
Abstract
Identifying mechanisms underlying the intergenerational transmission of risk for attention-deficit/hyperactivity disorder (ADHD) traits can inform interventions and provide insights into the role of parents in shaping their children’s outcomes. We investigated whether genetic transmission and genetic nurture (environmentally mediated effects) underlie associations between polygenic scores indexing parental risk and protective factors and their offspring’s ADHD traits. This birth cohort study included 19,506 genotyped mother-father-offspring trios from the Norwegian Mother, Father and Child Cohort Study. Polygenic scores were calculated for parental factors previously associated with ADHD, including psychopathology, substance use, neuroticism, educational attainment, and cognitive performance. Mothers reported on their 8-year-old children’s ADHD traits (n = 9,454 children) using the Parent/Teacher Rating Scale for Disruptive Behaviour Disorders. We found that associations between ADHD maternal and paternal polygenic scores and child ADHD traits decreased significantly when adjusting for the child polygenic score (pΔβ = 9.95 × 10−17 for maternal and pΔβ = 1.48 × 10−14 for paternal estimates), suggesting genetic transmission of ADHD risk. Similar patterns suggesting genetic transmission of risk were observed for smoking, educational attainment, and cognition. The maternal polygenic score for neuroticism remained associated with children’s ADHD ratings even after adjusting for the child polygenic score, indicating genetic nurture. There was no robust evidence of genetic nurture for other parental factors. Our findings indicate that the intergenerational transmission of risk for ADHD traits is largely explained by the transmission of genetic variants from parents to offspring rather than by genetic nurture. Observational associations between parental factors and childhood ADHD outcomes should not be interpreted as evidence for predominantly environmentally mediated effects.
Similar content being viewed by others

Introduction
Intergenerational psychiatry aims to understand how parents contribute to the emergence of psychiatric risk in their offspring [1]. A fundamental question is whether parents affect their offspring via genetic or environmental pathways, or both. Different mechanisms have different implications for intervention strategies aiming to disrupt the cycle of transmission across generations. Here, we implement a novel design capitalising on genomic data from parents and their children to systematically investigate the intergenerational transmission of inattention and hyperactivity-impulsivity traits comprising attention-deficit/hyperactivity disorder (ADHD) [2].
Observational studies have identified many parental factors associated with offspring ADHD: parental psychiatric conditions including ADHD itself, depression, anxiety disorders, schizophrenia, bipolar disorder and autism spectrum disorder (autism) [3,4,5,6,7,8,9,10]; personality traits like neuroticism [11]; cognitive disability [9], lower education and socioeconomic position [12,13,14,15]; and substance use including maternal smoking, alcohol consumption and cannabis use during pregnancy [16,17,18,19,20]. Like most complex phenotypes, these parental factors as well as offspring ADHD are partially heritable [21,22,23], which leaves findings from such studies vulnerable to confounding by shared genetic factors. Genetic variants transmitted from parents to children can independently affect both parental factors and their offspring’s outcomes. This can lead to associations between parental factors and ADHD traits in the absence of environmental pathways of causation [24]. Addressing this issue is fundamental to assess whether modifiable parental factors are likely to be effective targets for intervention.
Several genetically informed designs for causal inference [24, 25] such as family designs, have demonstrated that genetic transmission from parents to their children need to be accounted for in aetiological studies of childhood ADHD [10, 15, 26,27,28]. However, these studies require specific samples (e.g., twins or adopted children and their parents), leading to both scarcity and paucity of suitable samples. Furthermore, most studies focus on one or a few parental factors at a time. Each design also has its own limitations. For example, in the adoption design, the association between ADHD traits in the adoptive mother and child may be partly due to reverse causation. As such, triangulation of findings between studies that employ genetically informed designs with different underlying assumptions and limitations is essential [29, 30].
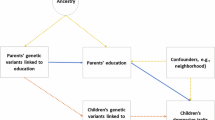
Intergenerational transmission can be investigated in population-based samples with genotype data on parents and offspring and phenotypic measures on offspring outcomes [31, 32]. Instead of phenotypic parental measures, genetic variants associated with the parental factor (e.g., ADHD) can be combined into a polygenic score which reflects an individual’s genetic propensity. As illustrated in Fig. 1, relevant estimates can be obtained by regressing children’s outcomes jointly onto their own and both their parents’ polygenic scores. This multivariate trio model provides estimates of: (i) the direct genetic effects on the offspring’s outcome due to parent-to-offspring allele transmission (path c), and (ii) the indirect genetic effects of parental alleles not due to influences of alleles inherited by the offspring (paths m and f). In this trio design, the associations between parental polygenic scores and offspring traits are adjusted for the offspring polygenic score to account for the confounding induced by the transmission of genetic variants from parents to offspring. Such adjusted associations are called indirect genetic effects, which originate in parental genomes but affect their offspring via environmental pathways (i.e., independently of genetic transmission). In the case of ADHD, parental ADHD variants influence parental ADHD traits, which in turn may shape the child rearing environment (i.e., nurture), ultimately affecting their offspring’s ADHD traits. This indirect parental genetic effect is henceforth referred to as “genetic nurture” [32]. Importantly, all genetic variants comprising parental genomes can contribute to the genetic nurture effect, for example by influencing parenting behaviours, whether or not variants are transmitted to the offspring. In contrast, only transmitted parental alleles can be involved in direct genetic effects in the offspring generation. In the trio design, direct genetic effects are estimated from the association between children’s polygenic scores and their ADHD traits whilst adjusting for the maternal and paternal polygenic scores. In this way, confounding by genetic nurture effects is removed. This approach is approximately analogous to computing distinct polygenic scores based on transmitted and non-transmitted alleles, and using these to derive estimates of genetic nurture and direct genetic effects [32]. These two approaches have been shown to produce similar results [33].

Path diagram illustrating a regression model with polygenic scores for mother (Gmother), father (Gfather) and child (Gchild) predicting child ADHD traits. Genetic influences shared between the child and their mother (path Tm) and father (path Tf) are illustrated in purple. Coefficients for path c represent direct genetic effects of the child’s polygenic score on their ADHD traits after controlling for the influences of parental polygenic scores. Genetic transmission is the path from parental genetics to child traits via child genetics (e.g., Gmother to child ADHD via paths Tm and c). Attenuation of associations between parental polygenic scores and child ADHD when adjusting for child polygenic scores is evidence for genetic transmission. Genetic nurture effects (highlighted in green) on childhood ADHD are estimated from path coefficients m for mothers and f for fathers. Coefficients for genetic nurture paths m and f reflects the effects of parental genotypes on children’s traits after statistically controlling for the genetic influences transmitted from parents to their child and for covariation between parental polygenic scores (path a). Additional influences captured by genetic nurture effect estimates can also include familial environments such as sibling or grandparent effects, and estimates may be influenced by demographic factors such as population stratification and assortative mating. Note that path a will be near zero in the absence of assortative mating, which was the case for correlations between most parental polygenic scores used in the analyses, apart from educational attainment (r = 0.11; Supplementary Table 3).
To date, only one study, in the Netherlands Twin Register cohort, has used polygenic scores to investigate the association between parental factors and their offspring’s ADHD traits [34]. They found that the association between parental factors and their offspring’s ADHD traits could mainly be attributed to genetic transmission rather than to genetic nurture. However, the Netherlands Twin Register study focused on only two parental factors (ADHD and education) and had a relatively small sample (N = 2518 families).
In the present study, we use a large population-based cohort of genotyped mother-father-offspring trios to systematically investigate a range of putative parental risk factors for offspring ADHD, including psychiatric conditions, substance use, neuroticism, educational attainment (EA) and cognitive performance. We jointly model maternal, paternal and offspring polygenic scores to quantify genetic versus environmental routes of risk transmission.
Samples and methods
Participants and genotype data
The Norwegian Mother, Father and Child Cohort Study (MoBa) [35, 36] is a population-based pregnancy cohort study conducted by the Norwegian Institute of Public Health. Participants were recruited from across Norway during 1999–2008. Mothers consented to participation in 41% of the pregnancies. The cohort includes 114,500 children, 95,200 mothers and 75,200 fathers. Blood samples were obtained from both parents during pregnancy and from mothers and children (umbilical cord) at birth. The Medical Birth Registry (MBRN) is a national health registry containing information about all births in Norway and was used to obtain children’s year of birth and sex. The current study is based on version 12 of the quality-assured data files released for research in January 2019. The establishment of MoBa and initial data collection was based on a license from the Norwegian Data Protection Agency and approval from The Regional Committees for Medical and Health Research Ethics (REK). MoBa is based on regulations based on the Norwegian Health Registry Act. The current study was approved by REK (2016/1702).
The eligible sample was MoBa families with genotypic data [37] available for mother-father-offspring trios. After quality control (procedures are outlined in the Supplementary Note), we excluded up to second-degree relatives within generations from within groups of related family trios (leaving no families with e.g., siblings in the parental or offspring generation) whilst prioritising trios with complete phenotypic data, resulting in 19,506 trios with genotypic data available (Supplementary Fig. 1).
Measures
Childhood traits of attention-deficit hyperactivity disorder
Children’s ADHD traits were reported by mothers using the Parent/Teacher Rating Scale for Disruptive Behaviour Disorders (RS-DBD) [38] that contains 18 items related to DSM-IV criteria for ADHD. Items were rated on a four-point scale. Participants with fewer than half completed items were excluded, which resulted in 9,454 families with phenotypic data available. We computed a total ADHD summary score and repeated analyses with inattention and hyperactivity-impulsivity subscales (9 items each). To mitigate bias due to attrition in cohort studies, multiple imputation was performed to derive ADHD scores for the full study sample (N = 19,506 genotyped family trios) imputed from polygenic scores (which were available for all eligible participants and, thus, not imputed) and 23 auxiliary variables previously found to predict ADHD in MoBa [39], including records of ADHD diagnoses and earlier measures of ADHD traits, socioeconomic variables and child motor and language development (see Supplementary Note for additional information). Descriptive statistics on auxiliary variables and evaluation of imputation quality are provided in Supplementary Table 1 and Supplementary Figs. 2-3.
Polygenic scores indexing parental factors
Polygenic scores were used as indicators of genetic liabilities for the corresponding parental factors (e.g., the maternal polygenic score for depression). Polygenic scores are individual-level scores that summarize common variant genetic risk for a given phenotype. The scores are calculated as the weighted sum of effect alleles for all measured single nucleotide polymorphisms (SNPs) in an individual. SNP weights are obtained from summary statistics from Genome-Wide Associations Studies (GWAS; e.g. for depression).
We derived polygenic scores corresponding to 12 putative parental risk factors identified based on previous literature [40,41,42,43,44,45,46,47,48,49,50,51] and the availability of well-powered GWAS of relevant phenotypes (Supplementary Table 2). One polygenic score per parental factor was generated for each mother, father and child using PRS-PC [52]. PRS-PC takes the first principal component of polygenic scores computed at several p-value thresholds, which increases the predictive accuracy of polygenic scores while avoiding overfitting (see Supplementary Note).
Correlations between parental polygenic scores are provided in Supplementary Table 3. Compared to family trios with complete data, families with missing offspring ADHD scores had higher polygenic scores for ADHD, schizophrenia, depression, neuroticism and smoking, and lower polygenic scores for cognition and EA (Supplementary Table 4).
Data analysis
Prior to analyses, polygenic scores were standardized and residualized for genotyping and imputation batch, array effects, and the 10 first principal components to control for population stratification.
Unadjusted models
Linear regressions of offspring ADHD traits on each polygenic score were conducted. False Discovery Rate (FDR) of 5% was applied to account for multiple testing [53], the number of tests being equal to 36, i.e. 12 parental factors times three family members.
Trio models
For each of the 12 parental factors, offspring ADHD traits were regressed onto three polygenic scores, i.e., for father, mother, and child (Fig. 1). FDR was applied as above.
Complementary analyses
We considered parental factors for which at least one family member’s polygenic score significantly predicted offspring ADHD traits in the trio models, jointly in one model across parental factors. This was to assess whether correlations between polygenic scores could affect estimates of genetic nurture and genetic transmission. Unadjusted and trio models were also conducted on the sample with complete phenotypic data available (prior to multiple imputation for missingness), as well as on the two ADHD subscales (inattention and hyperactivity-impulsivity).
All analyses controlled for child sex and year of birth. We tested whether estimates from the unadjusted, trio and across factor models differed significantly across models (pΔβ < 0.05) [54].
Results
Sample demographic characteristics are summarized in Table 1. Results from unadjusted and trio models are shown in Fig. 2 and Supplementary Table 5.

ADHD Attention-deficit/hyperactivity disorder, Autism Autism spectrum disorder, EA Educational attainment. ADHD scores were standardized to have a mean of zero and a standard deviation of 1. Trio results adjusted for polygenic scores of other family members. All models adjusted for child sex and year of birth. N = 19,506 family trios. * indicates estimates that remained significant after correction for multiple testing (36 tests) at a false-discovery rate (FDR) of 5%.
Polygenic scores for ADHD
As reported in Fig. 2, the offspring and both parental ADHD polygenic scores predicted offspring ADHD traits in unadjusted models (i.e., not adjusting for polygenic scores of other family members). In trio models, controlling for parental polygenic scores, the effect of offspring polygenic scores was not attenuated and remained significant indicating direct genetic effects on ADHD traits (illustrated as path c in Fig. 1). Conversely, parental associations were attenuated compared to estimates from unadjusted models (pΔβ = 1.48 × 10−14 for paternal and pΔβ = 9.95 × 10−17 for maternal estimates), findings which indicate that genetic transmission accounted for associations between parental polygenic scores and offspring ADHD traits, with little evidence for genetic nurture (paths f and m in Fig. 1).
Direct genetic effects and genetic transmission
For smoking polygenic scores, Fig. 2 shows a similar pattern of results suggesting direct genetic effects and attenuation of parental polygenic score associations in trio models (pΔβ = 3.93 × 10−7 for paternal and pΔβ = 2.12 × 10−10 for maternal estimates) – findings in line with the genetic transmission of risk. Offspring’s polygenic scores for cognitive performance and EA similarly indicated direct genetic effects. The paternal polygenic score for EA showed evidence of attenuation between unadjusted and trio models (pΔβ = 1.42 × 10−8), consistent with the genetic transmission.
Genetic nurture
The maternal polygenic score for neuroticism was associated with offspring ADHD traits in unadjusted models and remained significant in trio models, which supports genetic nurture as a mechanism linking maternal neuroticism to offspring ADHD traits.
The findings suggested some genetic nurture effects in unexpected directions, which should be interpreted with caution (see Discussion). The paternal polygenic score for alcohol use was negatively associated with offspring ADHD traits in both unadjusted and trio models, which would suggest genetic nurture as a mechanism linking paternal liability to drink more drinks per week with decreased ADHD traits in offspring. In trio models only, higher maternal polygenic scores for cognitive performance and EA associated with increased offspring ADHD traits.
To further investigate the negative genetic nurture effect of paternal alcohol use reported above, we reran the analyses using polygenic scores for alcohol dependence [55, 56]. This additional analysis indicated no direct genetic effect (β = 0.01, 95% CI: −0.02 to 0.04), nor genetic nurture effects from either parent (paternal β = 0.004, 95% CI: −0.02 to 0.03 and maternal β = 0.02, 95% CI: −0.01 to 0.04; Supplementary Table 6).
Some parental polygenic scores were associated with offspring traits in unadjusted models, and while confidence intervals for genetic nurture estimates overlapped with zero, trio model estimates showed little evidence of attenuation (suggesting genetic transmission) for paternal autism (pΔβ = 0.220) and for maternal autism (pΔβ = 0.183), depression (pΔβ = 0.051), and cannabis use (pΔβ = 0.796). As such, genetic nurture effects may not have been supported due to limited power of current polygenic scores.
Complementary analyses
Results on the sample prior to multiple imputation of missing phenotypic data (Supplementary Table 7) were consistent with results obtained after imputation. Compared to effect estimates reported in Fig. 2, estimates from the across-parental factor model did not differ significantly in magnitude (Supplementary Table 8).
Results for trio models on offspring inattention and hyperactivity-impulsivity subscales (Supplementary Tables 9, 10) yielded findings similar to those for the total ADHD scores. However, for hyperactivity-impulsivity, the unexpected genetic nurture estimates for paternal alcohol use and for maternal cognition and EA, as well as the direct genetic effects for offspring cognition did not reach statistical significance after correction for multiple testing.
Discussion
We examined whether genetic transmission or genetic nurture most likely explain the intergenerational transmission of ADHD traits. Specifically, we estimated the associations of polygenic scores indexing genetic liability for putative parental (risk and protective) factors and offspring ADHD traits. We used a multivariate within-family trio design that simultaneously models polygenic scores for parents and offspring. The trio design provides estimates of genetic nurture, unconfounded by direct genetic effects due to parent-to-offspring genetic transmission. Conversely, it provides estimates of direct genetic effects on offspring ADHD traits accounting for confounding by genetic nurture effects. Several parental polygenic scores correlated with offspring ADHD traits prior to accounting for genetic transmission, including ADHD itself, autism and smoking, maternal scores for depression, neuroticism and cannabis use, and paternal scores for EA and alcohol use. Trio models provided strong evidence that the offspring polygenic scores for ADHD and smoking predicted ADHD traits while parental effects were no longer evident and attenuated compared to the unadjusted parental effects. These findings support the role of parent-to-child genetic transmission of ADHD, smoking and EA-associated alleles as important mechanisms underlying associations between these parental factors and their children’s ADHD traits, with little evidence of genetic nurture. Evidence of genetic nurture was found in the case of maternal neuroticism since higher maternal polygenic liability associated with children’s ADHD traits in trio models.
Genetic transmission versus genetic nurture
We found that both parental ADHD polygenic scores associated with offspring ADHD traits prior to accounting for genetic transmission. However, that was no longer the case in within-trio analyses and associations between parental polygenic scores and offspring ADHD attenuated significantly, consistent with the genetic transmission. This finding is in line with a previous polygenic score study in a Dutch sample [34]. An adoption study found evidence that ADHD traits in biological mothers predicted child ADHD traits, in line with genetic transmission [26]. The study also found an association of ADHD traits in adoptive mothers and child ADHD, possibly consistent with nurture effects. However, the latter association may arise from reverse causation – e.g., the adoptive mother may find it harder to focus or stay attentive with a hyperactive child. Such reverse causation cannot happen in the design we used since offspring traits cannot change parental genotypes.
Similar patterns were observed for smoking, suggesting that genetic transmission partly explains the association between parental smoking and offspring ADHD traits. These findings are consistent with evidence from multiple genetically informed and causal inference methods, showing that the association between maternal smoking and offspring ADHD traits is likely due to genetic transmission, rather than to the impact of smoking per se [28, 57,58,59]. Furthermore, our findings show that the offspring polygenic score for smoking is independently associated with ADHD traits in 8-year-old children, i.e. prior to smoking initiation. This confirms that genetic variants implicated in smoking liability are associated with the emergence of ADHD independently of (parental) smoking itself. GWAS for smoking have identified variants implicated in dopaminergic pathways [60], which are also implicated in ADHD [40]. Such variants can contribute to smoking liability in parents, but in children, can also contribute to the emergence of ADHD. This genetic transmission of risk can inflate or generate spurious phenotypic associations between parental smoking and child ADHD.
We found evidence that maternal genetic liability to neuroticism not transmitted to their offspring associated with offspring ADHD traits – findings in line with environmental mechanisms of transmission (e.g., genetic nurture). Neuroticism reported by parents have been phenotypically associated with parenting difficulties [61], which may explain the association we found between maternal genetic variation and offspring traits.
Contrary to previous evidence that children’s ADHD is associated with lower parental cognitive performance and EA [8, 12], we found little evidence that maternal polygenic scores for these factors predicted offspring ADHD traits. After accounting for genetic transmission, we found associations with maternal predisposition to higher EA and cognition, a finding which was not robust across complementary analyses. Instead, our findings suggest that direct genetic transmission likely explains the direction of these genetic nurture effects since offspring predisposition to lower EA and cognition remained stronger predictors of their ADHD traits in trio models.
We also found that paternal genetic predisposition to drinking fewer drinks per week associated with offspring ADHD traits in both unadjusted and trio models, albeit not robustly so in complementary analyses of the hyperactivity-impulsivity subscale. A possible reason for this direction of association may be that lower polygenic scores for the number of drinks consumed per week index lower socioeconomic status [62]. Nonetheless, our results do not support the notion that more regular parental alcohol use is an environmental risk factor contributing to children’s ADHD despite previous evidence [16, 17, 19, 20]. Furthermore, trio models using polygenic scores for alcohol dependence (instead of quantity of drinks per week) did not support genetic nurture effects.
Implications for intergenerational psychiatry
Our findings suggest that genetic transmission plays an important role in explaining associations between parental factors and their children’s ADHD traits. Genetic transmission means that phenotypes arise separately in parents and children due to genetic influences they share, but independently of the environment created by parents. Conversely, genetic nurture effects imply that some parental factors matter as they foster environments that affect their offspring (either within the home like parenting or outside like school selection). Implications for interventions are diametrically opposed. In the case of genetic nurture, the effects of interventions targeting parental factors would be expected to affect child attention and hyperactive-impulsive behaviours and cascade across generations. In the case of sole genetic transmission, interventions directly targeting parental factors may successfully improve parental health, but benefits are unlikely to transfer to the child’s attention and hyperactive-impulsive behaviours. Instead, interventions directly aiming to support regulation of attention, activity and impulsivity in children may be more beneficial, whether such interventions are medical, school-based or parent-mediated [63]. Such findings for ADHD traits contrast with findings for EA in children where substantial genetic nurture effects have been found [31, 32]. As such, we should not be expecting universal patterns when it comes to explaining the role of intergenerational risk factors in children’s developmental outcomes. Emerging genetically informed methods [24, 31, 64, 65] should shortly render a detailed depiction of the intergenerational transmission of risk for psychiatric traits.
Limitations
Despite relying on the largest genotyped cohort of trios so far, power may be an issue to detect small intergenerational effects, especially given the limited accuracy of current polygenic scores. However, power unlikely accounts for our finding that genetic transmission is prominent in explaining the intergenerational transmission of risk for ADHD. For example, maternal genetic nurture effects for ADHD were close to zero with an upper CI of 0.023, whereas the lower CI for child-direct genetic effects was 0.075. Replications of findings in multiple cohorts will help with addressing limitations such as generalizability and power. In the meanwhile, while our findings clearly demonstrate the importance of genetic risk transmission, the limited evidence of genetic nurture effects should be considered as suggestive rather than definitive.
Although polygenic scores can, under strict assumptions, provide estimates of the causal effect of risk factors equal to those obtained from Mendelian randomisation [66], explicit causal inference methods such as intergenerational Mendelian randomization should be implemented in future analyses if genetic nurture effects were to be detected.
While mothers rated their children’s ADHD traits (model outcome), polygenic scores for the child, the father and even the mother (model predictors) did not depend on maternal ratings, limiting possible biases due to shared raters. However, maternal liability to ADHD as indexed by polygenic scores may still influence how they rate their children’s traits. Replication of our findings with non-parental ADHD ratings would be informative. We also did not account for variation in parental involvement in children’s upbringing (e.g., one parent less involved after separation). Future studies should test whether genetic nurture effects are moderated by parental involvement (e.g., larger for the more involved parent).
We cannot rule out possible bias due to assortative mating or population stratification. For educational attainment, maternal and paternal polygenic scores were correlated (r = 0.11), which suggests assortative mating (in Fig. 1, path a will not be near zero). However, simulation research indicates that assortative mating and population stratification may lead to inflated estimates of genetic nurture while estimates of direct parent-to-offspring genetic transmission effects are expected to be unaffected [33]. As such, our findings of direct genetic effects and limited evidence for genetic nurture should hold robust despite these possible limitations.
In conclusion, our study provides the most reliable evidence from genomic data to date, and the first evidence on many parental factors considered here, that genetic transmission may partly account for associations between some parental factors and children’s ADHD traits. Our findings also corroborate results from other genetically informed designs on the importance of genetics in the intergenerational transmission of ADHD. The implication for psychiatry is that the benefits of interventions targeting parental risk factors (e.g., parental ADHD or smoking) are likely to be restricted to the parental generation rather than extend across generations to the child’s ADHD. Directly targeting children’s traits should be more effective to improve children’s outcomes.
Data availability
The study website provides details on how to access data and information on the available variables (https://www.fhi.no/en/studies/moba/for-forskere-artikler/viktige-dokumenter-for-moba-forskere/). GWAS summary statistics used to compute polygenic scores are available from publicly available repositories from GWASatlas (https://atlas.ctglab.nl/) and the Psychiatric Genomics Consortium website (https://www.med.unc.edu/pgc/download-results/).
References
Duarte CS, Monk C, Weissman MM, Posner J. Intergenerational psychiatry: a new look at a powerful perspective. World Psychiatry. 2020;19:175–6.
Conley D, Domingue BW, Cesarini D, Dawes C, Rietveld CA, Boardman JD. Is the Effect of Parental Education on Offspring Biased or Moderated by Genotype? Socio Sci. 2015;2:82–105.
Galéra C, Côté SM, Bouvard MP, Pingault J-B, Melchior M, Michel G, et al. Early risk factors for hyperactivity-impulsivity and inattention trajectories from age 17 months to 8 years. Arch Gen Psychiatry. 2011;68:1267–75.
Lindblad F, Ringbäck Weitoft G, Hjern A. Maternal and paternal psychopathology increases risk of offspring ADHD equally. Epidemiol Psychiatr Sci. 2011;20:367–72.
Willoughby MT, Pek J, Greenberg MT. Parent-Reported Attention Deficit/Hyperactivity Symptomatology in Preschool-Aged Children: Factor Structure, Developmental Change, and Early Risk Factors. J Abnorm Child Psychol. 2012;40:1301–12.
Van Batenburg-Eddes T, Brion MJ, Henrichs J, Jaddoe VWV, Hofman A, Verhulst FC, et al. Parental depressive and anxiety symptoms during pregnancy and attention problems in children: a cross-cohort consistency study. J Child Psychol Psychiatry. 2013;54:591–600.
Birmaher B, Axelson D, Goldstein B, Monk K, Kalas C, Obreja M, et al. Psychiatric disorders in preschool offspring of parents with bipolar disorder: the Pittsburgh Bipolar Offspring Study (BIOS). Am J Psychiatry. 2010;167:321–30.
Cheung K, Theule J. Parental Psychopathology in Families of Children with ADHD: A Meta-analysis. J Child Fam Stud. 2016;25:3451–61.
Joelsson P, Chudal R, Uotila J, Suominen A, Sucksdorff D, Gyllenberg D, et al. Parental psychopathology and offspring attention-deficit/hyperactivity disorder in a nationwide sample. J Psychiatr Res. 2017;94:124–30.
Eilertsen EM, Hannigan LJ, McAdams TA, Rijsdijk FV, Czajkowski N, Reichborn-Kjennerud T, et al. Parental Prenatal Symptoms of Depression and Offspring Symptoms of ADHD: A Genetically Informed Intergenerational Study. J Atten Disord. 2020:1087054720914386.
Koutra K, Roumeliotaki T, Kyriklaki A, Kampouri M, Sarri K, Vassilaki M, et al. Maternal depression and personality traits in association with child neuropsychological and behavioral development in preschool years: Mother-child cohort (Rhea Study) in Crete, Greece. J Affect Disord. 2017;217:89–98.
Foulon S, Pingault J-B, Larroque B, Melchior M, Falissard B, Côté SM. Developmental predictors of inattention-hyperactivity from pregnancy to early childhood. PloS one. 2015;10:e0125996–e0125996.
Hegelund ER, Flensborg-Madsen T, Vassard D, Niclasen J, Mortensen EL. Parental socioeconomic position and risk of ADHD in offspring: a cohort study of 9648 individuals in Denmark 1976-2013. Eur Child Adolesc Psychiatry. 2019;28:685–93.
Russell AE, Ford T, Williams R, Russell G. The Association Between Socioeconomic Disadvantage and Attention Deficit/Hyperactivity Disorder (ADHD): A Systematic Review. Child Psychiatry Hum Dev. 2016;47:440–58.
Torvik FA, Eilertsen EM, McAdams TA, Gustavson K, Zachrisson HD, Brandlistuen R, et al. Mechanisms linking parental educational attainment with child ADHD, depression, and academic problems: a study of extended families in The Norwegian Mother, Father and Child Cohort Study. J Child Psychol Psychiatry. 2020: https://doi.org/10.1111/jcpp.13197.
Froehlich TE, Anixt JS, Loe IM, Chirdkiatgumchai V, Kuan L, Gilman RC. Update on environmental risk factors for attention-deficit/hyperactivity disorder. Curr Psychiatry Rep. 2011;13:333–44.
Thapar A, Cooper M, Eyre O, Langley K. Practitioner Review: What have we learnt about the causes of ADHD? J Child Psychol Psychiatry. 2013;54:3–16.
Roncero C, Valriberas-Herrero I, Mezzatesta-Gava M, Villegas JL, Aguilar L, Grau-López L. Cannabis use during pregnancy and its relationship with fetal developmental outcomes and psychiatric disorders. A systematic review. Reprod Health. 2020;17:25.
Molina BSG, Pelham WE, Lang AR. Alcohol Expectancies and Drinking Characteristics in Parents of Children with Attention Deficit Hyperactivity Disorder. Alcohol Clin Exp Res. 1997;21:557–66.
Eilertsen EM, Gjerde LC, Reichborn-Kjennerud T, Ørstavik RE, Knudsen GP, Stoltenberg C, et al. Maternal alcohol use during pregnancy and offspring attention-deficit hyperactivity disorder (ADHD): a prospective sibling control study. Int J Epidemiol. 2017;46:1633–40.
Larsson H, Chang Z, D’Onofrio BM, Lichtenstein P. The heritability of clinically diagnosed attention deficit hyperactivity disorder across the lifespan. Psychol Med. 2014;44:2223–9.
Eilertsen EM, Gjerde LC, Kendler KS, Røysamb E, Aggen SH, Gustavson K, et al. Development of ADHD symptoms in preschool children: Genetic and environmental contributions. Dev Psychopathol. 2019;31:1299–305.
Polderman TJ, Benyamin B, de Leeuw CA, Sullivan PF, van Bochoven A, Visscher PM, et al. Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nat Genet. 2015;47:702–9.
Pingault J-B, O’Reilly PF, Schoeler T, Ploubidis GB, Rijsdijk F, Dudbridge F. Using genetic data to strengthen causal inference in observational research. Nat Rev Genet. 2018;19:566–80.
McAdams TA, Hannigan LJ, Eilertsen EM, Gjerde LC, Ystrom E, Rijsdijk FV. Revisiting the Children-of-Twins Design: Improving Existing Models for the Exploration of Intergenerational Associations. Behav Genet. 2018;48:397–412.
Harold GT, Leve LD, Barrett D, Elam K, Neiderhiser JM, Natsuaki MN, et al. Biological and rearing mother influences on child ADHD symptoms: revisiting the developmental interface between nature and nurture. J Child Psychol Psychiatry. 2013;54:1038–46.
Silberg JL, Maes H, Eaves LJ. Unraveling the effect of genes and environment in the transmission of parental antisocial behavior to children’s conduct disturbance, depression, and hyperactivity. J Child Psychol Psychiatry. 2012;53:668–77.
Thapar A, Rice F, Hay D, Boivin J, Langley K, van den Bree M, et al. Prenatal smoking might not cause attention-deficit/hyperactivity disorder: evidence from a novel design. Biol Psychiatry. 2009;66:722–7.
Munafò MR, Higgins JPT, Davey Smith G. Triangulating Evidence through the Inclusion of Genetically Informed Designs. Cold Spring Harb Perspect Med. 2021;11:a040659.
Pingault JB, Richmond R, Davey Smith G. Causal Inference with Genetic Data: Past, Present, and Future. Cold Spring Harb Perspect Med. 2022;12:a041271.
Bates TC, Maher BS, Medland SE, McAloney K, Wright MJ, Hansell NK, et al. The Nature of Nurture: Using a Virtual-Parent Design to Test Parenting Effects on Children’s Educational Attainment in Genotyped Families. Twin Res Hum Genet. 2018;21:73–83.
Kong A, Thorleifsson G, Frigge ML, Vilhjalmsson BJ, Young AI, Thorgeirsson TE, et al. The nature of nurture: Effects of parental genotypes. Science. 2018;359:424–8.
Demange PA, Hottenga JJ, Abdellaoui A, Eilertsen EM, Malanchini M, Domingue BW, et al. Estimating effects of parents’ cognitive and non-cognitive skills on offspring education using polygenic scores. Nat Commun. 2022;13:4801.
de Zeeuw EL, Hottenga J-J, Ouwens KG, Dolan CV, Ehli EA, Davies GE, et al. Intergenerational Transmission of Education and ADHD: Effects of Parental Genotypes. Behav Genet. 2020;50:221–32.
Magnus P, Birke C, Vejrup K, Haugan A, Alsaker E, Daltveit AK, et al. Cohort Profile Update: The Norwegian Mother and Child Cohort Study (MoBa). Int J Epidemiol. 2016;45:382–8.
Magnus P, Irgens LM, Haug K, Nystad W, Skjaerven R, Stoltenberg C. Cohort profile: the Norwegian Mother and Child Cohort Study (MoBa). Int J Epidemiol. 2006;35:1146–50.
Paltiel L, Anita H, Skjerden T, Harbak K, Bækken S, Kristin SN, et al. The biobank of the Norwegian Mother and Child Cohort Study–present status. Norsk epidemiologi. 2014;24:1–2.
Silva RR, Alpert M, Pouget E, Silva V, Trosper S, Reyes K, et al. A rating scale for disruptive behavior disorders, based on the DSM-IV item pool. Psychiatr Q. 2005;76:327–39.
Oerbeck B, Overgaard KR, Pripp AH, Reichborn-Kjennerud T, Aase H, Zeiner P. Early Predictors of ADHD: Evidence from a Prospective Birth Cohort. J Atten Disord. 2020;24:1685–92.
Demontis D, Walters RK, Martin J, Mattheisen M, Als TD, Agerbo E, et al. Discovery of the first genome-wide significant risk loci for attention deficit/hyperactivity disorder. Nat Genet. 2019;51:63–75.
Purves KL, Coleman JRI, Meier SM, Rayner C, Davis KAS, Cheesman R, et al. A major role for common genetic variation in anxiety disorders. Mol Psychiatry. 2019;25:3292–303.
Grove J, Ripke S, Als TD, Mattheisen M, Walters RK, Won H, et al. Identification of common genetic risk variants for autism spectrum disorder. Nat Genet. 2019;51:431–44.
Stahl EA, Breen G, Forstner AJ, McQuillin A, Ripke S, Trubetskoy V, et al. Genome-wide association study identifies 30 loci associated with bipolar disorder. Nat Genet. 2019;51:793–803.
Pasman JA, Verweij KJH, Gerring Z, Stringer S, Sanchez-Roige S, Treur JL, et al. GWAS of lifetime cannabis use reveals new risk loci, genetic overlap with psychiatric traits, and a causal influence of schizophrenia. Nat Neurosci. 2018;21:1161–70.
Howard DM, Adams MJ, Clarke TK, Hafferty JD, Gibson J, Shirali M, et al. Genome-wide meta-analysis of depression identifies 102 independent variants and highlights the importance of the prefrontal brain regions. Nat Neurosci. 2019;22:343–52.
Savage JE, Jansen PR, Stringer S, Watanabe K, Bryois J, de Leeuw CA, et al. Genome-wide association meta-analysis in 269,867 individuals identifies new genetic and functional links to intelligence. Nat Genet. 2018;50:912–9.
Nagel M, Jansen PR, Stringer S, Watanabe K, de Leeuw CA, Bryois J, et al. Meta-analysis of genome-wide association studies for neuroticism in 449,484 individuals identifies novel genetic loci and pathways. Nat Genet. 2018;50:920–7.
Pardinas AF, Holmans P, Pocklington AJ, Escott-Price V, Ripke S, Carrera N, et al. Common schizophrenia alleles are enriched in mutation-intolerant genes and in regions under strong background selection. Nat Genet. 2018;50:381–9.
Karlsson Linnér R, Biroli P, Kong E, Meddens SFW, Wedow R, Fontana MA, et al. Genome-wide association analyses of risk tolerance and risky behaviors in over 1 million individuals identify hundreds of loci and shared genetic influences. Nat Genet. 2019;51:245–57.
Watanabe K, Stringer S, Frei O, Umićević Mirkov M, de Leeuw C, Polderman TJC, et al. A global overview of pleiotropy and genetic architecture in complex traits. Nat Genet. 2019;51:1339–48.
Lee JJ, Wedow R, Okbay A, Kong E, Maghzian O, Zacher M, et al. Gene discovery and polygenic prediction from a genome-wide association study of educational attainment in 1.1 million individuals. Nat Genet. 2018;50:1112–21.
Coombes BJ, Ploner A, Bergen SE, Biernacka JM. A principal component approach to improve association testing with polygenic risk scores. Genet Epidemiol. 2020;44:676–86.
Benjamini Y, Hochberg Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J R Stat Soc Ser B Methodol. 1995;57:289–300.
Clogg CC, Petkova E, Haritou A. Statistical Methods for Comparing Regression Coefficients Between Models. Am J Socio. 1995;100:1261–93.
Knopik VS, Heath AC, Jacob T, Slutske WS, Bucholz KK, Madden PAF, et al. Maternal alcohol use disorder and offspring ADHD: disentangling genetic and environmental effects using a children-of-twins design. Psychol Med. 2006;36:1461–71.
Walters RK, Polimanti R, Johnson EC, McClintick JN, Adams MJ, Adkins AE, et al. Transancestral GWAS of alcohol dependence reveals common genetic underpinnings with psychiatric disorders. Nat Neurosci. 2018;21:1656–69.
Thapar A, Rice F. Family-Based Designs that Disentangle Inherited Factors from Pre- and Postnatal Environmental Exposures: In Vitro Fertilization, Discordant Sibling Pairs, Maternal versus Paternal Comparisons, and Adoption Designs. Cold Spring Harb Perspect Med. 2020;11:a038877.
Rice F, Langley K, Woodford C, Davey Smith G, Thapar A. Identifying the contribution of prenatal risk factors to offspring development and psychopathology: What designs to use and a critique of literature on maternal smoking and stress in pregnancy. Dev Psychopathol. 2018;30:1107–28.
Gustavson K, Ystrom E, Stoltenberg C, Susser E, Surén P, Magnus P, et al. Smoking in Pregnancy and Child ADHD. Pediatrics 2017;139:e20162509.
Liu M, Jiang Y, Wedow R, Li Y, Brazel DM, Chen F, et al. Association studies of up to 1.2 million individuals yield new insights into the genetic etiology of tobacco and alcohol use. Nat Genet. 2019;51:237–44.
Prinzie P, Stams GJ, Deković M, Reijntjes AH, Belsky J. The relations between parents’ Big Five personality factors and parenting: a meta-analytic review. J Pers Soc Psychol. 2009;97:351–62.
Marees AT, Smit DJA, Ong JS, MacGregor S, An J, Denys D, et al. Potential influence of socioeconomic status on genetic correlations between alcohol consumption measures and mental health. Psychol Med. 2020;50:484–98.
Daley D, Van Der Oord S, Ferrin M, Cortese S, Danckaerts M, Doepfner M, et al. Practitioner Review: Current best practice in the use of parent training and other behavioural interventions in the treatment of children and adolescents with attention deficit hyperactivity disorder. J Child Psychol Psychiatry. 2018;59:932–47.
Eilertsen EM, Jami ES, McAdams TA, Hannigan LJ, Havdahl AS, Magnus P, et al. Direct and Indirect Effects of Maternal, Paternal, and Offspring Genotypes: Trio-GCTA. Behav Genet. 2021;51:154–61.
Young AI, Frigge ML, Gudbjartsson DF, Thorleifsson G, Bjornsdottir G, Sulem P, et al. Relatedness disequilibrium regression estimates heritability without environmental bias. Nat Genet. 2018;50:1304–10.
Dudbridge F. Polygenic Mendelian Randomization. Cold Spring Harb Perspect Med. 2021;11:a039586.
Acknowledgements
This project has received funding from the European Research Council (ERC) under the European Union’s Horizon 2020 research and innovation programme attributed to JBP (grant agreement No. 863981). JBP and WB are supported by the Medical Research Foundation 2018 Emerging Leaders 1st Prize in Adolescent Mental Health (MRF-160-0002-ELP-PINGA). WB was supported by the Wellcome Trust [224092/Z/21/Z]. The Research Council of Norway supported EME (#262177), EY (#262177 and #288083), NMD (#295989), TRK, EC and RBA (#274611), AH (#288083 and #300668) and OAA (#223273, #273291). AH and LJH was supported by the South-Eastern Norway Regional Health Authority (2020022 and 2018058/2019097, respectively). GDS and NMD work in a Unit supported by the Medical Research Council for the Integrative Epidemiology Unit (MC_UU_00011/1) at the University of Bristol. EY is funded by the European Union (Grant agreement No. 101045526). Views and opinions expressed are however those of the authors only and do not necessarily reflect those of the European Union or the European Research Council Executive Agency. Neither the European Union nor the granting authority can be held responsible for them. The Norwegian Mother, Father and Child Cohort Study is supported by the Norwegian Ministry of Health and Care Services and the Ministry of Education and Research. We are grateful to all the participating families in Norway who take part in this ongoing cohort study. We thank the Norwegian Institute of Public Health (NIPH) for generating high-quality genomic data, as part of the HARVEST collaboration which is supported by the Research Council of Norway (#229624). For providing genotype data, we also thank deCODE Genetics, and the NORMENT Centre (funded by the Research Council of Norway (#223273), South East Norway Health Authority and KG Jebsen Stiftelsen). We further thank the Center for Diabetes Research, the University of Bergen for providing genotype data and performing quality control and imputation of the data funded by the ERC AdG project SELECTionPREDISPOSED, Stiftelsen Kristian Gerhard Jebsen, Trond Mohn Foundation, the Research Council of Norway, the Novo Nordisk Foundation, the University of Bergen, and the Western Norway Health Authorities (Helse Vest). This work was performed on the TSD (Tjeneste for Sensitive Data) facilities, owned by the University of Oslo, operated and developed by the TSD service group at the University of Oslo, IT-Department (USIT; tsd-drift@usit.uio.no). The analyses were performed on resources provided by Sigma2 - the National Infrastructure for High Performance Computing and Data Storage in Norway. Disclaimer: Data from the Norwegian Patient Registry has been used in this publication. The interpretation and reporting of these data are the sole responsibility of the authors, and no endorsement by the Norwegian Patient Registry is intended nor should be inferred.
Author information
Authors and Affiliations
Contributions
JBP and WB were responsible for statistical analysis, and JBP, WB, EC and LH contributed to phenotypic and genetic data preparation. JBP, WB, EY and AH contributed to the study conception and design. JBP, WB, BW, LH, EME, EC, OAA, HA, MT, RBA, GDS, CS, NMD, TRK, EY and AH were responsible for data interpretation. JBP, WB and AH drafted the manuscript, and all authors critically reviewed the manuscript and made important intellectual contributions.
Corresponding author
Ethics declarations
Competing interests
OAA is a consultant to HealthLytix, with no conflict of interest relevant to this work. The other authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pingault, JB., Barkhuizen, W., Wang, B. et al. Genetic nurture versus genetic transmission of risk for ADHD traits in the Norwegian Mother, Father and Child Cohort Study. Mol Psychiatry 28, 1731–1738 (2023). https://doi.org/10.1038/s41380-022-01863-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41380-022-01863-6
This article is cited by
-
Estimating the impact of transmitted and non-transmitted psychiatric and neurodevelopmental polygenic scores on youth emotional problems
Molecular Psychiatry (2024)
-
Examining intergenerational risk factors for conduct problems using polygenic scores in the Norwegian Mother, Father and Child Cohort Study
Molecular Psychiatry (2024)
-
Gene–environment interactions in human health
Nature Reviews Genetics (2024)
-
Parental education and children’s depression, anxiety, and ADHD traits, a within-family study in MoBa
npj Science of Learning (2024)
-
Less for more? Cuts to child benefits, family adjustments, and long-run child outcomes in larger families
Journal of Population Economics (2024)
