
Abstract
Background
The impact of degree of prematurity at birth on premature infant gut microbiota has not been extensively studied in comparison to term infants in large cohorts.
Methods
To determine the effect of gestational age at birth and postnatal exposures on gut bacterial colonization in infants, we analyzed 65 stool samples from 17 premature infants in the neonatal intensive care unit, as well as 13 samples from 13 mostly moderate-to-late premature infants and 189 samples from 176 term infants in the New Hampshire Birth Cohort Study. Gut colonization patterns were determined with 16S rDNA microbiome profiling.
Results
Gut bacterial alpha-diversity differed between premature and term infants at 6 weeks of age, after adjusting for exposures (p = 0.027). Alpha-diversity varied between extremely premature (<28 weeks gestation) and very premature infants (≥28 but <32 weeks, p = 0.011), as well as between extremely and moderate-to-late premature infants (≥32 and <37 weeks, p = 0.004). Newborn antibiotic use among premature infants was associated with lower Bifidobacterium and Bacteroides abundance (p = 0.015 and p = 0.041).
Conclusion
Gestational age at birth and early antibiotic exposure have significant effects on the premature infant gut microbiota.
Similar content being viewed by others


Introduction
Premature infants have an immature gastrointestinal tract, with a gut epithelium that has diminished barrier function and increased permeability. This allows for translocation of bacteria from the gut to the bloodstream, leading to systemic inflammation or sepsis.1,2,3,4,5,6 In premature infants it has been hypothesized that antibiotic use, coupled with the infants’ underdeveloped immune systems and gut epithelia, promotes intestinal bacterial communities that are less diverse and enriched with potential pathogens.7,8,9,10,11 Prematurity, lack of diversity in the gut bacterial communities, and the presence of pathogenic bacteria have also been associated with systemic inflammation and the development of sepsis, as well as necrotizing enterocolitis, all of which are important complications of prematurity.8,12,13 The bacteria that premature intestinal tracts lack include taxa frequently found to colonize the gut of healthy term infants, as well as bacteria that are associated with immune training, metabolic function, and ultimately decreased risk of lifelong health issues such as obesity and allergy.11,14,15,16
The factors responsible for the altered colonization patterns observed in premature infants compared to their healthy term counterparts are not well-defined. Physiological differences in the premature gut epithelium likely fosters abnormal colonization patterns from birth. Premature infants’ exposure to antibiotics before, during, and after birth, likely further contributes to aberrant gut bacterial colonization. Understanding how infant gut bacterial communities vary by gestational age at birth and how they are shaped by exposures can help inform future treatment in the hopes of decreasing pathogen colonization and increasing abundance of beneficial bacteria.
In this study, we aimed to evaluate how gut bacterial community structure varies across the spectrum of gestational age at birth and to assess the effects of various exposures, such as newborn antibiotic administration, delivery mode, and breastmilk feeding, on the gut microbiota of preterm infants.
METHODS
Ethical approval
Institutional review board approval was obtained from the Committee for the Protection of Human Subjects at Dartmouth College with yearly renewal. Parents provided written, informed consent.
Subject cohorts and inclusion criteria
We studied 17 premature infants hospitalized in a New Hampshire neonatal intensive care unit (NICU) and an additional 13 premature infants enrolled in the New Hampshire Birth Cohort Study (NHBCS) (Table 1). We compared these with 176 term infants also enrolled in the NHBCS. Inclusion criteria for the cohort of infants hospitalized in the NICU were appropriate size for gestational age (AGA) and ≤14 days of age at the time of enrollment. We excluded infants with major anomalies, such as trisomies, other genetic syndromes, and clinical syndromes. Major birth defects such as gastroschisis, complex congenital heart disease, or renal anomalies were excluded. Infants had a length of stay in the NICU between 29 and 174 days. Three infants in the NICU had sepsis, none had necrotizing enterocolitis, and five had bronchopulmonary dysplasia. A subset of these infants have been previously studied.8,17 The NHBCS is a prospective longitudinal large epidemiological cohort study of pregnant women and their children.18,19 Pregnant women aged 18–45 in New Hampshire are enrolled at ~24–28 weeks of gestation. Inclusion criteria for mothers include having a singleton pregnancy, a private drinking well at home, and no plans to move. We included infants with no noted congenital abnormality. Gut microbiome data on a large proportion of these infants has been previously described.20
Infant exposures
Data was collected on the following exposures: newborn antibiotic exposure (antibiotics given during the first days after birth while in hospital), delivery mode (vaginal delivery or Cesarean section), and breastmilk exposure (still consuming breastmilk at time of sample collection) (Table 2). Antibiotic administration data was abstracted from the infant’s hospital medical record. For infants hospitalized in the NICU, delivery and breastmilk feeding information was also abstracted from the medical record; for NHBCS subjects, these exposures were ascertained by telephone follow-up questionnaires with mothers. Additional exposure data was collected for the premature infants in the NICU, such as antibiotic exposure on day of sample collection and exclusive breastmilk feeding at time of sample collection (Table 3).
Sample collection
In the NICU cohort, stool samples were collected weekly beginning with first meconium and ending with hospital discharge. The samples were collected from diapers using sterile wooden depressors, transferred to sterile tubes, and aliquoted and stored at −80 °C. In the NHBCS, the stool samples were collected at two time points: at time of first stool after passing of meconium (‘transitional’ stool, 0–3 days of life) and at 6 weeks. Most NHBCS infants contributed samples at only one of these 2 time points. Premature infants from NHBCS born at <36 weeks gestation only contributed 6-week stool, so it was unlikely that they were hospitalized at time of sample collection. Stool was immediately frozen at home at −20 °C after collection. Then the stool samples were delivered in cold pack, thawed to 4 °C, aliquoted in sterile tubes, and suspended in RNA later. After 24 h in RNAlater in refrigerator, the samples were moved to −80 °C long term storage.
Sample processing
For the samples from the preterm infants in the NICU, DNA extraction was performed after thawing of samples using the MoBio Powersoil bacterial DNA isolation kit (MoBio, Carlsbad, California), as previously described.17 DNA was extracted from thawed NHBCS stool samples using Zymo Research’s ZR fecal DNA MiniPrep Kit, also described previously.20 Both kits include a bead-beating step. The MoBio kit used 0.7-mm garnet beads, while the Zymo kit used 0.5-mm high density ceramic beads.
Sample sequencing and data analysis
Sequencing of the V4–V5 hypervariable region of 16S ribosomal DNA was performed using the Illumina MiSeq platform. Details of sequencing methods, taxonomic assignment, and data analysis are described in Supplemental Methods.
RESULTS
Sequencing
After excluding 25 samples due to low read counts, 78 samples from 30 premature infants and 189 samples from 176 term infants were evaluated. The range of gestational ages for infants in each group is shown in Table 1. Sequencing of these samples resulted in a total of 22,012,483 reads for the 267 (mean: 82443.76, range: 14,002–1,072,107). The dominant bacteria identified in premature infant stool samples was Enterobacteriaceae, making up 32% of normalized reads, followed by the genera Clostridium (10%), Enterococcus (10%), and Veillonella (9%) (Fig. 1a). In term infants, bacterial genera from the family Enterobacteriaceae were also the most common, making up 20% of all normalized reads, followed by Bacteroides (19%), Bifidobacterium (15%), and Streptococcus (9%) (Fig. 1a). Bacteroides, Bifidobacterium, and Streptococcus made up only 7, 3, and 2% of total normalized reads among premature infants, respectively (Fig. 1a).

Top 10 most abundant taxa (a) and bacterial alpha-diversity differences between premature and term infants (b, c). a Bar charts depicting the percentage of normalized reads that a bacterial taxon makes up in the stool of premature infants and term infants. The top 10 most dominant taxa are depicted. b, c Bacterial alpha-diversity differences (b) over time and c across all time points. The bacterial alpha-diversity (SDI) of stool samples from premature and term infants is indicated on the y-axis. P-value is adjusted for exposures (newborn antibiotics, delivery mode, and breastmilk feeding at time of sample collection) and age (day of life). b Straight lines were fitted to the bacterial alpha-diversity data, and indicate the change in stool bacterial alpha-diversity over time (slope) in premature infants (red) and term infants (blue). The p-value represents the significance of the difference in slopes, and is adjusted for exposures. On the x-axis is the day of life at which a stool sample was collected. c Box-plots showing bacterial alpha-diversity differences between premature and term infants across all time points
Bacterial alpha-diversity and abundance differences between premature and term infants
Bacterial alpha-diversity, which was measured by Simpson’s Diversity Index (SDI), varied throughout the 6 weeks of life for individual premature infants with longitudinal stool samples (Fig. 1b). After adjusting for the effects of exposures, including delivery mode, newborn antibiotics, and breastmilk exposure at time of sample collection, there was no significant difference in the change in bacterial alpha-diversity between term and premature infants over time (p = 0.054, Fig. 1b). Across all time points, bacterial alpha-diversity was not statistically significantly different between term and premature infants after adjusting for exposures (p = 0.700, Fig. 1c).
Differences in bacterial abundance between premature and term infants were also assessed. After adjusting for exposures in addition to age at time of stool sample collection, there were no statistically significant differences in bacterial abundance between premature and term infants.
Hierarchical clustering
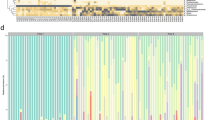
Hierarchical clustering analysis revealed that premature infant samples clustered separately from term infant samples, however, the clustering was not statistically significant (p = 1 for the cluster containing mostly premature infant samples, Fig. 2a). Moderate and late preterm infant samples (32 weeks to <37 weeks) interspersed between the extremely (<28 weeks) and very preterm (<32 weeks) and term samples (Fig. 2a).

Hierarchical clustering of stool samples of term and premature infants (a) and bacterial alpha-diversity differences at 6 weeks of age (b). a Below the dendrogram are bars depicting which samples belong generally to premature (black) or term infants (gray), as well as which samples belong to extremely preterm infants (green), very preterm (blue), or moderate-late preterm infants (red). b The bacterial alpha-diversity (SDI) of stool samples from premature and term infants is indicated on the y-axis. P-value is adjusted for exposures
Bacterial alpha-diversity and abundance differences at 6 weeks
At ~6 weeks of age, the difference in bacterial alpha-diversity, after accounting for exposures, was statistically significant (p = 0.027), with premature infants having lower bacterial alpha-diversity than term infants at 6 weeks (Fig. 2b). However, there were no statistically significant differences in bacterial abundance in premature infants compared to term infants at 6 weeks of age.
Bacterial phylogenetic relatedness differences between premature and term infants
Bacterial phylogenetic relatedness differences between premature and term infants were evaluated at the 6-week time point to address the more mature, established microbiome in both groups. After adjusting for exposures (but not repeated measures, see Methods), the difference in phylogenetic relatedness between the two groups of infants was not significant (p = 0.557, Fig. 3a).

Bacterial phylogenetic relatedness differences between term and premature infants at 6 weeks of age (a), and bacterial alpha-diversity differences based on gestational maturity (b) and bacterial abundance differences between infants born at an earlier or later gestational age (c). a Principal coordinates analysis plot showing bacterial phylogenetic relatedness differences between 6-week stool samples from premature (red) and term infants (blue). P-value is adjusted for exposures. PC1 principal coordinate 1, PC2 principal coordinate 2. Percentage refers to percentage of variance explained by the principal coordinate. b The bacterial alpha-diversity (SDI) of infant stool samples is indicated on the y-axis. P-value is adjusted for exposures and age (day of life). Infants are divided into groups based on gestational age at birth. Term is infants born at ≥37 weeks of gestation, moderate-late (Mod-Late) preterm infants are born at ≥32 weeks of gestation but <37 weeks, very preterm infants are born between 28 and <32 weeks of gestation, and extremely preterm infants are born between 24 and <28 weeks of gestation. Number of stool samples for each group is noted. However, exposure data for infants was occasionally not available, so number of samples evaluated for the adjusted analyses may be lower. c Bacterial taxa were evaluated for differences in abundance between infants born at a later compared to earlier gestational age at birth after with adjustment for exposures and age (day of life). On the x-axis, an abundance difference of greater than zero indicates that the bacterial taxon is found in greater abundance in infants born at a later gestational age (GA ≥ 32 weeks), while an abundance difference of less than zero indicates that the taxa is found in greater abundance in infants born at an earlier gestational age (GA < 32 weeks). On the y-axis is the -log10 of the p-value for the abundance difference. A greater -log10 of the p-value indicates a more significant p-value. Red circles indicate taxa whose abundance difference is significant (p-value is <0.05) after adjusting for multiple comparisons
Differences among premature infants by gestational age at birth
To test the hypothesis that gut bacterial community structure varied with degree of gestational immaturity at birth, we divided the premature infants into 3 groups: extremely premature infants (gestational age at birth <28 weeks), very premature infants (GA ≤28 to <32 weeks), and moderate-late premature infants (GA ≥32 weeks). We observed differences in bacterial alpha-diversity between extremely premature and moderate-late premature, and between extremely and very premature infants after adjusting for age and exposures (p = 0.004 and p = 0.011, respectively, Fig. 3b). We did not find differences in bacterial alpha-diversity between very and moderate-late premature infants after accounting for exposures and age (p = 0.697, Fig. 3b). There were differences in bacterial phylogenetic relatedness at 6 weeks of age between extremely and very premature infants (p = 0.033) after adjusting for exposures, but not between very and moderate-late premature infants (p = 0.138) or extremely and moderate-late premature infants (p = 0.160).
Earlier gestational age at birth compared to later gestational age at birth
Given the clustering of many moderate-late premature samples with term samples, we divided the infants into two groups: those who had an earlier gestational age at birth (GA < 32 weeks) and those who had a later gestational age (GA ≥ 32 weeks). Infants born at a later gestational age had greater abundance of Streptococcus and Bifidobacterium (p = 0.001, p = 0.001, respectively, Fig. 3c). At 6 weeks of age, after adjusting for exposures, infants born >32 weeks gestation had greater abundance of Bacteroides, while earlier GA infants <32 weeks had greater abundance of Parabacteroides (p = 0.001 and p = 0.005, respectively, Supplemental Figure S1). Across all time points, bacterial alpha-diversity was greater in later GA infants than in earlier GA infants, even after adjusting for exposures and age (day of life) (p = 0.015, Fig. 4a). Bacterial alpha-diversity did not, however, change differently over time between earlier and later GA infants (p = 0.298). Similarly, at 6 weeks of age, bacterial alpha-diversity, after accounting for exposures, remained distinct between earlier and later GA infants (p = 0.014). Bacterial phylogenetic relatedness differences between earlier vs. later GA infants at 6 weeks of age, after adjusting for exposures, were marginally statistically significant (p = 0.088, Fig. 4b).

Bacterial alpha-diversity (a) and phylogenetic relatedness differences (b) between infants with an earlier or later gestational age at birth. a Bacterial alpha-diversity differences in infants with an earlier gestational age at birth (<32 weeks GA, i.e., Earlier GA) and a later gestational age at birth (≥32 weeks, i.e., Later GA). P-value is adjusted for exposures and age (day of life). b Principal coordinates analysis plot showing bacterial phylogenetic relatedness differences between 6-week stool samples from infants with an earlier gestational age at birth (<32 weeks GA, shown in red) and a later gestational age at birth (≥32 weeks, shown in blue). P-value is adjusted for exposures. PC1 principal coordinate 1, PC2 principal coordinate 2. Percentage refers to percentage of variance explained by the principal coordinate
Additionally, when comparing moderate-late premature infants to term infants, there were no apparent differences in bacterial alpha-diversity after adjusting for exposures and age (p = 0.764), though the number of moderate-late premature compared to term samples was small (n = 16 compared to n = 174 after omitting samples missing exposure data). Bacterial phylogenetic relatedness differences at 6 weeks were not evident between moderate-late and term infants after accounting for exposures (p = 0.354). The abundance of Pantoea was statistically significantly greater in moderate-late premature infants compared to term infants after adjusting for exposures and age (p = 0.034), but there were no other statistically significant differences in bacterial abundance between these two groups of infants. These results indicate that moderate-late premature infants most likely have a community structure that more closely resembles term infants.
Analysis of corrected gestational age in premature infants
Corrected gestational age was defined at gestational age at birth + day of life at time of sample collection. We compared bacterial composition in premature infants born at different gestational ages when they were at similar corrected gestational ages at time of sample collection (Supplemental Figure S2). Although composition appears roughly different between infants born at different gestational ages, there were no significant differences in bacterial alpha-diversity between these groups of infants after accounting for exposures (Supplemental Table 1). Bacterial alpha-diversity does appear to increase as corrected gestational age increases (p = 0.003, Fig. 5).

Bacterial alpha-diversity in premature infants across corrected gestational age at time of sample collection. The bacterial alpha-diversity (SDI) of stool samples from premature indicated on the y-axis. A straight line was fitted to the bacterial alpha-diversity data, and indicates the change in stool bacterial alpha-diversity over corrected gestational age (slope) in premature infants. The p-value is adjusted for exposures, and represents the independent effect of corrected gestational age on bacterial alpha-diversity. On the x-axis is the corrected gestational age (gestational age at birth + day of life) at which a stool sample was collected
Associations with exposures among premature infants
We identified three potential exposures thought to affect premature infant gut microbial structure, which included newborn antibiotic exposure, delivery mode, and breastmilk exposure (Table 2). Additionally, we collected additional exposure data from the premature infants in the NICU, such as antibiotic exposure on day of sample collection and exclusive breastmilk feeding at time of sample collection (Table 3). Corrected gestational age at time of stool sample collection was also adjusted for. Among the premature infants, none of the three exposures (antibiotics in the first few days of life, a vaginal vs. C-section delivery, or breastmilk exposure) had statistically significant independent effects on bacterial alpha-diversity (p = 0.311, p = 0.324, p = 0.660) after adjustment for corrected gestational age. Earlier gestational age at birth negatively associated with bacterial alpha-diversity, but it was not significant after adjusting for exposures and corrected gestational age (p = 0.390). We also evaluated the effects of exposures and corrected gestational age on the abundance of the bacterial genera known to be abundant in term infants, such as Streptococcus, Bifidobacterium, Bacteroides, and Lactobacillus. Newborn antibiotic use was associated with lower Bifidobacterium and Bacteroides abundance (p = 0.015 and p = 0.041). Vaginal delivery was associated with greater Bacteroides abundance (p = 0.007), while breastmilk exposure at time of sample collection was associated with a decreased abundance of Lactobacillus (p = 0.017). Corrected gestational age at time of sample collection was positively associated with greater Lactobacillus and Streptococcus abundance (p = 0.031 and p < 0.001, respectively).
In the premature infant cohort in the NICU, after adjusting for corrected gestational age, no exposure was found to have significant independent effects on bacterial alpha-diversity, though corrected gestational age at time of sample collection had a positive independent effect (p = 0.015). Consumption of breastmilk at time of sample collection had marginally significant effects on bacterial phylogenetic relatedness at 6 weeks after adjusting for other exposures and corrected gestational age (p = 0.054). Antibiotics and corrected gestational age, but not delivery mode, exclusive breastmilk feeding or any breastmilk consumption at time of sample collection, had significant effects on bacterial abundance of Streptococcus, Bacteroides, and Lactobacillus. Antibiotic use at time of sample collection was associated with greater Bacteroides and Lactobacillus abundance (p = 0.047 and p = 0.042). Corrected gestational age at time of sample collection was positively associated with greater Lactobacillus and Streptococcus abundance (p = 0.017 and p = 0.021, respectively).
DISCUSSION
In our study, we identified differences in gut bacterial composition between premature and term infants in the first 6 weeks of life, using data from a cohort of premature infants in the NICU, as well as premature and term infants enrolled in a large, comprehensive molecular epidemiology study, the NHBCS. We found that dominant taxa differ between premature and term infants; however, there are no statistically significant differences in bacterial alpha-diversity, abundance, and phylogenetic relatedness after adjustment for the effects of exposures, such as newborn antibiotic administration, delivery mode, and breastmilk exposure at time of sample collection. We also evaluated the effects of various exposures on the premature infant intestinal microbiota, but also did not find any significant effects on bacterial alpha-diversity or phylogenetic relatedness. We did find significant differences in gut bacterial alpha-diversity between premature infants depending on gestational age at birth, as well as between premature and term infants at 6 weeks of age.
The primary difference between premature and term infants with respect to dominant taxa was the decreased abundance of the genera Bifidobacterium, Bacteroides, and Streptococcus. We also observed decreased abundances of Bifidobacterium, Bacteroides, and Streptococcus in the earlier GA at birth infants compared to those born at a later GA. A paucity of Bacteroides, Bifidobacterium, Streptococcus, and Lactobacillus in premature infants have been described previously.14,16 Bifidobacterium, some species of which may help prevent colonization of the infant gut by pathogenic organisms,21 is not well established in the gut of premature infants born at a gestational age earlier than 33 weeks.15 In other studies of term, vaginally delivered infants, there is high abundance of Bacteroides and Bifidobacterium less than a week after birth.22 Bifidobacterium and Bacteroides are bacterial genera considered health-promoting and interact directly with the burgeoning immune system in infancy.21,23,24 Bacteroides fragilis has been found to produce a bacterial polysaccharide that aids in the maturation of the immune system and production of regulatory T cells,23,24,25 and in a murine model noted for gut barrier defects, oral treatment with B. fragilis reduced gut permeability and altered microbial composition.26 We did find that early antibiotic use was associated with lower Bifidobacterium and Bacteroides abundance in premature infants, suggesting that early antibiotic exposure can affect future gut bacterial composition among premature infants. Streptococcus is also known to be one of the first colonizers of the gut in term infants,27,28 and in our study, we found that it was not among the top 10 dominant taxa in premature infants, while it was among the top 4 dominant tax in our term infants. Cesarean-section delivery has been associated with depletion of Lactobacillus;29 however, we did not find that delivery mode was associated with Lactobacillus abundance in the premature infants in our study.
We found that after performing hierarchical clustering, extremely and very preterm infants clustered together (GA < 32 weeks), but moderate-late preterm (GA ≤32 to <37) infant samples were interspersed among both the extremely and very preterm samples and the term samples. Furthermore, we found significant differences in the premature infant gut microbiota based on gestational age at birth. Extremely premature infants (born before 28 weeks of gestational age) had significantly lower bacterial alpha-diversity compared to very premature (GA ≥ 28 but <32 weeks) and moderate-late premature infants (GA ≥ 32 but < 37 weeks) even after adjusting for exposures. There were also significant differences in bacterial phylogenetic relatedness between the extremely and very premature infants. These results suggest that gestational age at birth has important associations with gut bacterial structure in premature infants, whether it be related to the maturity of the intestinal tract and the bacteria that are able to colonize this intestinal environment, or potentially related to medical treatment and length of stay in a hospital environment.
We compared bacterial alpha-diversity and phylogenetic relatedness between the moderate-late preterm infants and term infants, and found that they did not differ even after accounting for exposures, supporting the finding of moderate-late preterm and term samples clustering together. Comparing the extremely and very preterm infants (GA < 32 weeks) to the moderate-late preterm and term infants (GA ≥ 32 weeks), differences in bacterial alpha-diversity were identified. In an analysis of premature infants only, earlier gestational age was negatively associated with bacterial alpha-diversity, suggesting that greater gestational immaturity at birth is related to a less diverse gut microbiome. These results all together appear to suggest that infants born between 32 and 37 weeks of gestation have gut bacterial communities in the first 6 weeks of life are more similar to those of term infants than those of infants born before 32 weeks of gestation. This may point to the potential benefit of probiotic supplementation in extremely and very preterm infants, as their gut bacterial communities differ significantly from those of infants with a later gestational age at birth (≥32 weeks). Additionally, these differences in gut bacterial composition between infants born at an earlier vs. a later gestational age may reflect differences in environmental exposures between these two groups of infants, as more premature infants were hospitalized, exposed to more antibiotics and other medical interventions, while moderate-late preterm and term infants likely hospitalized for much shorter periods or not hospitalized at all, and were much less likely to have experienced medical interventions.
We did observe an upward trend in bacterial alpha-diversity with increasing corrected gestational age in the premature infants. Though we did not observe significant similarities in bacterial composition among premature infants of different degrees of gestational prematurity at birth when they were at similar corrected gestational ages, there were no significant differences in bacterial alpha-diversity between groups of infants. Corrected gestational age had a significant positive independent effect on bacterial alpha-diversity in premature infants, as well as a positive effect on the abundance of the bacterial genera Lactobacillus and Streptococcus.
Limitations of this study include a relatively small number of premature infants enrolled compared to the number of term infants, and a much smaller number of longitudinal samples among the term infants compared to premature infants. The study subjects were drawn from a relatively homogeneous northern New England population. Although this compromises the generalizability of our findings, it also reduces the sources of variation that could affect our observations. Also, two different DNA extraction kits were used, each for a different cohort, potentially confounding results comparing premature and term infants, and earlier GA at birth vs. later GA infants. Few studies have been published comparing the MoBio and Zymo DNA extraction methods, but one study suggests that representation of bacterial genera in samples does not significantly differ across these DNA extraction methods, though this study looked at plaque and saliva samples.30 A research report comparing our two kits showed that significantly different microbiomes were not produced if tested from the same intestinal sample, suggesting that the two kits give comparable results.31
Our analysis of the effects of diet on the premature gut microbiome was limited since all of our premature infants had exposure to breastmilk at some point. However, recent research involving the NHBCS cohort suggests that infants fed both breastmilk and formula more closely resemble infants that are exclusively formula fed than infants that are exclusively breastmilk fed.20 Thus, while most of our premature infants were consuming breastmilk at time of stool sample collection, they were also often receiving human milk fortifier or formula. We were, however, able to identify a number of samples where the premature infants had only been exposed to breastmilk. Additionally, our premature infants in the NICU were often fed donor breastmilk when maternal breastmilk was not available. Donor breastmilk is pasteurized, which may have affected gut colonization patterns as breastmilk contains commensal bacteria.32 Finally, in the bacterial phylogenetic relatedness analyses, we had to assume that each sample was independent, which may have biased results since not all infants donated the same number of samples.
CONCLUSION
In our study of premature and term infants, we found that premature infants had lower abundance of a number of keystone bacterial genera found in healthy term infants, many of which are considered immune training and health-promoting. We found an association between early antibiotic use and lower Bifidobacterium and Bacteroides abundance in our premature infants, pointing to the effect of early antibiotic exposure on future gut bacterial composition. We also found significant differences in bacterial alpha-diversity between premature infants based on gestational age at birth, with lower bacterial alpha-diversity seen in infants with an earlier gestational age at birth. When we compared infants born at an earlier gestational age (GA < 32 weeks) compared to infants born at a later gestational age (GA ≥ 32 weeks), we found differences in bacterial alpha-diversity (both at 6 weeks of age and across all time points). Bacterial alpha-diversity trended up with increased corrected gestational age. Our results suggest that premature infants with a later gestational age at birth resemble term infants, and that interventions to alter the gut microbiota may be of greatest value in extremely and very preterm infants.
References
Duffy, L. C. Interactions mediating bacterial translocation in the immature intestine. J. Nutr. 130, 432S–436S (2000).
Sherman, M.P. New concepts of microbial translocation in the neonatal intestine: mechanisms and prevention. Clin. Perinatol. 37, 565–579 (2010).
Carl, Ma et al. Sepsis from the gut: the enteric habitat of bacteria that cause late-onset neonatal bloodstream infections. Clin. Infect. Dis. 58, 1211–1218 (2014).
Tarr, P.I. & Warner, B.B. Gut bacteria and late-onset neonatal bloodstream infections in preterm infants. Semin. Fetal Neonatal Med. 21(6), 388–393 (2016).
Halpern, M. D. The role of intestinal epithelial barrier function in the development of NEC. Tissue Barriers 3, e1000707 (2015).
Mshvildadze, M. & Neu, J. The infant intestinal microbiome: friend or foe? Early Hum. Dev. 86 (Suppl 1), 67–71 (2010).
Greenwood, C. et al. Early empiric antibiotic use in preterm infants is associated with lower bacterial diversity and higher relative abundance of Enterobacter. J. Pediatr. 165, 23–29 (2014).
Madan, J. C. et al. Gut microbial colonisation in premature neonates predicts neonatal sepsis. Arch. Dis. Child Fetal Neonatal Ed. 97, F456–F462 (2012).
Arboleya, S. et al. Impact of prematurity and perinatal antibiotics on the developing intestinal microbiota: a functional inference study. Int J. Mol. Sci. 17, 649 (2016).
Gibson, M. K. et al. Developmental dynamics of the preterm infant gut microbiota and antibiotic resistome. Nat. Microbiol. 1, 16024 (2016).
Jacquot, A. et al. Dynamics and clinical evolution of bacterial gut microflora in extremely premature patients. J. Pediatr. 158, 390–396 (2011).
Mai, V. et al. Distortions in development of intestinal microbiota associated with late onset sepsis in preterm infants. PLoS One 8, e52876 (2013).
Zhou, Y. et al. Longitudinal analysis of the premature infant intestinal microbiome prior to necrotizing enterocolitis: a case-control study. PLoS One 10, 1–16 (2015).
Arboleya, S. et al. Establishment and development of intestinal microbiota in preterm neonates. FEMS Microbiol. Ecol. 79, 763–772 (2012).
Butel, M.-J. et al. Conditions of bifidobacterial colonization in preterm infants: a prospective analysis. J. Pediatr. Gastroenterol. Nutr. 44, 577–582 (2007).
Westerbeek, E. A. et al. The intestinal bacterial colonisation in preterm infants: a review of the literature. Clin. Nutr. 25, 361–368 (2006).
Chernikova, D. A. et al. Fetal exposures and perinatal influences on the stool microbiota of premature infants. J. Matern. Fetal Neonatal Med. 29, 99–105 (2016).
Gilbert-Diamond, D. et al. Rice consumption contributes to arsenic exposure in US women. Proc. Natl. Acad. Sci. USA 108, 20656–20660 (2011).
Farzan, S. F. et al. In utero arsenic exposure and infant infection in a United States cohort: a prospective study. Environ. Res. 126, 24–30 (2013).
Madan, J. C. et al. Association of cesarean delivery and formula supplementation with the intestinal microbiome of 6-week-old infants. JAMA Pediatr. 170, 1–8 (2016).
Butel, M. J. et al. Clostridial pathogenicity in experimental necrotising enterocolitis in gnotobiotic quails and protective role of bifidobacteria. J. Med. Microbiol. 47, 391–399 (1998).
Backhed, F. et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 17, 690–703 (2015).
Round, J. L. & Mazmanian, S. K. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc. Natl. Acad. Sci. USA 107, 12204–12209 (2010).
Mazmanian, S. K., Liu, C. H., Tzianabos, A. O. & Kasper, D. L. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 122, 107–118 (2005).
Telesford, K. M. et al. A commensal symbiotic factor derived from Bacteroides fragilis promotes human CD39(+)Foxp3(+) T cells and Treg function. Gut Microbes 6, 234–242 (2015).
Hsiao, E. Y. et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 155, 1451–1463 (2013).
Stark, P. L. & Lee, A. The microbial ecology of the large bowel of breast-fed and formula-fed infants during the first year of life. J. Med. Microbiol. 15, 189–203 (1982).
Songjinda, P. et al. Molecular monitoring of the developmental bacterial community in the gastrointestinal tract of Japanese infants. Biosci. Biotechnol. Biochem. 69, 638–641 (2005).
Dominguez-Bello, M. G. et al. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 107, 11971–11975 (2010).
Vesty, A., Biswas, K., Taylor, M. W., Gear, K. & Douglas, R. G. Evaluating the impact of DNA extraction method on the representation of human oral bacterial and fungal communities. PLoS One 12, 1–13 (2017).
Olsen K., Chaston J. Effects of Host Microbiota on Intestinal Fiber Content [Internet]. Brigham Young Univ. (2016) Available from: http://jur.byu.edu/?p=20043
Jost, T., Lacroix, C., Braegger, C. & Chassard, C. Assessment of bacterial diversity in breast milk using culture-dependent and culture-independent approaches. Br. J. Nutr. 110, 1253–1262 (2013).
Acknowledgements
The authors would like to acknowledge the children and families who participated in our studies, for which we are very grateful. We would also like to acknowledge the nurses and staff involved in the collection of samples and data. Additionally, we’d like to thank Dr. Elizabeth Brickley for her help and suggestions. NIH grants NIEHS P01 ES022832, US EPA RD83544201, NIGMS P20 GM104416, K01LM011985, R01AI59694, GM103534, GM103506; P20RR16448, 4UH3DK083993; K24AT003683, 5T32DK007301-35, AI116794, R01LM010098, 1P20ES018175-02 and RD-83459901-0, the Hearst Foundation, the Joshua Burnett Fellowship (the Hitchcock Foundation), Neukom Institute, and March of Dimes Ohio Prematurity Research Center Ohio Collaborative.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests or disclosures.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chernikova, D.A., Madan, J.C., Housman, M.L. et al. The premature infant gut microbiome during the first 6 weeks of life differs based on gestational maturity at birth. Pediatr Res 84, 71–79 (2018). https://doi.org/10.1038/s41390-018-0022-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41390-018-0022-z
This article is cited by
-
The gut microbiome, resistome, and mycobiome in preterm newborn infants and mouse pups: lack of lasting effects by antimicrobial therapy or probiotic prophylaxis
Gut Pathogens (2024)
-
CD71 + erythroid cells promote intestinal symbiotic microbial communities in pregnancy and neonatal period
Microbiome (2024)
-
Bifidobacterium regulates premature infant gut metabolites, reducing serum inflammatory factors: a randomised controlled trial
Pediatric Research (2024)
-
Age influences the temporal dynamics of microbiome and antimicrobial resistance genes among fecal bacteria in a cohort of production pigs
Animal Microbiome (2023)
-
Development of the gut microbiota during early life in premature and term infants
Gut Pathogens (2023)
