
Abstract
Insulin-like growth factor 1 (IGF1) is a key factor for tissue growth and fuel metabolism. The potential function of central IGF1 remains unclear. We previously observed that IGF1 expression is increased in the hypothalamus of obese mice lacking STAT5 in the central nervous system (CNS). In this study, we explored the potential metabolic function of central IGF1 by intracerebroventricular (ICV) injection of IGF1, over-expression of central IGF1 by administering an adeno-associated virus (AAV), and ICV injection of an anti-IGF1 antibody. Mice that over-expressed central IGF1 displayed increased appetite, improved glucose tolerance and insulin sensitivity, decreased Pomc levels in the hypothalamus, and increased UCP1 expression in brown fat tissue. This is the first study demonstrating that central IGF1 regulates several important metabolic functions.
Similar content being viewed by others


Introduction
Insulin-like growth factors (IGFs) regulate a very large number of important physiological processes. Insulin-like growth factor 1 IGF1 regulates seed somatomedin C, is adin C, is a factors (IGFIgf1 gene that has a similartors (IGFs) regulate a very lar1. IGF1 has multiple biological effects, including promoting cell growth and proliferation2 and regulating fuel metabolism peripherally3. IGF1, which acts through IGF1 receptors and/or hybrid insulin/IGF1 receptors, has significant amino acid sequence homology with insulin, and enhances insulin sensitivity in both animal models and human subjects4. IGF1 is primarily secreted by the liver and mediates the endocrine actions of growth hormone (GH). GH, the main regulator of circulating IGF1 level in mammals, has an intricate regulatory relationship with IGF1. Mice lacking IGF1 in the liver have shown to display enhanced insulin sensitivity and glucose homeostasis5. A recent study has found that IGF1 is produced by all cell types in the brain6. IGF1 plays an important role in childhood growth and continues to havee brainrowth hormone (GH). GH, the main reF1 production can be retarded by under-nutrition, growth hormone insensitivity, lack of growth hormone receptors, or failure of downstream signaling pathways including SHP2 and STAT5B (signal transducer and activator of transcription 5B)7, 8. Low serum IGF1 levels are associated with reduced insulin sensitivity, metabolic syndrome, glucose intolerance, and the development of type 2 diabetes9.
The potential role of central IGF1 in regulating whole body energy homeostasis has not been investigated. To address this issue, we sought to study the effects of central IGF1 in mice. Mice that lack Stat5a/b in the central nervous system (Stat5NKO mice) develop severe obesity, accompanied by hyperphagia, hyperleptinemia, impaired thermal response to the cold, and insulin resistance10. We recently completed RNA-seq studies showing increased expression of members of the Igf family, including Igf1, Igf2, Igfals, and Igfbps, in the hypothalamus of Stat5NKO mice. Igf1 expression was very low in wild-type mice but elevated 8-fold in the Stat5NKO mice (Figure S1A). To determine the potential function of central IGF1 in energy metabolism, we subjected wild-type mice to ICV injections of IGF1 and an anti-IGF1 antibody. We found that IGF1 increased food intake while the anti-IGF1 antibody decreased appetite. We also over-expressed IGF1 long-term in the brain of wild-type mice by administering ICV injections of a recombinant adeno-associated virus 2 (AAV2). The phenotypes that we observed with the virus-treated mice were similar to those observed after acute ICV IGF1 injection.
Materials and methods
Generation of the IGF1 over-expression construct
The AAV2 construct encoding Igf1 (AAV-Igf1; Figure S2A) was generated by using a genomic 7.4 kb fragment containing all Igf1 exon sequences of the murine Igf1 gene. The flag–tag followed the IGF1-promoter fragment after the restriction enzyme was cut. AAV packaging was performed by the University of Pennsylvania Vector Core (Lot: V4738MI-S).
Animals and metabolic measurements
Male C57BL/6J mice (8–12 weeks old) were handled and housed in the Experimental Animal Center, located in building 14C at the National Institutes of Health, Bethesda, Maryland. This study was approved by the Institutional Animal Care and Use Committee of National Institute of Diabetes, Digestive and Kidney Diseases (NIDDK, K005-LBC-15), and all procedures were conducted in accordance with the guidelines of the National Institute of Health Animal Care and Use Committee. Mice were randomly (using a computerized, random number generator through the block-randomization method of Statistics Analysis System version 9.2.) divided into six groups: saline and IGF1 group, saline and anti-IGF1 group, AAV-empty and AAV-Igf1 injection group. Food intake (averaged per day), and body weight were measured daily after injection in each group. A total number of animals should be able to show a desired effect as few or many as possible (five animals in each group is required at least11). As our reaserch was a pilot study, so according to the reduced requirement (the number of animals used in experiment) from U.S. law and animal experimentation: 10 mice were included in each group for our experiment to avoid unnecessary suffer. In vivo metabolic tests were performed using standard procedures. To measure glucose tolerance (IGTT), mice were fasted overnight for 12 h, and blood was collected from the tail vein immediately before and 15, 30, 60, 90, and 120 min after i.p. injection of glucose (2g/kg) to determine blood glucose concentrations. To obtain a measure of peripheral insulin sensitivity (ITT), mice were fasted for 4 h, and blood glucose levels were measured before and at the indicated time points after i.p. injection of human insulin (0.75 U/kg). Blood glucose levels were determined using an automated blood glucose reader (Glucometer Elite Sensor; Bayer). Plasma insulin concentrations were determined by using an ELISA kit (Crystal Chem Inc. Cat.No.:#90080).
AAV-Igf1, IGF1 & anti-IGF1 injection
Pre-experiments were carried out for each injection before the formal study. Only those mice that recovered from the surgery could be allocated to experimental groups, others were euthanized according to the guidelines of the National Institute of Health Animal Care and Use Committee.
Mice were injected with 2 ul (1 ug/ml) of IGF1 (Peprotech, Catalog #: 100-11) or an anti-IGF1 antibody (Abcam, ab9572), or with 1011 GC/mouse of the AAV-Igf1 virus into the arcuate nucleus of the hypothalamus (ARC). Following injections, mice were individually housed, and body weight and food intake were determined as described above.
Mice were anesthetized with ketamine and xylazine (100–200 mg/kg and 5–10 mg/kg, respectively, given i.p.) and placed on a heating pad during anesthesia, surgery, and recovery. The mice were placed in a small-animal stereotaxic instrument (head held with ear bars and incisor holder). The surface of the skull were exposed with sterile surgical instruments then opened with a small burr drill over the appropriate stereotaxic coordinates. The dura was opened with a fine needle. A 30 gauge stainless steel needle attached to a 5-ul Hamilton syringe was lowered into the burr hole and 500 to 2 ul of the preparation was administered over 3–5 min per site (ARC). The animal was returned to its cage and monitored carefully twice daily for the first 48 h. Post-operative analgesia consisted of ketoprofen (5 mg/kg i.m. or s.c.) given every 24 h. A plastic cap was placed at the top of the cannula and removed for each injection after a 1-week recovery.
Brain immunofluorescence staining and liver histology
Twelve days after injection, the mice brains were collected and stored in 4% PFA overnight. Brains were sectioned at 40 um in thickness with Leica VT1000S. Sections were collected for the entire hypothalamus. Three sets of brain sections were stored at 4 °C in paraformaldehyde/phosphate-buffered saline (PBS). Flag antibodies: flag (F1804 sigma, 1:300, Secondary antibodies: Life Technology, A11017 Goat anti mouse 488; NeuN (MAB377, 1:1000); Secondary antibodies: Life technology, A11020, Goat anti Mouse 594; GFAP(C9205, 1:1000), Fluor conjugated 594 antibody, IGF1(abcam ab9572, 1:1000), secondary antibody Life technology, A11008, (1:1000)), Goat anti rabbit 488, and abcam Ab150064 (1:1000), donkey anti rabbit 594. Data analysis: fluorescent images were taken with NIDDK ALMIAC Keyence Digital Microscope and captured under exactly the same condition for both AAV-GFP (empty) control virus-injected and AAV-Igf1 virus-injected mice. Fiji free software (formerly image J, NIH) and Fiji program were used to run “MeanSigPerPixel_ROI-bg.ijm” Macro (For details, see word format text in 4th supplementary Material). Images were processed as original TIFF images after background deduction. Average total pixels were obtained for AAV-GFP (empty) control images and AAV-Igf1 images.
Mouse liver was harvested and fixed in 4% paraformaldehyde/phosphate-buffered saline for overnight and embedded in paraffin. For each liver, 5 um-thick sections were mounted on slides and stained with hematoxylin and eosin (HE) staining. Image acquisitions were performed using a BZ-II Viewer.
RNA and protein analyses
Total RNA was isolated using Trizol reagent (Cat. No.: 15596018) and QIAGEN RNase kit (Cat .No.: 74104). RNA concentrations were determined by a Qubit bio-analyzer (QUBIT2.0, Invitrogen, Q32866) prior to real-time PCR for gene expression. Western blotting was carried out with lysates prepared from isolated mouse liver or BAT by using standard techniques. Protein bands were quantitated using Image J software. The primers and antibodies used are listed below (See supplemental Tables 1 & 2).
Measurement of hepatic glycogen and triglyceride levels
Liver samples were homogenized in PBS, and glycogen and triglyceride contents were detected by using commercially available kits (Sigma; Cat. No.: MAK016 and Cat. No.: TR0100, respectively), strictly following the manufacturer’s instructions.
Statistical analysis
Statistical analyses were performed using SPSS 18.0. Multiple group comparisons were made with ANOVA, followed by the Tukey HSD test for multiple comparisons. Comparisons between two groups were performed using unpaired 2-tailed Student’s t test, and rank sum test was used for more than two time points. Data are shown as means ± SD, P < 0.05 was considered statistically significant.
Results
Hypothalamic IGF1 expression is increased in obese mice
We first examined IGF1 protein expression in the hypothalamus of the Stat5NKO mice. We found a higher IGF1 expression in Stat5NKO mice compared to their Stat5fl/fl lean control littermates (Figure S1B). To test the hypothesis that IGF1 expression is increased in other obese mouse-models, we also studied mice that had been maintained on a high-fat diet. In this model of diet-induced obesity (DIO), hypothalamic IGF1 expression was also significantly increased, compared to mice fed a standard chow diet (CD) (Figure S1C).
Over-expression efficiency of Igf1 in the arcuate nucleus of hypothalamus
After injecting recombinant AAV virus into the arcuate nucleus (ARC, 1011 genome copies/mouse) of wild-type mice, IGF1 over-expression in the ARC was confirmed via immunofluorescence (Fig. 1a-c). Both A and B are stained sections of the arcuate nucleus to show virus expression through GFP fluorescence and IGF1 antibody staining in AAV-GFP (empty) injected mice (Fig. 1a). We then applied flag tag antibody-staining to show AAV-Igf1 virus-expression after AAV-Igf1 injection into the mouse brain. Both IGF1 and GFAP (Glial marker) expressions were detected via immunostaining in AAV-Igf1-injected mice. The stained astrocytes showed only GFAP expression, and no IGF1 (Fig. 1b). Furthermore, both IGF1 and NeuN (neuron marker) expression in AAV-Igf1-injected mice were observed via immunostaining (Fig. 1c). These results showed that most IGF1-positive cells were NeuN-positive as well, indicating the expression of injected AAV-Igf1 in neurons. We also examined for IGF1 over-expression efficiency after AAV-GFP and AAV-Igf1 virus injection in the hypothalamus of the mice using immunostaining and western blot. Our experiment showed that IGF1 expression increased significantly in the hypothalamus of AAV-Igf1 mice than that in the AAV-GFP-injected mice. Total pixel-calculation that followed also verified that IGF1-expression level was higher in the AAV-Igf1group than in the AAV-GFP group (Figs. 2a, b, Figure S2B). These results indicated that AAV-Igf1 injected mice had increased IGF1 expression in the brain, indicating a successful IGF1 over-expression model.

a Representative immunofluorescence staining of GFP fluorescence and IGF1 to show viral expression in the hypothalamus of AAV-GFP (empty) injected mice. b Representative immunofluorescence staining to detect IGF1 expression and GFAP (Glial marker) expression in the hypothalamus of AAV-Igf1 injected mice. c Representative immunofluorescence staining to determine IGF1 and NeuN (neuron marker) expression in the hypothalamus of AAV-Igf1 injected mice. Scale bar indicates 100 um, n = 10

a IGF1 antibody staining and total-pixel calculation in both AAV-GFP- and AAV-IGF1-injected mouse brain sections (n = 10, scale bar = 100 um). b IGF1 antibody staining and total-pixel calculation in the hypothalamus of AAV-GFP- or AAV-Igf1-injected mice (n = 10, scale bar = 100 um). Data are shown as means ± SD, *P < 0.01 vs. AAV-GFP group
Central IGF1 increases food intake
To explore the potential function of central IGF1, we injected IGF1 or an anti-IGF1 antibody into the lateral cerebral ventricle (ICV) of wild-type mice. The IGF1-injected mice displayed a significant increase in food intake (Fig. 3a), where as the mice injected with the anti-IGF1 antibody showed reduced food intake (Fig. 3b). Neither treatment affected body weight during the 48-hour observation period (Figs. 3c, d). To study the effects of long-term IGF1 over-expression in the brain, we injected the wild-type mice with a recombinant AAV coding for IGF1. Specifically, the virus was injected into the ARC, which plays a key role in the central regulation of food intake and energy homeostasis. We found that the virus-treated mice displayed enhanced appetite but unchanged body weight (Figs. 3e, f), mimicking the effects of acute central IGF1 administration.

IGF1 or an anti-IGF1 antibody were injected ICV (2 ul, 1 ug/ml). AAV-Igf1 was injected into the ARC (1011 GC/mouse) of wild-type mice. a & b Food consumption 48 h after IGF1 and anti-IGF1 injection, n = 10 per group. c & d Body weight 48 h after IGF1 and anti-IGF1 injection, n = 10 per group. (e & f) Food consumption and body weight 2 weeks after virus injection, n = 12 per group. Data are expressed as means ± SD. *P < 0.05, **P < 0.01 vs. saline or AAV-GFP group
Central IGF1 improves glucose homeostasis and insulin sensitivity
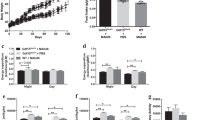
We then measured serum insulin and blood glucose levels in mice fasted for 4 or 16 (o/n) hours. We found that acute central IGF1 treatment (Figs. 4a, b) or prolonged IGF1 over-expression in the ARC (Figs. 4c, d) significantly increased serum insulin and reduced blood glucose levels. Glucose and insulin tolerance tests showed that both treatments led to improved glucose tolerance (Figs. 4e, f) and enhanced insulin sensitivity (Figs. 4g, h). In contrast, ICV injection of the anti-IGF1 antibody caused decreased serum insulin and elevated blood glucose levels (Figs. 4i, j). Somewhat surprisingly, these mice displayed normal glucose tolerance and insulin sensitivity (Figs. 4k, l).

. Glucose and insulin responses were measured after ICV injection of IGF1 or anti-IGF1, or ARC injection of AAV-Igf1 injection. a & b Free-fed and fasting blood glucose and serum insulin levels after IGF1 injection. (c & d) Free-fed and fasting glucose and serum insulin levels after AAV-Igf1 virus injection. e & f Glucose tolerance tests (GTT) after IGF1 or AAV-Igf1 injection. Glucose (2 g/kg) was administered i.p after a 14 h fast (mouse age: 13 weeks). g & h Insulin tolerance tests (ITT) after IGF1 or AAV-Igf1 injection. Insulin (0.75 U/kg) was administered i.p. after a 4 h fast (mouse age: 12 weeks). i & j Free-fed and fasting (4 hr fast) blood glucose and serum insulin level after anti-IGF1 injection. k & l GTT and ITT after anti-IGF1 injection. (m) Immunoblots of insulin receptor β subunit and Akt in liver after AAV-Igf1 virus injection. n Ratio of p-Akt, p-IR β in liver of AAV-Igf1 mice relative to AAV-GFP mice (quantitative analysis was normalized by total IR or total Akt). o Immunoblots of UCP1 expression in brown adipose tissue (BAT) after AAV-Igf1 virus injection. p Ratio of UCP1 expression in BAT of AAV-Igf1 mice relative to AAV-GFP mice. Data are repeated twice and expressed as means ± SD. *P < 0.05, **P < 0.01 vs. saline or AAV-GFP group, n = 10
To explore the mechanism by which central IGF1 improved glucose homeostasis and insulin sensitivity, we examined the levels of Akt and the insulin receptor beta subunit (ISR-beta) in the liver, which are predicted to play a key role in IGF1 signaling12,13,14. Akt activation is associated with its auto-phosphorylation at Thr-47315. Western blotting studies demonstrated that the abundance of total ISR-beta and Akt did not differ between control and AAV-Igf1 mice, whereas the expressions of the phosphorylated (active) forms of Akt and ISR-beta were significantly increased in the AAV-Igf1 mice (Figs. 4m, n). These results provide further evidence that enhanced central IGF1 is critical for insulin responsiveness through activation of Akt and ISR-beta.
The presence of UCP1 in brown adipose tissue (BAT) directs its oxidative metabolism almost entirely to thermogenesis16, 17. UCP1/BAT activity is a well-known regulator of energy homeostasis. Western blotting studies showed that UCP1 expression in BAT from AAV-Igf1 mice was significantly increased, compared to control mice (mice injected with empty AAV) (Fig. 4o, p). Strong neuroanatomical and functional evidence has shown that WAT and BAT are innervated and regulated by the sympathetic nervous system (SNS) for thermoregulation and beige adipocyte formation18. Since UCP1 is the hallmark protein responsible for cold- and diet-induced thermogenesis in BAT19, its expression can also be regulated by SNS activity. These data suggest that central IGF1 action enhances central sympathetic outflow.
Central IGF1 reduces liver weight and lowers triglyceride and glycogen levels
We also examined the effects of long-term central IGF1 over-expression on liver weight and triglyceride and glycogen levels, and found that liver weight and hepatic triglyceride and glycogen levels were significantly reduced in AAV-Igf1 mice, compared to control mice (Figs. 5a, c). Liver morphology did not differ between AAV-Igf1 and control mice (Fig. 5b). These results suggest that central IGF1 signaling promotes glucose and lipid metabolism in the liver. This observation is consistent with the concept that central IGF1 enhances stimulates sympathetic outflow (see previous paragraph).

The AAV-Igf1 virus or AAV- GFP was injected into the ARC of wild-type mice. a Liver tissue was collected from 14-week-old male mice for weight measurements. b H&E staining of liver sections; scale bar, 100 um. c Liver triglyceride and glycogen content. Data are repeated twice and expressed as means ± SD. *P < 0.05, **P < 0.01 vs. AAV-GFP group, n = 10
Effects of IGF1 on gene expression of Pomc, Npy, Agrp in the hypothalamus
The ARC is a key hypothalamic nucleus in the regulation of appetite20, 21. Although the ARC contains other cell types, two main opposing types of output neurons have been defined: those expressing pro-opiomelanocortin (Pomc; Pomc neurons) and those containing Agouti-related protein (AgRP) and neuropeptide-Y (NPY) (AgRP neurons). The Pomc neurons release α-melanocyte stimulating factor (α-MSH), which collectively promotes activity and energy expenditure, and suppresses food intake. AgRP and NPY are neuropeptides that suppress energy use and promote food intake22, 23. To study whether central IGF1 modulates the expression of these appetite-regulating peptides, we used qRT-PCR to measure their expression levels after central administration of IGF1 or an anti-IGF1 antibody, or after long-term IGF1 expression in AAV-Igf1 mice. Acute IGF1 injection and long-term over-expression of IGF1 in the mouse brain did not affect Agrp and Npy expression, but significantly reduced Pomc expression (Figs. 6a, b). In contrast, central administration of the anti-IGF1 antibody increased Pomc expression, but did not change Agrp and Npy expression. It is therefore likely that the orexigenic effect observed after central IGF1 administration or over-expression is caused, at least partially, by a reduced Pomc expression.

. mRNA levels were determined via qRT-PCR. a Pomc, Npy, and Agrp expression after IGF1 or anti-IGF1 injection (ICV). b Pomc, Npy, and Agrp expression after AAV-Igf1 injection (into the ARC). Data are repeated twice and expressed as means ± SD. **P < 0.01 vs. saline or AAV- GFP group, n = 5
Discussion
The central nervous system plays an important role in the development of obesity24, 25. Loss of cytokine-STAT5 signaling in the CNS and pituitary gland alters energy balance and leads to obesity in mice10. We recently identified a large number of genes with altered expression levels in the hypothalamus of Stat5NKO mice (unpublished data). Surprisingly, in the hypothalamus of Stat5NKO mice, most members of the Igf1 family were found to be up-regulated. STAT5 has been shown to positively regulate IGF1 function in peripheral organs26, such as skeletal muscle and liver27. Some studies showed that modulating IGF1 has a dramatic effect on glucose metabolism in mice under both normal and high-fat diet feeding. For example, whole body over-expression of IGFBP1 effectively reduced the hypoglycemic effect of endogenous IGF1 and caused elevated plasma insulin levels, yet reduced insulin-stimulated glucose transport and glycogen synthesis in skeletal muscle, indicative of mild insulin resistance28. Pituitary-derived GH exerts its growth effects primarily by regulating the expression of IGF-I in both hepatic and non-hepatic tissues29. GH’s action is initiated upon binding to the cell-surface GH receptor (GHR), a homodimeric transmembrane protein. The GH-GHR interaction induces signal transduction through recruitment and activation of the cytosolic Janus kinase 2 (JAK2)30. The activated complex of signaling pathways includes four STAT pathways (STAT1, 3, 5a, and 5b), the MAPK (mitogen-activated protein kinase), and the PI3K (phosphoinositide-3 kinase) pathways. Activation of these pathways culminates in the regulation of multiple genes, including IGF1, IGFBP3, and IGFALS (acid labile subunit, ALS). One study has reported that brain-specific over-expression of IGFBP-6 under the control of glial fibrillary acidic protein promoter causes glucose intolerance, insulin resistance, and elevated weight gain both on a normal diet and on a high-fat diet31. Reduction of IGF1 signaling by inactivating the IGF1R gene in mice can also lead to impaired glucose homeostasis32, 33. However, these previous studies did not explore the potential role of IGF1 in the hypothalamus of the mouse brain. Given that Stat5 is a positive regulator of Igf1, Stat5 deficiency is expected to reduce Igf1 expression. However, we observed the opposite effect—an increase of Igf1 gene expression in the hypothalamus of Stat5NKO mice. As the central role of IGF1 remains controversial34,35,36, we performed the current study to explore the potential roles of central IGF1 in a more direct and systematic fashion.
In this study, we describe the metabolic phenotypes of IGF1- and IGF1 antibody-treated mice. Gain of function studies (ICV injection of IGF1 or AAV-mediated over-expression of IGF1) as well as loss of function experiments (ICV injection of an anti-IGF1 antibody) demonstrated that central IGF1 promoted food intake and insulin secretion, reduced blood glucose levels, and led to improved glucose tolerance (Fig. 4). Moreover, over-expression of IGF1 in the brain led to enhanced phosphorylation/activation of the insulin receptor (IRb subunit) and AKT, a key mediator of signaling pathways stimulated by insulin-like peptides36 (Fig. 4m). In addition, an increase in central IGF1 expression caused an increase of UCP1 expression in BAT (Fig. 4o), suggesting that central IGF1 can promote BAT activity and enhance energy expenditure. Since BAT activity is primarily under the control of the sympathetic nervous system, central IGF1 activity likely leads to an increase in central sympathetic outflow. Consistent with this notion, we also observed reduced hepatic triglyceride and glycogen levels (Fig. 5a-c). To the best of our knowledge, this is the first study that directly examined the role of central IGF1 in regulating lipid and glucose metabolism.
The regulation of energy homeostasis is closely controlled by the central nervous system and integrated signals from the periphery, includes insulin signaling37. Insulin acts on ARC neurons to inhibit the expression of the orexigenic neuropeptides NPY and AgRP and to enhance the expression of Pomc, an appetite-suppressing peptide, resulting in reduced food intake38,39,40,41. In contrast, we observed in the present study that over-expression of IGF1 in the ARC promoted food consumption, most likely due to an increase in Pomc expression. The receptors and neuronal pathways involved in this phenotype will be explored in the future studies. Meanwhile, the autonomic nervous system also plays a key role in the response to these signals, innervating peripheral metabolic tissues, including BAT, WAT, liver, and skeletal muscle42. Taking together, these new data shed light onto the role of central IGF1. In particular, the reported metabolic effects in BAT and in liver, caused by AAV-induced expression in brain of Igf1, are attributed mainly to a direct effect of IGF1 on neurons, and then mediated by the sympathetic nervous system. This is however highly speculative, and more experiments are required to justify this hypothesis.
Nevertheless, some limitations should be acknowledged in the interpretation of our results. One is that we only used single IGF1 and anti-IGF1 antibody doses. In addition, the detailed neuronal/cellular pathways through which central IGF1 exerts its metabolic actions remain to be explored. Also, the potential ICV IGF1 actions on other (non-hypothalamic) brain regions regulating food-intake need to be studied further. IGF2, which has similar homology with IGF1, is primarily secreted by the liver and in utero by the placenta. Igf2 has been reported to play an important role in the subventricular zone and in the subgranular zone of the hippocampus for adult-neurogenesis in mice43. Moreover, over-expression of Igf2 is associated with the intrauterine programming of adipose tissue and fetal overgrowth44. Current data about IGF2 and obesity is both limited and conflicting, so the potential role of central IGF2 in regulating energy and glucose homeostasis also need to be further elucidated. Nevertheless, our findings may guide future efforts to develop novel classes of drugs useful for the treatment of obesity and diabetes.
Conclusion
In the present study, we systematically explored the role of central IGF1. We found that central IGF1 can promote feeding, improve insulin sensitivity and glucose tolerance, and stimulate energy expenditure via thermogenesis.
Change history
19 February 2021
A Correction to this paper has been published: https://doi.org/10.1038/s41387-021-00153-4
References
Hoppener, J. W. et al. The human gene encoding insulin-like growth factor I is located on chromosome 12. Hum. Genet. 69, 157–160 (1985).
Jones, J. I. & Clemmons, D. R. Insulin-like growth factors and their binding proteins: biological actions. Endocr. Rev. 16, 3–34 (1995).
White, M. F. IRS2 integrates insulin/IGF1 signalling with metabolism, neurodegeneration and longevity. Diabetes. Obes. Metab. 16(Suppl 1), 4–15 (2014).
Yakar, S. et al. Liver-specific igf-1 gene deletion leads to muscle insulin insensitivity. Diabetes. 50, 1110–1118 (2001).
Yakar, S. et al. Inhibition of growth hormone action improves insulin sensitivity in liver IGF-1-deficient mice. J. Clin. Invest. 113, 96–105 (2004).
Bach, M. A., Shen-Orr, Z., Lowe, W. L. Jr, Roberts, C. T. Jr & LeRoith, D. Insulin-like growth factor I mRNA levels are developmentally regulated in specific regions of the rat brain. Brain research. Mol Brain Res 10, 43–48 (1991).
Baik M et al. Muscle-specific deletion of signal transducer and activator of transcription 5 augments lipid accumulation in skeletal muscle and liver of mice in response to high-fat diet. Eur J Nutr https://doi.org/10.1007/s00394-015-1101-0 (2015).
Baik, M. et al. Liver-specific deletion of the signal transducer and activator of transcription 5 gene aggravates fatty liver in response to a high-fat diet in mice. J. Nutr. Biochem. 29, 56–63 (2016).
Succurro, E. et al. Low plasma insulin-like growth factor-1 levels are associated with reduced insulin sensitivity and increased insulin secretion in nondiabetic subjects. Nutr Metab Cardiovasc Dis 19, 713–717 (2009). 19.
Lee, J. Y. et al. Loss of cytokine-STAT5 signaling in the CNS and pituitary gland alters energy balance and leads to obesity. PLoS. ONE. 3, e1639 (2008).
Benjamin, M. & Rainer, M. Chances and obstacles of pilot projects in animal research for statistical sample size. Int J Biol Med Res. 2012, 2319–2323 (2012). 3.
Lu, M. et al. Insulin regulates liver metabolism in vivo in the absence of hepatic Akt and Foxo1. Nat. Med. 18, 388–395 (2012).
Mackenzie, R. W. & Elliott, B. T. Akt/PKB activation and insulin signaling: a novel insulin signaling pathway in the treatment of type 2 diabetes. Diabetes Metab Syndr Obes 7, 55–64 (2014).
Vander Haar, E., Lee, S. I., Bandhakavi, S., Griffin, T. J. & Kim, D. H. Insulin signalling to mTOR mediated by the Akt/PKB substrate PRAS40. Nat. Cell. Biol. 9, 316–323 (2007).
Dai, N. et al. IGF2BP2/IMP2-Deficient mice resist obesity through enhanced translation of Ucp1 mRNA and Other mRNAs encoding mitochondrial proteins. Cell. Metab. 21, 609–621 (2015).
Nedergaard, J. et al. UCP1: the only protein able to mediate adaptive non-shivering thermogenesis and metabolic inefficiency. Biochim. Biophys. Acta. 1504, 82–106 (2001).
Kwon, M. M., O'Dwyer, S. M., Baker, R. K., Covey, S. D. & Kieffer, T. J. FGF21-Mediated Improvements in Glucose Clearance Require Uncoupling Protein 1. Cell Rep 13, 1521–1527 (2015).
Nguyen, N. L. et al. Separate and shared sympathetic outflow to white and brown fat coordinately regulates thermoregulation and beige adipocyte recruitment. Am. J. Physiol. Regul. Integr. Comp. Physiol. 312, R132–R145 (2017).
Bonet, M. L., Mercader, J. & Palou, A. A nutritional perspective on UCP1-dependent thermogenesis. Biochimie. 134, 99–117 (2017).
Fei, W., Tian de, R., Tso, P. & Han, J. S. Arcuate nucleus of hypothalamus is involved in mediating the satiety effect of electroacupuncture in obese rats. Peptides. 32, 2394–2399 (2011).
Stanley, S., Wynne, K., McGowan, B. & Bloom, S. Hormonal regulation of food intake. Physiol. Rev. 85, 1131–1158 (2005).
Hill, J. W. et al. Direct insulin and leptin action on pro-opiomelanocortin neurons is required for normal glucose homeostasis and fertility. Cell. Metab. 11, 286–297 (2010).
Lin, H. V. et al. Divergent regulation of energy expenditure and hepatic glucose production by insulin receptor in agouti-related protein and POMC neurons. Diabetes. 59, 337–346 (2010).
Coll, A. P., Farooqi, I. S., Challis, B. G., Yeo, G. S. & O'Rahilly, S. Proopiomelanocortin and energy balance: insights from human and murine genetics. J. Clin. Endocrinol. Metab. 89, 2557–2562 (2004).
Herrera, B. M., Keildson, S. & Lindgren, C. M. Genetics and epigenetics of obesity. Maturitas. 69, 41–49 (2011).
Li, Q. et al. NO-1886 suppresses diet-induced insulin resistance and cholesterol accumulation through STAT5-dependent upregulation of IGF1 and CYP7A1. J. Endocrinol. 204, 47–56 (2010).
Klover, P., Chen, W., Zhu, B. M. & Hennighausen, L. Skeletal muscle growth and fiber composition in mice are regulated through the transcription factors STAT5a/b: linking growth hormone to the androgen receptor. FASEB J 23, 3140–3148 (2009).
Rajkumar, K., Krsek, M., Dheen, S. T. & Murphy, L. J. Impaired glucose homeostasis in insulin-like growth factor binding protein-1 transgenic mice. J. Clin. Invest. 98, 1818–1825 (1996).
Martinez, C. S. et al. Growth hormone STAT5-mediated signaling and its modulation in mice liver during the growth period. Growth Horm IGF Res 23, 19–28 (2013).
Kofoed, E. M. et al. Growth hormone insensitivity associated with a STAT5b mutation. N. Engl. J. Med. 349, 1139–1147 (2003).
Bienvenu, G. et al. Dysregulation of energy homeostasis in mice overexpressing insulin-like growth factor-binding protein 6 in the brain. Diabetologia. 48, 1189–1197 (2005).
Garg, N., Thakur, S., McMahan, C. A. & Adamo, M. L. High fat diet induced insulin resistance and glucose intolerance are gender-specific in IGF-1R heterozygous mice. Biochem. Biophys. Res. Commun. 413, 476–480 (2011).
Holzenberger, M. et al. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature. 421, 182–187 (2003).
Wit, J. M. & Walenkamp, M. J. Role of insulin-like growth factors in growth, development and feeding. World. Rev. Nutr. Diet. 106, 60–65 (2013).
Werner, H. & LeRoith, D. Insulin and insulin-like growth factor receptors in the brain: physiological and pathological aspects. Eur Neuropsychopharmacol 24, 1947–1953 (2014).
Fernandez, A. M. & Torres-Aleman, I. The many faces of insulin-like peptide signalling in the brain. Nat. Rev. Neurosci. 13, 225–239 (2012).
Marino, J. S., Xu, Y. & Hill, J. W. Central insulin and leptin-mediated autonomic control of glucose homeostasis. Trends. Endocrinol. Metab. 22, 275–285 (2011).
Myers, M. G. Jr & Olson, D. P. Central nervous system control of metabolism. Nature. 491, 357–363 (2012).
Myers, M. G. Jr & Olson, D. P. SnapShot: neural pathways that control feeding. Cell. Metab. 19, 732–732 (2014). e731.
Belgardt, B. F., Okamura, T. & Bruning, J. C. Hormone and glucose signalling in POMC and AgRP neurons. J. Physiol. 587, 5305–5314 (2009).
Varela, L. & Horvath, T. L. Leptin and insulin pathways in POMC and AgRP neurons that modulate energy balance and glucose homeostasis. EMBO. Rep. 13, 1079–1086 (2012).
Seoane-Collazo, P. et al. Hypothalamic-autonomic control of energy homeostasis. Endocrine. 50, 276–291 (2015).
Ferron, S. R. et al. Differential genomic imprinting regulates paracrine and autocrine roles of IGF2 in mouse adult neurogenesis. Nature Commun 6, 8265 (2015).
Kadakia, R. & Josefson, J. The Relationship of Insulin-Like Growth Factor 2 to Fetal Growth and Adiposity. Horm Res Paediatr 85, 75–82 (2016).
Acknowledgements
This study was supported by the National Natural Science Foundation of China (Grant No. 81574063, 81273838), and the Intramural Research Program (IRP) of NIDDK. We thank Dr. Jürgen Wess (NIH, NIDDK) for advice during the course of this study and for helpful comments on the manuscript; Ms. Wan-Xin Liu (University of Pennsylvania School of Veterinary Medicine) for the editing of English; Ms. Yi Yang (Chengdu University of Chinese Medicine) for statistical analysis and Dr. Lothar Hennighausen (NIH, NIDDK) for providing Stat5 floxed mice.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Authors’ contributions
HH did experiments, analyzed data and wrote the manuscript; ZZC allocated experimental groups, performed the IGF1, anti-IGF1 and virus injections, and IF staining analysis; LZ helped generate the AAV construct; SPF did data analysis; YHC and MR helped with the physiological studies; BMZ guided the experiments, wrote and revised the manuscript.
Competing interests
The authors declare that they have no competing financial interests.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hong, H., Cui, ZZ., Zhu, L. et al. Central IGF1 improves glucose tolerance and insulin sensitivity in mice. Nutr & Diabetes 7, 2 (2017). https://doi.org/10.1038/s41387-017-0002-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41387-017-0002-0
This article is cited by
-
Imprinted Dlk1-Gtl2 cluster miRNAs are potential epigenetic regulators of lamb fur quality
BMC Genomics (2023)
-
The metabolic effects of resumption of a high fat diet after weight loss are sex dependent in mice
Scientific Reports (2023)
-
Testicular ultrastructure and hormonal changes following administration of tenofovir disoproxil fumarate-loaded silver nanoparticle in type-2 diabetic rats
Scientific Reports (2022)
-
Neuroendocrine microRNAs linked to energy homeostasis: future therapeutic potential
Pharmacological Reports (2022)
-
Adeno‐associated virus (AAV)-based gene therapy for glioblastoma
Cancer Cell International (2021)
