
Abstract
A3 adenosine receptor (A3AR) is a cell membrane protein, which has been found to be overexpressed in a large number of cancer types. This receptor plays an important role in cancer by interacting with adenosine. Specifically, A3AR has a dual nature in different pathophysiological conditions, as it is expressed according to tissue type and stimulated by an adenosine dose-dependent manner. A3AR activation leads to tumor growth, cell proliferation and survival in some cases, while triggering cytostatic and apoptotic pathways in others. This review aims to describe the most relevant aspects of A3AR activation and its ligands whereas it summarizes A3AR activities in cancer. Progress in the field of A3AR modulators, with a potential therapeutic role in cancer treatment are reported, as well.
Similar content being viewed by others
Adenosine and adenosine receptors
Biology of adenosine
Nucleosides and nucleotides are ubiquitous molecules with several biological functions, which constitute a number of molecule types that are essential for biological processes, including nucleic acids, co-enzymes, energy intermediates and intra-/extra-cellular messengers. The role of both nucleosides and nucleotides, as extracellular messengers, is relevant in some mechanisms, such as cell growth, migration, differentiation, bacterial-induced inflammation and growth factor secretion [1,2,3,4]. Adenosine is found at low concentrations (nanomolar range) in physiological conditions, while in stress conditions its concentration increases (micromolar range) [3]. In addition, physiological concentration of circulating adenosine can also vary in different species [5, 6]. In the extra-cellular environment, adenosine partly derives either from ATP, ADP and AMP hydrolysis by specific ectonucleotidases called ectonucleoside triphosphate diphosphohydrolase, or cluster of differentiation 39 (CD39) and ecto-5′-nucleotidase, or cluster of differentiation 73 (CD73) [4, 7]. Intra-cellular adenosine derives from AMP and S-adenosylhomocysteine (SAH) hydrolysis by endo-5′-nucleotidase and SAH hydrolase, respectively. This molecule can either be converted into AMP by adenosine kinase or be deaminated into inosine by the adenosine deaminase enzyme (ADA1 and ADA2) [4]. Adenosine can also be generated through a de novo biosynthesis pathway [8]. In particular, adenosine biosynthesis begins with the generation of ribosyl hypoxanthine monophosphate (IMP), which represents the first purine nucleotide that is synthesized de novo. IMP is initially converted to adenylo-succinate by adenylosuccinate synthase enzyme, while the latter is converted to AMP by adenylosuccinate lyase enzyme [8]. The intra-/extra-cellular concentration of adenosine is mediated by cell membrane complexes, named equilibrative (ENT1–4) and concentrative (CNT1–3) nucleoside transporters [9]. ENTs allow the passive transport of adenosine based on concentration differences, while CNTs use the gradient generated by sodium ions as an energy source to ensure adenosine transport against the concentration gradient [9]. Under physiological conditions, adenosine is transported from the extracellular to the intracellular environment, while, in hypoxic conditions, ENT1 downregulation blocks this flow, thereby leading to an increase in extracellular adenosine [9, 10]. Adenosine is abundant in the tumor microenvironment (TME), which includes tumor-surrounding blood vessels, immune cells, fibroblasts, signalling molecules and the extracellular matrix [11,12,13]. Adenosine also plays a role in tumor progression, as it is secreted by tumor/immune system cells in TME during this phase [3]. Both tumor promoting and antitumor properties have been reported for adenosine, while high adenosine levels have been reported in TME, as a consequence of hypoxia, which is a typical condition of solid tumors. This increase leads to a pro-angiogenic effect thereby leading to tumor development [3]. Contrariwise, a pro-apoptotic effect for this nucleoside has also been described in leukemia and melanoma in vitro models [3]. The dual role of adenosine seems to also depend on its concentration. Low concentrations (<25 nM) of this nucleoside inhibit tumor growth [14], while high concentrations (100 nM), similar to those determined in TME, confer to adenosine a pro-angiogenic effect [3].
Adenosine receptors
Adenosine performs its function by binding four different G-protein-coupled adenosine receptors (AR), i.e., A1, A2A, A2B and A3 (Fig. 1). The interaction between adenosine and its receptors not only activates pathways involved in different pathological processes, but also promotes receptor expression, in an autocrine manner. ARs comprise a group of glycoproteins with seven transmembrane domains and are coupled to G proteins (Fig. 1) [15]. Adenosine binding to A1R/A3R causes a decrease in cAMP, while adenosine binding to A2AR and A2BR causes an increase in cAMP. Evidence indicates that ARs expression appears to be mediated by DNA methylation [16], which is a critical epigenetic process for gene expression regulation [17, 18] in a variety of cell types [19,20,21,22,23].

In normal physiological condition adenosine mediates its activation via four G-protein coupled receptors A1, A2A, A2B, and A3. They are primarily associated with activation and inhibition of Adenylate Cyclase. The accumulation of cAMP is linked to the modulation of ion-channel activity.
A number of studies have reported that A3 receptor (A3AR) is overexpressed in cancers [24]. However, the role of A3AR in regulating cell proliferation and death is a relatively well debated issue, as this receptor acts differently depending on the tissue type in which it is expressed (Fig. 2) [25, 26]. In vitro models indicate that low (nanomolar) selective synthetic A3AR agonists concentrations protect normal cells from death, while A3AR agonists present pro-apoptotic effects in both normal and tumor cells at high (micromolar) concentrations. Notably, this dual A3AR-ligand mode of action comprises the same signaling pathway following A3AR activation (Fig. 2). Based on this evidence, A3AR is gaining interest for its potential use as a therapeutic antitumor target [15, 27]. It is thus clear that adenosine and A3AR, play a fundamental role in cancer.The aim of this review is to discuss the characteristics of A3AR and its cancer-related activities. Progress in the field of A3AR modulators, with a potential therapeutic role in cancer treatment, will be discussed, too.

The pro-tumoral activity of A3AR includes the increase of VEGF, HIF-1, MMP-9, angiogenesis, migration, proliferation, and invasion in tumor cells. The A3AR antitumoral activity leads to the decrease of cell proliferation and migration in several tumor cells and the induction of G-CSF and IL-2 secretion from cells of the immune system.
A3AR characteristics
A3AR genetic and protein characteristics and tissue distribution
The human A3AR coding gene maps on chromosome 1p21-p13. This gene contains two exons separated by a 2.2 kb intron, while encoding a protein of 318 amino acids. The gene regulatory region contains a cis sequence, which binds to several transcription factors including NF-κB [28]. The A3AR gene promoter upstream region presents a CCAAT consensus sequence and consensus binding sites for SP1, NF-IL6, GATA1, and GATA3 transcription factors [4]. Of these transcription factors, GATA3 is involved in the A3AR-dependent role in immune function [4]. A3AR protein presents seven alpha-helices containing about 20–27 amino acids. Each helix crosses the cell membrane seven times, while it is connected to an adjacent helix through three intracellular loops and three extracellular loops [29]. The amino-terminal region (N-terminus) is located outside the cell, while the carboxyl-terminal region (C-terminus) is oriented toward the cell cytoplasm. The presence of several tyrosine and serine residues at the C-terminus, confers a desensitization potential on A3AR during agonist administration [30]. The phosphorylation of this region leads to a decrease in agonist affinity and an increase in the ability of the agonist to inhibit adenylate cyclase activity, which catalyzes ATP conversion to cAMP [30].
A3AR is expressed in enteric neurons, epithelial cells, colon mucosa and lung parenchyma cells, chondrocytes, osteoblasts and also in cells responsible for inflammatory processes, such as mast cells, eosinophils, neutrophils, monocytes, macrophages, dendritic cells, lymphocytes, and bone marrow cells [31, 32]. A3AR plays an unclear role in inflammatory processes, as it has been shown to have both anti-inflammatory and pro-inflammatory activites [33, 34]. A3AR stimulation on mouse mast cells has been shown to induce degranulation [35]. Conversely, in eosinophils it inhibits chemotaxis, degranulation and generation of superoxide anion [36, 37]. In monocytes and macrophages, A3AR inhibits the TNF-α release through the NF-κB signal transduction pathway [38]. In neutrophils, it promotes chemotaxis and inhibits the superoxide anion generation [39]. Furthermore, overexpression of this receptor has been reported in the lungs of patient affected by airway inflammation [40].
A3AR intracellular signaling transduction
A3AR can interact with different G proteins, including Gi, Gq, and Go [41]. Specifically, interaction between A3AR and Gi protein inhibits adenylate cyclase activity, thus leading to a decrease in cAMP (Fig. 1) [41]. This process causes protein kinase A (PKA) inhibition, which leads to a glycogen synthase kinase-3β (GSK-3β) increase, a β-catenin and cyclin D1 down-regulation, as well as NF-κB-DNA binding potential reduction [41]. Furthermore, A3AR regulates the mitogen-activated protein kinase (MAPK), phosphatidylinositol 3-kinase (PI3K)/Akt and NF-κB signaling pathways [29]. It is known that PKA/PKB/Atk phosphorylate and inactivate GSK-3β, which is a key element of the Wnt signal. In its active form, GSK-3β suppresses cell proliferation [42]. When A3AR is activated, a decrease in cAMP levels occurs thereby leading to a reduction in the phosphorylated PKB/Akt and PKA active form [42]. This phenomenon causes a dysregulation in the Wnt signal transduction pathway, which increases cell proliferation and, thus, tumorigenesis [28]. When A3AR activates G proteins, phospholipase C activity is stimulated with a consequent increase in calcium concentration and protein kinase C (PKC) stimulation. This process induces TNF-α release in active macrophages [4].
The role of A3AR in cancer
General overwiew
Cancer can be a deadly disease, which is caused by alterations in gene expression and pathways [43, 44]. A number of useful diagnostic/prognostic markers have been identified for numerous tumor types [44, 45]. A3AR is overexpressed in cancer and it is considered a tumor diagnostic/prognostic marker, as previous studies have demonstrated its overexpression in different malignant tumors including melanoma, breast, prostate, liver, pancreatic and lung cancers, as well as lymphoma, glioblastoma and malignant pleural mesothelioma (MPM) [3, 29, 46, 47]. High levels of A3AR in cancer cells and blood cells have also been demonstrated in colorectal cancer patients [47, 48].
A3AR stimulation is known to inhibit tumor growth by regulating the Wnt pathway [49]. GSK-3β plays a key role in this molecular process, as it is responsible for β-catenin phosphorylation. When phosphorylated, β-catenin induces the transcription of genes, which are fundamental for the cell cycle progression, such as c-myc and cyclin D1. By treating cancer cells with A3AR agonists, GSK-3β levels increase, while cyclin D1 and c-myc expression is suppressed. This molecular effect, which induces a decrease in cancer cell proliferation [28], has been found in melanoma, hepatocellular carcinoma, as well as in the synoviocytes of patients suffering from rheumatoid arthritis [4].
The dual nature of A3AR in cancer has been remarked upon (Fig. 2). In some tumors it promotes cell proliferation and survival, while in others it triggers cytostatic and apoptotic pathways [28]. A3AR stimulation inhibits lung cancer proliferation by arresting the cell cycle [50]. A similar effect has also been observed in vitro in murine lymphoma [51]. Adenosine-dependent A3AR stimulation induces apoptosis in stomach cancer cells via a mechanism that involves PKC activation [52]. An inhibition of tumor growth following receptor stimulation has been demonstrated in lymphoma [51], leukemia, [53] as well as colon [54] and pancreatic carcinoma [55]. Contrariwise, A3AR stimulation prompts cell proliferation in other cancer types, such as colorectal cancer and adenocarcinoma [56]. In human glioblastoma cells, A3ARs stimulation induces an increase in MMP-9 following ERK, PKA/Akt activation, causing an increase in cell invasiveness [46, 52, 57].
Hypoxia and A3AR regulation
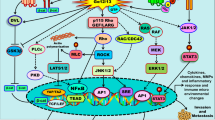
Hypoxia is a common phenomenon in many solid tumors [58]. In hypoxia conditions, where there is a lack of O2, adenosine accumulates in TME, while factors involved in cellular response to hypoxia, such as hypoxia inducible factor 1 (HIF-1) are released [59]. HIF-1 is abundant in tumor cells surrounded by TME, where it plays a fundamental role in angiogenesis, invasion as well as in the alteration of tumor cell metabolism [60, 61]. HIF-1 is a heterodimer protein made up of an α and a β subunits, which are HIF-1α and HIF-1β. Although both subunits are constitutively expressed, the expression level of α subunit increases when O2 concentrations decrease [62]. Since both HIF-1 and A3AR have been found to be overexpressed in cancer, a link between A3AR stimulation and the modulation of HIF-1α expression in hypoxic conditions has been explored [59, 63]. In some tumor types, such as melanoma, glioblastoma and colon cancer, HIF-1α expression has been found to be increased by adenosine-induced Atk and/or MAPK signal pathway activation [59]. Adenosine involvement in the production of vascular endothelial growth factor (VEGF) in cancer has also been investigated. In vitro evidence indicates that in glioblastoma cell lines, A3AR activation stimulates VEGF expression [64]. Another study has reported that treating colon cancer cell lines with A3AR antagonists blocks both HIF-1α and VEGF expression in hypoxic conditions [12]. A similar effect has also been observed in melanoma cell lines, where adenosine stimulated A3AR causing HIF-1α over-expression in response to hypoxia [59]. To demonstrate that the HIF-1α increase depends on A3AR stimulation, whereas it is not due to other ARs, antagonists from other AR family members, including A1R, A2AR, A2BR, were used. The positive effect of adenosine in increasing HIF-1α was only determined following A3AR stimulation. In the same study, p44/p42 and p38 MAPK were demonstrated as necessary in order to increase HIF-1α levels (Fig. 3) [59].

Cellular pathways activated by the stimulation of the A3 receptor in normoxia and hypoxia conditions.
Additional data support a correlation between A3AR and MAPK/ERK pathway [65]. A3AR-mediated ERK activation has been reported in human fetal astrocytes, microglia and several tumors, such as colon carcinoma, glioblastoma, and melanoma [59, 66,67,68]. On the contrary, ERK inhibition leads to a reduction in cell proliferation in melanoma, prostate cancer and glioma [67, 68]. Different in vitro models, such as hamster ovary cells [69], melanoma [59], colon carcinoma and glioblastoma [56, 70], indicated that A3AR is also responsible for p38 MAPK activation, while the opposite result has been reported in synoviocyte cultures [71]. A3AR is also responsible for activating C-Jun N-terminal kinase patway in microglia and glioblastoma cells [4]. As a result, this mechanism causes an increase in cell migration and matrix metallopeptidase-9 (MMP-9) overexpression [4].
A3AR stimulation causes Akt phosphorylation [65]. In glioblastoma cells and in mouse basophilic cancer cells Akt phosphorylation causes apoptosis inhibition [70, 72], while the same pathway demonstrates an anti-proliferative effect in human melanoma cells [73]. A3AR also mediates PI3K/Akt signal activation [65]. The PI3K/Akt and NF-κB signal transduction pathways are mediators of the anti-inflammatory effect, which has been observed in BV-2 microglial cells [74], monocytes and mesothelioma cells [46, 75]. PI3K/Akt and NF-κB pathway inhibition reduces HIF-α and GSK-3β concentration (Fig. 4) [4, 25].

Representation of signal transduction pathways induced by the stimulation of the A3 receptor.
Animal models indicate that adenosine plays a key role in regulating vascularization in melanoma [76]. Specific activation of A1AR, A2AR and A3AR in CD73-knockout mice affected melanoma growth, neovascularization, angiogenesis, and macrophage infiltration. Specifically, A3AR activation induced pro-angiogenic factor expression and secretion by mast cells, macrophages infiltration and cytokines expression in TME via a paracrine mechanism [76]. In another study conducted on a human melanoma cell line, it has been reported that A3AR stimulation can induce a reduction in cell proliferation [73]. Indeed, after receptor stimulation, the PI3K/Akt signal transduction pathway is activated, leading to a reduction in ERK1/2 [73]. This molecular effect ultimately inhibits cell proliferation [73].
A3AR agonists and antagonists in cancer treatment
General overview
A3AR offers a promising therapeutic target for inflammatory diseases, such as rheumatoid arthritis and psoriasis [77]. Its importance in treating cancer is also increasing [78]. Numerous agonists, partial agonists, allosteric modulators and antagonists have been developed [79]. The main drugs employed in clinical trials are N6-(3-Iodobenzyl)-adenosine-5′-N-methyluronamide (IB-MECA; CF101) and 2-chloro-N6-(3-iodobenzyl)-adenosine-5′-N-methyluronamide (Cl-IB-MECA; CF102) (Fig. 5). These molecules, which have shown positive results in preclinical studies, are considered safe and have been well tolerated during clinical trials [80,81,82,83]. IB-MECA and Cl-IB-MECA derive from adenosine and contain a lipophilic substituent (3-iodobenzyl) in position N6 and modified ribose in position 5′ [84]. Cl-IB-MECA contains a further substituent, chlorine, which makes it more selective than IB-MECA (Fig. 5). The first developed selective agonist IB-MECA is 50 times more selective for A3AR than other ARs [84]. However, in addition to affinity, other parameters must also be considered, such as the half-life of the agonist, the duration of the response induced by the receptor link and efficacy in vivo. Additional molecules, derived from other nucleosides, act as partial agonists or antagonists [4]. Both have been shown to be safe and effective. Currently, IB-MECA is being tested in phase I clinical trial for psoriasis (NCT00428974) and phase II and III clinical trials for rheumatoid arthritis (NCT01034306 and NCT02647762), while Cl-IB-MECA is being tested in phase I and II clinical trials for liver cancer (NCT00790218 and NCT02128958) and in the treatment of non-alcoholic steatohepatitis (NASH, NCT02927314). Both IB-MECA and Cl-IB-MECA, which are administered orally, are safe and well tolerated.

Chemical structures of (A) N6-(3-Iodobenzyl)adenosine-5′-N-methyluronamide (IB- MECA; CF101); B 2-chloro-N6-(3- iodobenzyl)-adenosine-5′-N-methyluronamide (Cl-IB-MECA; CF102).
In vivo experiments indicate that A3AR is not desensitized following chronic treatment [85]. Other data indicate that shortly after its last administration, A3AR down-regulation occurs, while 24 h after the last administration of the drug, expression levels of the receptor return to those shown by the control group. Therefore, chronic treatment does not reduce A3AR expression [85].
The antitumor effect of A3AR agonists occurs via Wnt/NF-κB pathway modulation [34]. PKB/Akt, NF-κB and TNF-α expression levels have been shown to be reduced during treatment with A3AR agonists in both in vitro/in vivo liver carcinoma models [86]. Based on reported data, synthetic A3AR agonists induce apoptosis and inhibit cell proliferation in different types of cancer cells in in vitro/in vivo models. These drugs are safe and well tolerated, mainly due to their cardio- and neuro-protective effects and therefore, they can be considered as new therapeutic approaches to some cancer types.
A3AR agonist: IB-MECA
The antitumor effects of IB-MECA have been associated with GSK-3β up-regulation and NF-κB, cyclin D and c-Myc down-regulation [87]. Furthermore, colon cancer animal models have indicated that A3AR activation via IB-MECA interaction inhibits tumor growth. Indeed, in tumor tissues taken from IB-MECA-treated mice, a decrease in the level of PKA expression and an increase in GSK-3β, which led to the down-regulation of β-catenin, has been observed [54]. Subsequent in vitro studies revealed that this effect is hampered by administrating GSK-3β inhibitors, which confirms the involvement of A3AR [54]. IB-MECA induces down-regulation of the PKB/Akt signal transduction pathway, consequently inhibiting NF-κB activity in vivo/in vitro [54]. IB-MECA also prevents the formation of liver metastases from colon cancer cells inoculated into the spleen [25]. In addition, IB-MECA significantly blocks breast cancer cell motility and metastasis [88]. IB-MECA is also responsible for prostate cancer cell proliferation suppression. At low concentrations IB-MECA arrests the cell cycle in phase G1, while at a high concentrations it induces apoptosis by increasing the activity of pro-apoptotic proteins caspase-3 (CASP3) and Bax, while Bcl-2 expression levels decrease [89]. However, IB-MECA also blocks cell cycle progression in phase G1 at concentrations between 0.001 and 10 µM in ovarian cancer cell lines, where it also causes a decrease in cyclin D1 and cyclin-dependent kinase 4 levels [90].
A3AR agonist: Cl-IB-MECA
Cl-IB-MECA pharmacokinetic parameters and long-term safety have been analyzed in a study involving hepatocellular carcinoma patients. Cl-IB-MECA-induced apoptosis has been reported to occur by modulating the Wnt signal transduction pathway, while showing a protective effect on healthy cells. These promising results suggest that this compound could also be used on patients with other diseases, such as liver cirrhosis or inflammation [91]. Cl-IB-MECA is able to inhibit lung metastases formation in mice with melanoma at a nanomolar concentration range [27]. In addition, Cl-IB-MECA shows a synergistic antitumor effect when employed in combination with cyclophosphamide [27]. A study conducted on MPM indicates a key role for Cl-IB-MECA as an anticancer agent [46]. This neoplasm is strongly correlated to asbestos exposure. Evidence also suggests the involvement of Simian Virus 40, which is an oncogenic virus member of the Polyomaviridae family [92, 93], as an MPM co-factor [94]. During MPM onset, macrophages secrete TNF-α, which activates pro-inflammatory pathways, leading to NF-κB activation, thereby promoting survival. In inflammatory conditions, adenosine concentrations increases, as A3AR have been shown to be involved in the NF-κB/Akt pathway. When tumor cells are treated with Cl-IB-MECA, the level of Akt phosphorylation decreases [42, 65]. Cl-IB-MECA is able to inhibit the effect of TNF-α on the survival/proliferation of cells exposed to asbestos, by inhibiting NF-κB activation [46]. This treatment induces lactate dehydrogenase and CASP3 release in MPM cells, implying a cytotoxic effect mediated by A3AR [46]. These findings suggest that A3AR could represent a target for preventing MPM development.
Cl-IB-MECA can be effective against other cancer types. Data from lung cancer cell lines experiments indicated that 0.01–10 mM of both adenosine and Cl-IB-MECA induces apoptosis in a dose-dependent manner [42]. A study conducted on thyroid carcinoma reported that Cl-IB-MECA inhibits tumor growth and blocks cell cycle progression in tumor cells [95]. This effect is related to a reduction in cyclin D1 expression and the dephosphorylation of ERK1/2, which both depend on treatment time and drug concentration. In addition, in vivo Cl-IB-MECA potentiates the activity of NK cells by inducing IL-12 production, a cytotoxic factor with antitumor effects [57].
A3AR has been found overexpressed in tumor cells and peripheral blood mononuclear cells in patients with hepatocellular carcinoma [48]. Treatment with Cl-IB-MECA inhibits the growth of hepatocellular carcinoma cells in orthopic mouse models in a dose-dependent manner. Moreover, the effect of Cl-IB-MECA is due to NF-κB down-regulation and an increase in GSK-3β [86]. Induced apoptosis also depends on an increase in the expression of pro-apoptotic proteins, such as Bad, BAX and casapase-3 [86].
A3AR agonist: Thio-Cl-IB-MECA
Using agonists as antitumor agents, such as 2-chloro-N6-(3-iodobenzyl)−4′-thioadenosine-5′-N-methyluronamide (thio-Cl-IB-MECA), has also brought encouraging results. Thio-Cl-IB-MECA is able to inhibit the cell cycle progression in lung cancer cells [50]. It can also induce apoptosis at a high concentrations via cyclin D1, c-Myc and cyclin-dependent kinase 4 (CDK4) downregulation as well as CASP3 and −9 activation [50]. Thio-Cl-IB-MECA induces apoptosis in human leukemic cells and has an anti-inflammatory effect, as it inhibits the expression of pro-inflammatory cytokines by modulating PKB/Akt pathways and NF-κB [50, 57, 96].
A3AR antagonists
Results on receptor stimulation support the hypothesis that receptor antagonists may also be useful in treating different cancer types [97]. Animal models have confirmed that activating the receptor in melanoma cells induces an increase in blood vessel density, pro-angiogenic molecule secretion, cytokine production and the invasion of macrophages into the tumor [4]. In glioblastoma, A3AR stimulation prompts MMP-9 expression with a consequent increase in cell invasiveness, while using antagonists co-adiuvates the antitumor effect of chemotherapy [70]. These results have been obtained following in vitro studies, but studies conducted on animal models are needed to support these data and confirm the idea that antagonists may also represent a valid therapeutic approach [4, 29]. A3AR antagonists employed in clinical trials include PBF-1650 (NCT03798236) phase I for Psoriasis and PBF-677 (NCT02639975, Glaucoma, phase I) (NCT03773952, Ulcerative Colitis, phase II).
Concluding remarks and future perspectives
In conclusion, the studies reported in this review indicate A3AR and its ligands as key players in cancer onset/development. Previous in vitro/in vivo data indicate that A3AR has been found as overexpressed in a variety of cancer types, including glioblastoma/glioma, colon, breast, and pancreatic cancers, as well as MPM and lymphoma. A3AR could thus potentially be used as a tumor diagnostic/prognostic marker and target for anticancer therapy. A3AR has become an attractive therapeutic antitumor target, as its agonists show encouraging results in preclinical studies. Furthermore, some of those agonists are also currently being tested in clinical trials, while A3AR antagonists have also obtained encouraging results in preclinical studies.
Research focused on the role of A3AR and its modulators in cancer is an essential future area of study. As A3AR appears to be enigmatic in terms of its effects, investigating the dual nature of this receptor is an important research field, which deserves attention. Although a growing number of studies have investigated the structure and function of A3AR, its regulative role on modulating both cell proliferation and cell death by interacting with its large variety of ligands, represents a relatively well debated issue due to its dual role depending on a tissue specific context and on ligands type/concentration [26]. On this ground, further studies focusing on the dual nature of A3AR activation, as well as the potential interaction of these two opposing responses to tumor growth, are needed. Since A3AR stimulation activates a number of pathways, the study of these mechanisms could be essential for understanding the role of these receptors in cancer onset/progression and metastasis. Further in vitro/in vivo studies into A3AR mechanisms upon cancer are, thus, also to be encouraged. Novel data could improve cancer diagnosis, the prognostic management of cancer patients, as well as the development of novel therapies.
References
Di Virgilio F, Adinolfi E. Extracellular purines, purinergic receptors and tumor growth. Oncogene. 2017. https://doi.org/10.1038/onc.2016.206.
Contini C, Rotondo JC, Magagnoli F, Maritati M, Seraceni S, Graziano A, et al. Investigation on silent bacterial infections in specimens from pregnant women affected by spontaneous miscarriage. J Cell Physiol. 2018;234:100–7.
Kazemi MH, Raoofi Mohseni S, Hojjat-Farsangi M, Anvari E, Ghalamfarsa G, Mohammadi H et al. Adenosine and adenosine receptors in the immunopathogenesis and treatment of cancer. J Cell Physiol. 2018. https://doi.org/10.1002/jcp.25873.
Jacobson KA, Merighi S, Varani K, Borea PA, Baraldi S, Aghazadeh Tabrizi M et al. A3 Adenosine Receptors as Modulators of Inflammation: from Medicinal Chemistry to Therapy. Med Res Rev. 2018. https://doi.org/10.1002/med.21456.
Phillis JW, O’Regan MH, Perkins LM. Measurement of rat plasma adenosine levels during normoxia and hypoxia. Life Sci. 1992;51. https://doi.org/10.1016/0024-3205(92)90363-T.
Löfgren L, Pehrsson S, Hägglund G, Tjellström H, Nylander S. Accurate measurement of endogenous adenosine in human blood. PLoS ONE 2018;13. https://doi.org/10.1371/journal.pone.0205707.
Hajizadeh F, Masjedi A, Heydarzedeh Asl S, Karoon Kiani F, Peydaveisi M, Ghalamfarsa G, et al. Adenosine and adenosine receptors in colorectal cancer. Int Immunopharmacol. 2020;87:106853.
Hershfield MS, Seegmiller JE. Regulation of de novo purine biosynthesis in human lymphoblasts. Coordinate control of proximal (rate-determining) steps and the inosinic acid branch point. J Biol Chem. 1976;251:7348–54.
Pastor-Anglada M, Pérez-Torras S. Who is who in Adenosine transport. Front Pharmacol. 2018. https://doi.org/10.3389/fphar.2018.00627.
Görlach A Control of adenosine transport by hypoxia. Circ Res. 2005. https://doi.org/10.1161/01.RES.0000174112.36064.77.
Joyce JA, Fearon DT. T cell exclusion, immune privilege, and the tumor microenvironment. Science (80-.). 2015. https://doi.org/10.1126/science.aaa6204.
Merighi S, Benini A, Mirandola P, Gessi S, Varani K, Leung E et al. Adenosine modulates vascular endothelial growth factor expression via hypoxia-inducible factor-1 in human glioblastoma cells. Biochem Pharmacol. 2006. https://doi.org/10.1016/j.bcp.2006.03.020.
Rotondo JC, Giari L, Guerranti C, Tognon M, Castaldelli G, Fano EA, et al. Environmental doses of perfluorooctanoic acid change the expression of genes in target tissues of common carp. Environ Toxicol Chem. 2018;37:942–8.
Fishman P, Bar-Yehuda S, Barer F, Madi L, Multani AS, Pathak S. The A3 adenosine receptor as a new target for cancer therapy and chemoprotection. Exp Cell Res. 2001;269:230–6.
Effendi WI, Nagano T, Kobayashi K, Nishimura Y. Focusing on Adenosine Receptors as a Potential Targeted Therapy in Human Diseases. Cells 2020. https://doi.org/10.3390/cells9030785.
Micioni Di Bonaventura MV, Pucci M, Giusepponi ME, Romano A, Lambertucci C, Volpini R et al. Regulation of adenosine A2A receptor gene expression in a model of binge eating in the amygdaloid complex of female rats. J Psychopharmacol. 2019. https://doi.org/10.1177/0269881119845798.
Rotondo JC, Bosi S, Bazzan E, Di Domenico M, De Mattei M, Selvatici R, et al. Methylenetetrahydrofolate reductase gene promoter hypermethylation in semen samples of infertile couples correlates with recurrent spontaneous abortion. Hum Reprod. 2012;27:3632–8.
Rotondo JC, Selvatici R, Di Domenico M, Marci R, Vesce F, Tognon M, et al. Methylation loss at H19 imprinted gene correlates with methylenetetrahydrofolate reductase gene promoter hypermethylation in semen samples from infertile males. Epigenetics. 2013;8:990–7.
Rotondo JC, Oton-Gonzalez L, Selvatici R, Rizzo P, Pavasini R, Campo GC, et al. SERPINA1 Gene Promoter Is Differentially Methylated in Peripheral Blood Mononuclear Cells of Pregnant Women. Front Cell Dev Biol. 2020;8:5505.
Rotondo JC, Borghi A, Selvatici R, Magri E, Bianchini E, Montinari E et al. Hypermethylation-induced inactivation of the IRF6 gene as a possible early event in progression of vulvar squamous cell carcinoma associated with lichen sclerosus. JAMA Dermatol. 2016;152. https://doi.org/10.1001/jamadermatol.2016.1336.
Stomper J, Rotondo JC, Greve G, Lübbert M. Hypomethylating agents (HMA) for the treatment of acute myeloid leukemia and myelodysplastic syndromes: mechanisms of resistance and novel HMA-based therapies. Leukemia. 2021;35:1873–89. https://doi.org/10.1038/s41375-021-01218-0.
Lanzillotti C, De Mattei M, Mazziotta C, Taraballi F, Rotondo JC, Tognon M, et al. Long Non-coding RNAs and MicroRNAs Interplay in Osteogenic Differentiation of Mesenchymal Stem Cells. Front Cell Dev Biol. 2021;9:646032.
Rotondo JC, Lanzillotti C, Mazziotta C, Tognon M, Martini F. Epigenetics of male infertility: the role of DNA methylation. Front Cell Dev Biol. 2021;9:689624.
Fishman P, Bar-Yehuda S, Liang BT, Jacobson KA. Pharmacological and therapeutic effects of A3 adenosine receptor agonists. Drug Discov Today 2012;17:359–66.
Borea PA, Varani K, Vincenzi F, Baraldi PG, Tabrizi MA, Merighi S et al. The a3 adenosine receptor: History and perspectives. Pharmacol Rev. 2015. https://doi.org/10.1124/pr.113.008540.
Mlejnek P, Dolezel P, Frydrych I. Effects of synthetic A3 adenosine receptor agonists on cell proliferation and viability are receptor independent at micromolar concentrations. J Physiol Biochem. 2013;69:405–17.
Marwein S, Mishra B, De UC, Acharya PC. Recent Progress of Adenosine Receptor Modulators in the Development of Anticancer Chemotherapeutic Agents. Curr Pharm Des. 2019. https://doi.org/10.2174/1381612825666190716141851.
Gorain B, Choudhury H, Yee GS, Bhattamisra SK. Adenosine Receptors as Novel Targets for the Treatment of Various Cancers. Curr Pharm Des. 2019. https://doi.org/10.2174/1381612825666190716102037.
Borea PA, Gessi S, Merighi S, Vincenzi F, Varani K. Pharmacology of adenosine receptors: the state of the art. Physiol Rev. 2018. https://doi.org/10.1152/physrev.00049.2017.
Klaasse EC, IJzerman AP, de Grip WJ, Beukers MW. Internalization and desensitization of adenosine receptors. Purinergic Signal. 2008;4:21–37.
Borea PA, Gessi S, Merighi S, Varani K. Adenosine as a Multi-Signalling Guardian Angel in Human Diseases: When, Where and How Does it Exert its Protective Effects? Trends Pharmacol. Sci. 2016. https://doi.org/10.1016/j.tips.2016.02.006.
Przybyla T, Sakowicz-Burkiewicz M, Pawelczyk T. Purinergic signaling in B cells. Acta Biochim Pol. 2018;65:1–7.
Bar-Yehuda S, Silverman MH, Kerns WD, Ochaion A, Cohen S, Fishman P. The anti-inflammatory effect of A3 adenosine receptor agonists: a novel targeted therapy for rheumatoid arthritis. Expert Opin Investig Drugs. 2007. https://doi.org/10.1517/13543784.16.10.1601.
Ren T, Tian T, Feng X, Ye S, Wang H, Wu W, et al. An adenosine A3 receptor agonist inhibits DSS-induced colitis in mice through modulation of the NF-κB signaling pathway. Sci Rep. 2015;5:9047.
Salvatore C, Tilley S, Latour A, Fletcher D, Koller B, Jacobson M. Disruption of the A(3) adenosine receptor gene in mice and its effect on stimulated inflammatory cells. J Biol Chem. 2000;275:4429–34.
da Rocha Lapa F, Macedo-Júnior SJ, Luiz Cerutti M, Santos ARS. Pharmacology of Adenosine Receptors and Their Signaling Role in Immunity and Inflammation. In: Pharmacology and Therapeutics. 2014. https://doi.org/10.5772/57206.
Ezeamuzie CI, Philips E. Adenosine A3 receptors on human eosinophils mediate inhibition of degranulation and superoxide anion release. Br J Pharmacol. 1999. https://doi.org/10.1038/sj.bjp.0702476.
Ren T, Qiu Y, Wu W, Feng X, Ye S, Wang Z et al. Activation of adenosine A3 receptor alleviates TNF-α-induced inflammation through inhibition of the NF-κB signaling pathway in human colonic epithelial cells. Mediators Inflamm. 2014; 2014. https://doi.org/10.1155/2014/818251.
Rose F, Hirschhorn R, Weissmann G, Cronstein B. Adenosine promotes neutrophil chemotaxis. J Exp Med. 1988;167:1186–94.
Gessi S, Merighi S, Varani K, Leung E, Mac Lennan S, Borea PA. The A3 adenosine receptor: an enigmatic player in cell biology. Pharmacol Ther. 2008. https://doi.org/10.1016/j.pharmthera.2007.09.002.
Gorzalczany Y, Sagi-Eisenberg R. Role of mast cell-derived adenosine in cancer. Int J Mol Sci. 2019. https://doi.org/10.3390/ijms20102603.
Fishman P, Bar-Yehuda S, Synowitz M, Powell JD, Klotz KN, Gessi S, et al. Adenosine receptors and cancer. Handb Exp Pharm. 2009;193:399–441.
Tognon M, Luppi M, Corallini A, Taronna A, Barozzi P, Rotondo JC, et al. Immunologic evidence of a strong association between non-Hodgkin lymphoma and simian virus 40. Cancer. 2015;121:2618–26.
Preti M, Rotondo JC, Holzinger D, Micheletti L, Gallio N, Robitaille A, et al. Role of human papillomavirus infection in the etiology of vulvar cancer in Italian women. Infect Agent Cancer. 2020;15:e2020.
Rotondo JC, Oton-Gonzalez L, Mazziotta C, Lanzillotti C, Iaquinta MR, Tognon M, et al. Simultaneous detection and viral DNA load quantification of different human papillomavirus types in clinical specimens by the high analytical droplet digital PCR method. Front Microbiol. 2020;11:591452.
Varani K, Maniero S, Vincenzi F, Targa M, Stefanelli A, Maniscalco P, et al. A3 receptors are overexpressed in pleura from patients with mesothelioma and reduce cell growth via Akt/nuclear factor-κB pathway. Am J Respir Crit Care Med. 2011;183:522–30.
Gessi S, Cattabriga E, Avitabile A, Gafa R, Lanza G, Cavazzini L et al. Elevated expression of A3 adenosine receptors in human colorectal cancer is reflected in peripheral blood cells. Clin Cancer Res. 2004. https://doi.org/10.1158/1078-0432.CCR-1134-03.
Bar-Yehuda S, Stemmer SM, Madi L, Castel D, Ochaion A, Cohen S et al. The A3 adenosine receptor agonist CF102 induces apoptosis of hepatocellular carcinoma via de-regulation of the Wnt and NF-κB signal transduction pathways. Int J Oncol. 2008. https://doi.org/10.3892/ijo_00000008.
Gorain B, Choudhury H, Yee GS, Bhattamisra SK. Adenosine Receptors as Novel Targets for the Treatment of Various Cancers. Curr Pharm Des. 2019;25:2828–41.
Kim SJ, Min HY, Chung HJ, Park EJ, Hong JY, Kang YJ, et al. Inhibition of cell proliferation through cell cycle arrest and apoptosis by thio-Cl-IB-MECA, a novel A3 adenosine receptor agonist, in human lung cancer cells. Cancer Lett. 2008;264:309–15.
Fishman P, Bar-Yehuda S, Ohana G, Pathak S, Wasserman L, Barer F, et al. Adenosine acts as an inhibitor of lymphoma cell growtha major role for the A3 adenosine receptor. Eur J Cancer. 2000;36:1452–8.
Tsuchiya A, Nishizaki T. Anticancer effect of adenosine on gastric cancer via diverse signaling pathways. World J Gastroenterol. 2015. https://doi.org/10.3748/wjg.v21.i39.10931.
Vaisitti T, Arruga F, Deaglio S. Targeting the adenosinergic axis in chronic lymphocytic leukemia: a way to disrupt the tumor niche? Int J Mol Sci. 2018;19:1167.
Fishman P, Bar-Yehuda S, Ohana G, Barer F, Ochaion A, Erlanger A, et al. An agonist to the A3 adenosine receptor inhibits colon carcinoma growth in mice via modulation of GSK-3β and NF-κB. Oncogene. 2004;23:2465–71.
Roth S, Hackert T. Adenosine in pancreatic cancer: Emerging combination therapies. EBioMedicine. 2019;48:20–1.
Gessi S, Merighi S, Varani K, Cattabriga E, Benini A, Mirandola P, et al. Adenosine receptors in colon carcinoma tissues and colon tumoral cell lines: Focus on the A3 adenosine subtype. J Cell Physiol. 2007;211:826–36.
Gessi S, Merighi S, Sacchetto V, Simioni C, Borea PA. Adenosine receptors and cancer. Biochim. Biophys. Acta - Biomembr. 2011. https://doi.org/10.1016/j.bbamem.2010.09.020.
Karakashev SV, Reginato MJ. Progress toward overcoming hypoxia-induced resistance to solid tumor therapy. Cancer Manag Res. 2015;12:253–64.
Merighi S, Benini A, Mirandola P, Gessi S, Varani K, Leung E et al. A3 adenosine receptors modulate hypoxia-inducible factor-1α expression in human A375 melanoma cells. Neoplasia. 2005. https://doi.org/10.1593/neo.05334.
Nigim F, Cavanaugh J, Patel AP, Curry WT, Esaki SI, Kasper EM, et al. Targeting Hypoxia-Inducible Factor 1α in a New Orthotopic Model of Glioblastoma Recapitulating the Hypoxic Tumor Microenvironment. J Neuropathol Exp Neurol. 2015;74:710–22.
Jin X, Dai L, Ma Y, Wang J, Liu Z. Implications of HIF-1α in the tumorigenesis and progression of pancreatic cancer. Cancer Cell Int. 2020;24:273.
Wang GL, Jiang BH, Rue EA, Semenza GL. Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS heterodimer regulated by cellular O2 tension. Proc Natl Acad Sci USA. 1995;92:5510–4.
Maugeri G, D’Amico AG, Federico C, Saccone S, Giunta S, Cavallaro S, et al. Involvement of A3 Adenosine Receptor in Neuroblastoma Progression via Modulation of the Hypoxic/Angiogenic Pathway. J Mol Neurosci. 2019;69:166–76.
Allard B, Cousineau I, Allard D, Buisseret L, Pommey S, Chrobak P, et al. Adenosine A2a receptor promotes lymphangiogenesis and lymph node metastasis. Oncoimmunology. 2019;26:1601481.
Merighi S, Benini A, Mirandola P, Gessi S, Varani K, Leung E, et al. Modulation of the Akt/Ras/Raf/MEK/ERK pathway by A3 adenosine receptor. Purinergic Signal. 2006;2:267–32.
Hammarberg C, Schulte G, Fredholm BB. Evidence for functional adenosine A3 receptors in microglia cells. J Neurochem. 2003. https://doi.org/10.1046/j.1471-4159.2003.01919.x.
Merighi S, Mirandola P, Milani D, Varani K, Gessi S, Klotz KN et al. Adenosine receptors as mediators of both cell proliferation and cell death of cultured human melanoma cells. J Investig Dermatol. 2002. https://doi.org/10.1046/j.1523-1747.2002.00111.x.
Jajoo S, Mukherjea D, Watabe K, Ramkumar V. Adenosine A3 receptor suppresses prostate cancer metastasis by inhibiting NADPH oxidase activity. Neoplasia. 2009;11:1132–45.
Schulte G, Fredholm BB. Human adenosine A1, A(2A), A(2B), and A3 receptors expressed in Chinese hamster ovary cells all mediate the phosphorylation of extracellular-regulated kinase 1/2. Mol Pharm. 2000;58:477–82.
Gessi S, Sacchetto V, Fogli E, Merighi S, Varani K, Baraldi PG, et al. Modulation of metalloproteinase-9 in U87MG glioblastoma cells by A3 adenosine receptors. Biochem Pharm. 2010;79:1483–95.
Varani K, Vincenzi F, Tosi A, Targa M, Masieri FF, Ongaro A, et al. Expression and functional role of adenosine receptors in regulating inflammatory responses in human synoviocytes. Br J Pharm. 2010;160:101–15.
Merighi S, Benini A, Mirandola P, Gessi S, Varani K, Leung E et al. Hypoxia inhibits paclitaxel-induced apoptosis through adenosine-mediated phosphorylation of bad in glioblastoma cells. Mol Pharmacol. 2007. https://doi.org/10.1124/mol.106.031849.
Merighi S, Benini A, Mirandola P, Gessi S, Varani K, Leung E, et al. A3 adenosine receptor activation inhibits cell proliferation via phosphatidylinositol 3-kinase/Akt-dependent inhibition of the extracellular signal-regulated kinase 1/2 phosphorylation in A375 human melanoma cells. J Biol Chem. 2005;280:19516–26.
Lee JY, Jhun BS, Oh YT, Lee JH, Choe W, Baik HH et al. Activation of adenosine A3 receptor suppresses lipopolysaccharide-induced TNF-α production through inhibition of PI 3-kinase/Akt and NF-κB activation in murine BV2 microglial cells. Neurosci Lett. 2006. https://doi.org/10.1016/j.neulet.2005.11.004.
Madi L, Cohen S, Ochayin A, Bar-Yehuda S, Barer F, Fishman P. Overexpression of A3 adenosine receptor in peripheral blood mononuclear cells in rheumatoid arthritis: Involvement of nuclear factor-κB in mediating receptor level. J Rheumatol. 2007.
Koszałka P, Gołuńska M, Urban A, Stasiłojć G, Stanisławowski M, Majewski M et al. Specific activation of A3, A2a and A1 adenosine receptors in CD73-knockout mice affects B16F10 melanoma growth, neovascularization, angiogenesis and macrophage infiltration. PLoS ONE. 2016. https://doi.org/10.1371/journal.pone.0151420.
Fishman P, Cohen S. The A3 adenosine receptor (A3AR): therapeutic target and predictive biological marker in rheumatoid arthritis. Clin Rheumatol. 2016;35:2359–62.
Mailavaram RP, Al-Attraqchi OHA, Kar S, Ghosh S. Current Status in the Design and Development of Agonists and Antagonists of Adenosine A3 Receptor as Potential Therapeutic Agents. Curr Pharm Des. 2019;25:2772–87.
Jacobson KA, Tosh DK, Jain S, Gao ZG. Historical and current adenosine receptor agonists in preclinical and clinical development. Front Cell Neurosci. 2019;13:124.
Montinaro A, Forte G, Sorrentino R, Luciano A, Palma G.ArraC, et al. Adoptive Immunotherapy with Cl-IB-MECA-Treated CD8. T Cells Reduces Melanoma Growth in Mice. PLoS ONE. 2012;7:e45401. https://doi.org/10.1371/journal.pone.0045401.
Abedi H, Aghaei M, Panjehpour M, Hajiahmadi S. Mitochondrial and caspase pathways are involved in the induction of apoptosis by IB-MECA in ovarian cancer cell lines. Tumor Biol. 2014. https://doi.org/10.1007/s13277-014-2396-9.
Taliani S, La Motta C, Mugnaini L, Simorini F, Salerno S, Marini AM et al. Novel N 2-Substituted pyrazolo[3,4- d]pyrimidine adenosine A3 receptor antagonists: Inhibition of A3-Mediated human glioblastoma cell proliferation. J Med Chem. 2010. https://doi.org/10.1021/jm901785w.
Kim TH, Kim YK, Woo JS. The adenosine A3 receptor agonist Cl-IB-MECA induces cell death through Ca2+/ROS-dependent down regulation of ERK and akt in A172 human glioma cells. Neurochem Res. 2012. https://doi.org/10.1007/s11064-012-0855-5.
Suresh RR, Jain S, Chen Z, Tosh DK, Ma Y, Podszun MC, et al. Design and in vivo activity of A3 adenosine receptor agonist prodrugs. Purinergic Signal. 2020;16:367–77.
Varani K, Vincenzi F, Targa M, Paradiso B, Parrilli A, Fini M, et al. The stimulation of A3 adenosine receptors reduces bone-residing breast cancer in a rat preclinical model. Eur J Cancer. 2013;49:482–91.
Cohen S, Stemmer SM, Zozulya G, Ochaion A, Patoka R, Barer F, et al. CF102 an A3 adenosine receptor agonist mediates anti-tumor and anti-inflammatory effects in the liver. J Cell Physiol. 2011;226:2438–447.
Koscsó B, Csóka B, Pacher P, Haskó G. Investigational A3 adenosine receptor targeting agents. Expert Opin Investig Drugs. 2011. https://doi.org/10.1517/13543784.2011.573785.
Ledderose C, Hefti MM, Chen Y, Bao Y, Seier T, Li L et al. Adenosine arrests breast cancer cell motility by A3 receptor stimulation. Purinergic Signal. 2016. https://doi.org/10.1007/s11302-016-9531-6.
Aghaei M, Panjehpour M, Karami-Tehrani F, Salami S. Molecular mechanisms of A3 adenosine receptor-induced G1 cell cycle arrest and apoptosis in androgen-dependent and independent prostate cancer cell lines: involvement of intrinsic pathway. J Cancer Res Clin Oncol. 2011. https://doi.org/10.1007/s00432-011-1031-z.
Joshaghani H, Jafari S, Aghaei M, Panjehpour M, Abedi H. A3 adenosine receptor agonist induce G1 cell cycle arrest via Cyclin D and cyclin-dependent kinase 4 pathways in OVCAR-3 and Caov-4 cell lines. J Cancer Res Ther. 2017. https://doi.org/10.4103/0973-1482.199381.
Stemmer SM, Benjaminov O, Medalia G, Ciuraru NB, Silverman MH, Bar-Yehuda S et al. 1411 CF102 For the treatment of hepatocellular carcinoma: a phasE I/II, open-label, dose-escalation study. J Hepatol. 2012. https://doi.org/10.1016/s0168-8278(12)61422-6.
Mazzoni E, Pellegrinelli E, Mazziotta C, Lanzillotti C, Rotondo JC, Bononi I, et al. Mother-to-child transmission of oncogenic polyomaviruses BKPyV, JCPyV and SV40. J Infect. 2020;163:91–8.
Mazziotta C, Lanzillotti C, Torreggiani E, Oton-Gonzalez L, Iaquinta MR, Mazzoni E, et al. Serum antibodies against the oncogenic Merkel Cell Polyomavirus detected by an innovative immunological assay with mimotopes in healthy subjects. Front Immunol. 2021;12:676627.
Rotondo JC, Mazzoni E, Bononi I, Tognon M, Martini F. Association Between Simian Virus 40 and Human Tumors. Front Oncol. 2019;9:670.
Morello S, Petrella A, Festa M, Popolo A, Monaco M, Vuttariello E et al. Cl-IB-MECA inhibits human thyroid cancer cell proliferation independently of A3 adenosine receptor activation. Cancer Biol Ther. 2008. https://doi.org/10.4161/cbt.7.2.5301.
Lee HS, Chung HJ, Lee HW, Jeong LS, Lee SK. Suppression of inflammation response by a novel A 3 adenosine receptor agonist thio-Cl-IB-MECA through inhibition of Akt and NF-κB signaling. Immunobiology. 2011. https://doi.org/10.1016/j.imbio.2011.03.008.
Ahmad A, White CW, Ahmad S. Hypoxia-inducible factors and adenosine signaling in vascular growth. In: Extracellular ATP and Adenosine as Regulators of Endothelial Cell Function: Implications for Health and Disease. 2010. https://doi.org/10.1007/978-90-481-3435-9_7.
Acknowledgements
The works of JCR and MT have been supported by Associazione Italiana per la Ricerca sul Cancro (AIRC), grants MFAG 21956 and IG 21617, respectively. JCR was also supported by The International Association for the Study of Lung Cancer (IASLC).
Author information
Authors and Affiliations
Contributions
GC wrote the first draft; CM, JCR, CL collected the scientific literature, prepared the figures and wrote the paper; FM and MT corrected the paper and edited the final version. All authors approved the final text.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mazziotta, C., Rotondo, J.C., Lanzillotti, C. et al. Cancer biology and molecular genetics of A3 adenosine receptor. Oncogene 41, 301–308 (2022). https://doi.org/10.1038/s41388-021-02090-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41388-021-02090-z
This article is cited by
-
Cryo-EM structures of adenosine receptor A3AR bound to selective agonists
Nature Communications (2024)
-
Immortalized erythroid cells as a novel frontier for in vitro blood production: current approaches and potential clinical application
Stem Cell Research & Therapy (2023)
-
Hsa-microRNA-1249-3p/Homeobox A13 axis modulates the expression of β-catenin gene in human epithelial cells
Scientific Reports (2023)
-
Purinergic signaling during Marek’s disease in chickens
Scientific Reports (2023)
