (2003).
American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th edn. (Washington, DC, 2013).
Carbon, M. & Correll, C. U. Thinking and acting beyond the positive: the role of the cognitive and negative symptoms in schizophrenia. CNS Spectr. 19, 35–53 (2014).
McCreary, A. & Newman-Tancredi, A. Serotonin 5-HT1A receptors and antipsychotics—an update in light of new concepts and drugs. Curr. Pharm. Des. 21, 3725–3731 (2015).
Newman-Tancredi, A. The importance of 5-HT1A receptor agonism in antipsychotic drug action: rationale and perspectives. Curent Opin. Investig. Drugs 11, 2040–3429 (2010).
Stahl, S. M. & Shayegan, D. K. The psychopharmacology of ziprasidone: receptor-binding properties and real-word psychiatric practice. Clin. Psychiatry 64, 6–12 (2003).
McIlwain, M. E., Harrison, J., Wheeler, A. J. & Russell, B. R. Pharmacotherapy for treatment-resistant schizophrenia. Neuropsychiatr. Dis. Treat. 7, 135–149 (2011).
McNabb, C. B. et al. Functional network dysconnectivity as a biomarker of treatment resistance in schizophrenia. Schizophr. Res. 195, 160–167 (2018).
Howes, O. H. & Kaar, S. J. Antipsychotic drugs: challenges and future directions. World Psychiatry 17, 170–171 (2018).
Ichikawa, J. et al. 5-HT2A and D2 receptor blockade increases cortical DA release via 5-HT1A receptor activation: a possible mechanism of atypical antipsychotic-induced cortical dopamine release: atypical antipsychotics and 5-HT1A activation. J. Neurochem. 76, 1521–1531 (2001).
Newman-Tancredi, A. et al. Agonist and antagonist actions of antipsychotic agents at 5-HT1A receptors: a [35S]GTPgammaS binding study. Eur. J. Pharm. 355, 245–256 (1998).
Maeda, K. et al. Brexpiprazole I: in vitro and in vivo characterization of a novel serotonin-dopamine activity modulator. J. Pharm. Exp. Ther. 350, 589–604 (2014).
Stark, A. D. et al. Interaction of the novel antipsychotic aripiprazole with 5-HT1A and 5-HT2A receptors: functional receptor-binding and in vivo electrophysiological studies. Psychopharmacology 190, 373–382 (2007).
Kiss, B. et al. Cariprazine (RGH-188), a dopamine D3 receptor-preferring, D3/D2 dopamine receptor antagonist–partial agonist antipsychotic candidate: in vitro and neurochemical profile. J. Pharm. Exp. Ther. 333, 328–340 (2010).
Jones, C. A. & McCreary, A. C. Serotonergic approaches in the development of novel antipsychotics. Neuropharmacology 55, 1056–1065 (2008).
Meltzer, H. Y. & Sumiyoshi, T. Does stimulation of 5-HT1A receptors improve cognition in schizophrenia? Behav. Brain Res. 195, 98–102 (2008).
Sumiyoshi, T. et al. The effect of tandospirone, a serotonin(1A) agonist, on memory function in schizophrenia. Biol. Psychiatry 49, 861–868 (2001).
Sumiyoshi, T. et al. Enhancement of cognitive performance in schizophrenia by addition of tandospirone to neuroleptic treatment. Am. J. Psychiatry 158, 1722–1725 (2001).
Newman-Tancredi, A. & Kleven, M. S. Comparative pharmacology of antipsychotics possessing combined dopamine D2 and serotonin 5-HT1A receptor properties. Psychopharmacology 216, 451–473 (2011).
Ye, N., Song, Z. & Zhang, A. Dual ligands targeting dopamine D2 and serotonin 5-HT1A receptors as new antipsychotical or anti-Parkinsonian agents. Curr. Med. Chem. 21, 437–457 (2014).
Harrison, P. J. Recent genetic findings in schizophrenia and their therapeutic relevance. J. Psychopharmacol. 29, 85–96 (2015).
Newman-Tancredi, A. & Albert, P. R. In Schizophrenia Research: Recent Advances (ed. Sumiyoshi, T.) Ch. 15 (Nova Science Publishers, New York, 2012).
Huang, Y. Y. et al. Human 5-HT1A receptor C(−1019)G polymorphism and psychopathology. Int. J. Neuropsychopharmacol. 7, 441–451 (2004).
Reynolds, G. P., Arranz, B., Templeman, L. A., Fertuzinhos, S. & San, L. Effect of 5-HT1A receptor gene polymorphism on negative and depressive symptom response to antipsychotic treatment of drug-naive psychotic patients. Am. J. Psychiatry 163, 1826–1829 (2006).
Wang, L. et al. The 1019 C/G polymorphism of the 5-HT1A receptor gene is associated with negative symptom response to risperidone treatment in schizophrenia patients. J. Psychopharmacol. 22, 904–909 (2008).
Bosia, M. et al. Effect of 5-HT1A-receptor functional polymorphism on Theory of Mind performances in schizophrenia. Psychiatry Res. 188, 187–190 (2011).
Albert, P. R. Transcriptional regulation of the 5-HT1A receptor: implications for mental illness. Philos. Trans. R. Soc. Lond. B Biol. Sci. 367, 2402–2415 (2012).
Selvaraj, S., Arnone, D., Cappai, A. & Howes, O. Alterations in the serotonin system in schizophrenia: a systematic review and meta-analysis of postmortem and molecular imaging studies. Neurosci. Biobehav. Rev. 45, 233–245 (2014).
Burnet, P. W., Eastwoord, S. L. & Harrison, P. J. 5-HT1A and 5-HT2A Receptor mRNAs and binding site densities are differentially altered in schizophrenia. Neuropsychopharmacology 15, 442–455 (1996).
Burnet, P. W., Eastwood, S. L. & Harrison, P. J. [3H]WAY-100635 for 5-HT1A receptor autoradiography in human brain: a comparison with [3H]-8OH-DPAT and demonstration of increased binding in the frontal cortex in schizophrenia. Neurochem. Int. 30, 565–574 (1997).
Hashimoto, T., Nishino, N., Nakai, H. & Tanaka, C. Increase in serotonin 5-HT1A receptors in prefrontal and temporal cortices of brains from patients with chronic schizophrenia. Life Sci. 48, 355–363 (1991).
Simpson, M. D. C., Lubman, D. I., Slater, P. & Deakin, J. F. Autoradiography with [3H]8-OH-DPAT reveals increases in 5-HT(1A) receptors in ventral prefrontal cortex in schizophrenia. Biol. Psychiatry 39, 919–928 (1996).
Gurevich, E. V. & Joyce, J. N. Alterations in the cortical serotonergic system in schizophrenia: a postmortem study. Biol. Psychiatry 42, 529–545 (1997).
Sumiyoshi, T., Stockmeier, C. A., Overholser, J. C., Dilley, G. E. & Meltzer, H. Y. Serotonin1A receptors are increased in postmortem prefrontal cortex in schizophrenia. Brain Res. 708, 209–214 (1996).
Gray, L., Scarr, E. & Dean, B. Serotonin 1a receptor and associated G-protein activation in schizophrenia and bipolar disorder. Psychiatry 143, 111–120 (2006).
Dean, B., Tomaskovic-Crook, E., Opeskin, K., Keks, N. & Copolov, D. No change in the density of the serotonin1A receptor, the serotonin4 receptor or the serotonin transporter in the dorsolateral prefrontal cortex from subjects with schizophrenia. Neurochem. Int. 34, 109–115 (1999).
Joyce, J. N. et al. Serotonin uptake sites and serotonin receptors are altered in the limbic system of schizophrenics. Neuropsychopharmacology 8, 315–336 (1993).
Scarr, E., Pavey, G., Copolov, D. & Dean, B. Hippocampal 5-hydroxytryptamine receptors: abnormalities in postmortem brain from schizophrenic subjects. Schizophr. Res. 71, 383–392 (2004).
Slater, P., Doyle, C. A. & Deakin, J. F. Abnormal persistence of cerebellar serotonin-1A receptors in schizophrenia suggests failure to regress in neonates. J. Neural Transm. 105, 305–315 (1998).
Bantick, R. A. et al. A positron emission tomography study of the 5-HT1A receptor in schizophrenia and during clozapine treatment. J. Psychopharmacol. 18, 346–354 (2004).
Frankle, W. G. et al. Serotonin 1A receptor availability in patients with schizophrenia and schizo-affective disorder: a positron emission tomography imaging study with [11C]WAY 100635. Psychopharmacology 189, 155–164 (2006).
Tauscher, J. et al. Brain serotonin 5-HT1A receptor binding in schizophrenia measured by positron emission tomography and [11C]WAY-100635. Arch. Gen. Psychiatry 59, 514 (2002).
Yasuno, F. et al. Decreased 5-HT1A receptor binding in amygdala of schizophrenia. Biol. Psychiatry 55, 439–444 (2004).
Frankle, W. G. et al. Measurement of the serotonin 1A receptor availability in patients with schizophrenia during treatment with the antipsychotic medication ziprasidone. J. Psychopharmacol. 25, 734–743 (2011).
Hedlund, P. B. et al. 8-OH-DPAT acts on both 5-HT1A and 5-HT7 receptors to induce hypothermia in rodents. Eur. J. Pharm. 487, 125–132 (2004).
Chemel, B. R., Roth, B. L., Armbruster, B., Watts, V. J. & Nichols, D. E. WAY-100635 is a potent dopamine D4 receptor agonist. Psychopharmacology 188, 244–251 (2006).
Aznavour, N. et al. A comparison of in vivo and in vitro neuroimaging of 5-HT1A receptor binding sites in the cat brain. J. Chem. Neuroanat. 31, 226–232 (2006).
Zimmer, L. Pharmacological agonists for more-targeted CNS radio-pharmaceuticals. Oncotarget 7, 49 (2016).
Colom, M., Vidal, B. & Zimmer, L. Is there a Role for GPCR agonist radiotracers in PET neuroimaging? Front. Mol. Neurosci. 12, 255 (2019).
Shalgunov, V. et al. Hunting for the high‐affinity state of G‐protein‐coupled receptors with agonist tracers: theoretical and practical considerations for positron emission tomography imaging. Med. Res. Rev. 39, 1014–1052 (2019).
Narendran, R. et al. Measurement of the proportion of D2 receptors configured in state of high affinity for agonists in vivo: a positron emission tomography using [11C]N-propyl-norapomorphine and [11C]raclopride in baboons. J. Pharm. Exp. Ther. 315, 80–90 (2005).
Ginovart, N. et al. Binding characteristics and sensitivity to endogenous dopamine of [11C]-(+)-PHNO, a new agonist radiotracer for imaging the high-affinity state of D2 receptors in vivo using positron emission tomography. J. Neurochem. 97, 1089–1103 (2006).
Placzek, M. S. et al. Discrepancies in kappa opioid agonist binding revealed through PET imaging. ACS Chem. Neurosci. 10, 384–395 (2019).
Mongeau, R., Welner, S. A., Quirion, R. & Suranyi-Cadotte, B. E. Further evidence for differential affinity states of the serotonin1A receptor in rat hippocampus. Brain Res. 590, 229–238 (1992).
Emerit, M. B., el Mestikawy, S., Gozlan, H., Rouot, B. & Hamon, M. Physical evidence of the coupling of solubilized 5-HT1A binding sites with G regulatory proteins. Biochem. Pharm. 39, 7–18 (1990).
Gozlan, H., Thibault, S., Laporte, A. M., Lima, L. & Hamon, M. The selective 5HT1A antagonist radioligand [3H]WAY 100635 labels both G-protein-coupled and free 5HT1A receptors in rat brain membranes. Eur. J. Pharm. 288, 173–186 (1995).
Vidal, B. et al. 18F-F13640 preclinical evaluation in rodent, cat and primate as a 5-HT1A receptor agonist for PET neuroimaging. Brain Struct. Funct. 223, 2973–2988 (2018).
Newman-Tancredi, A. et al. Distinctive in vitro signal transduction profile of NLX-112, a potent and efficacious serotonin 5-HT1A receptor agonist. J. Pharm. Pharm. 69, 1178–1190 (2017).
Colpaert, F. C. et al. Large-amplitude 5-HT1A receptor activation: a new mechanism of profound, central analgesia. Neuropharmacology 43, 945–958 (2002).
Lemoine, L. et al. [18F]F15599, a novel 5-HT1A receptor agonist, as a radioligand for PET neuroimaging. Eur. J. Nucl. Med. Mol. Imaging 37, 594–605 (2010).
Vidal, B. et al. Agonist and antagonist bind differently to 5-HT1A receptors during Alzheimer’s disease: a post-mortem study with PET radiopharmaceuticals. Neuropharmacology 109, 88–95 (2016).
Colom, M. et al. 18F-F13640 PET imaging of functional receptors in humans. Eur. J. Nucl. Med. Mol. Imaging 47, 220–221 (2020).
Parsey, R. V. et al. Regional heterogeneity of 5-HT1A receptors in human cerebellum as assessed by positron emission tomography. J. Cereb. Blood Flow. Metab. 25, 785–793 (2005).
Upthegrove, R., Marwaha, S. & Birchwood, M. Depression and Schizophrenia: cause, consequence or trans-diagnostic issue? Schizophr. Bull. 43, 240–244 (2017).
Underwood, M. D. et al. Serotonin receptors and suicide, major depression, alcohol use disorder and reported early life adversity. Transl. Psychiatry 8, 279 (2018).
Hor, K. & Taylor, M. Suicide and schizophrenia: a systematic review of rates and risk factors. J. Psychopharmacol. 24, 81–90 (2010).
Cuthbert, B. N. The role of RDoC in future classification of mental disorders. Dialogues Clin. Neurosci. 22, 81–85 (2020).
Vidal, B. et al. In vivo biased agonism at 5-HT1A receptors: characterisation by simultaneous PET/MR imaging. Neuropsychopharmacology 43, 2310–2319 (2018).
Jenkins, B. G. Pharmacologic magnetic resonance imaging (phMRI): Imaging drug action in the brain. NeuroImage 62, 1072–1085 (2012).
Mwansisya, T. E. et al. Task and resting-state fMRI studies in first-episode schizophrenia: a systematic review. Schizophr. Res. 189, 9–18 (2017).
Acknowledgements
O.R. was awarded a scholarship from the French Congress of Psychiatry (Congrès Français de Psychiatrie). This work was supported by the LABEX PRIMES (ANR-11-LABX-0063) of Université de Lyon and the programme “Investissements d’Avenir” (ANR-11-IDEX-0007) operated by the French National Research Agency (ANR) and by the imaging platform, CERMEP (Lyon, France).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Razakarivony, O., Newman-Tancredi, A. & Zimmer, L. Towards in vivo imaging of functionally active 5-HT1A receptors in schizophrenia: concepts and challenges. Transl Psychiatry 11, 22 (2021). https://doi.org/10.1038/s41398-020-01119-3
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41398-020-01119-3
This article is cited by
-
[18F]F13640: a selective agonist PET radiopharmaceutical for imaging functional 5-HT1A receptors in humans
European Journal of Nuclear Medicine and Molecular Imaging (2023)
-
Trace Amine-Associated Receptor 1 as a Target for the Development of New Antipsychotics: Current Status of Research and Future Directions
CNS Drugs (2021)
