
Abstract
Long-chain polyunsaturated fatty acids (LC PUFA) are associated with bone mass in animals and human adults, yet no data exist for human infants. Thus, the objective of this study was to establish that LC PUFA status is associated with bone mass in healthy infants. Thirty mother-infant pairs were studied for LC PUFA status by measuring maternal and cord blood red blood cells (RBC) for arachidonic acid (AA), eicosapentaenoic acid (EPA), and DHA. Infant anthropometry and lumbar spine 1–4, femur and whole-body bone mineral content (BMC) were measured within 15 d of delivery. Maternal and infant LC PUFA were tested for their relationship to BMC using Pearson correlation and backward step-wise regression analyses. At birth, the average gestational age was 39.3 ± 1.1 wk and body weight was 3433 ± 430 g. Cord RBC AA was positively correlated with whole-body BMC, AA:EPA positively correlated with lumbar spine 1–4 BMC and femur BMC. Maternal RBC AA was positively correlated with whole-body BMC. After accounting for infant weight using regression, whole-body BMC was positively predicted by cord RBC AA but none of the maternal LC PUFA; lumbar spine 1–4 BMC was positively predicted by cord RBCAA:EPA ratio but negatively by maternal DHA; and femur BMC was not predicted by cord LC PUFA but was negatively predicted by maternal DHA. Imbalances among the n-6 and n-3 LC PUFA by term gestation are associated with lower bone mass, suggesting that the maternal diet should be balanced in n-6 and n-3 LC PUFA.
Similar content being viewed by others


Main
It has been suggested that modulation of the intrauterine environment may offer primary prevention of osteoporosis later in life (1). A number of factors are known to affect human infant bone mass at birth, including maternal calcium intake (2), physical activity (1), and smoking (1). Maternal intake of fat and phosphorous (3) and smoking (4) while pregnant are linked with the child's subsequent bone mass until at least 8 y. The nutrients studied to date include primarily the macronutrients and minerals but not other nutrients such as the LC PUFA.
The role of LC PUFA in bone health is a relatively new subject of study. Thus, the science is controversial in revealing the effects of LC PUFA on bone and the responsible mechanisms of action. Of the LC PUFA, dietary supplementation with combinations of AA (20:4 n-6), EPA (20:5 n-3), or DHA (22:6 n-3) are positively associated with bone mass in infant animals (5–8). Among these studies, the LC PUFA supplementation ranged from a low amount of ∼1 g/100 g fat using semi-purified AA and DHA combined in piglets 6 to 17 g/100 g fat using fish oil in chicks (8). However, feeding too much fish oil (>80 g/100 g fat) as a source of n-3 LC PUFA during rapid growth postweaning is detrimental to bone growth and biomechanics of tibia of male rabbits (9) and spine of female rats (10). Feeding rat dams soybean oil (n-6 and n-3 essential fatty acids in a 9:1 ratio) compared with linseed oil (predominantly n-3 essential fatty acids) or sunflower seed oil (predominantly n-6 essential fatty acids) during late gestation and throughout lactation programs for higher long bone mass in the offspring (11), as observed by higher length, cortical cross-sectional area, and BMC at 30 wk of age (11), well after peak bone mass is achieved (12). To date, LC PUFA status acquired in utero has not been studied for an association with neonatal bone mass in animals or humans.
In humans, during pregnancy, dietary LC PUFA are preferentially transported to the fetus (13), resulting in higher LC PUFA values in RBC membranes from cord blood than in maternal blood (14). Thus, it is unclear whether maternal or infant LC PUFA status, or both, should be examined in relationship to infant bone mass. In newborn infants, the internationally recognized assessment of bone mass includes whole-body and lumbar spine measurements made using DXA (15). The best predictor of infant BMC is weight for whole body (16) and lumbar spine (17–20) with little to no effect of ethnicity or gender (20). Other regions such as the long bones are not typically assessed, although they are an important component of skeletal growth. Although measurement of femur is not a standard measurement, it could offer additional information regarding the effects of LC PUFA on bone inasmuch as research in piglets demonstrated effects in femur (6). One human study reports on infant femur BMC, although only in validating DXA measurements (19). Thus, the objective of this study was to establish that LC PUFA status (of mother or infant) is associated with whole-body and regional BMC in newborn infants after accounting for body weight.
SUBJECTS AND METHODS
Participants.
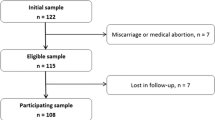
The study protocol was reviewed and approved by the University of Manitoba Ethics Review Board. The head obstetrical nurse screened all women admitted to the Winnipeg Health Sciences Centre (49.54°N) to deliver their infant (all days except holidays) until the sample size of 30 was achieved (August 2001 through April 2003). For entry to the study, all infants had to be born at appropriate weight for gestational age and sex as determined using the Centers for Disease Control growth charts and weight between the 5th and 95th percentiles (21), free of congenital malformations and with adequate vitamin D status (defined as >27.5 nmol 25(OH)D/L in plasma (22), and screened using a RIA (DiaSorin, Stillwater, MN)). Mothers had to have had ultrasound dating to confirm gestational age of the infant and be healthy as indicated by absence of gestational diabetes, hypertension, and any chronic medical therapy, and abstain from use of nonmedicinal drugs such as cocaine, marijuana, and alcohol. Women were entered into the study if they agreed to cord blood sampling and follow up where DXA was used to assess bone mass of the infant.
Sampling and biochemistry.
Cord blood at birth and maternal blood (within 48 h of delivery) were collected into heparinized vacutainers and plasma separated from RBC (2000 g) followed by nitrogen flush and storage at –80°C until analysis. The cord sample of RBC was taken to reflect infant LC PUFA status because RBC do not cross the placenta and would not be affected by changes in transport during parturition. Cord blood samples were stored as quickly as possible after collection, but, due to the time requirement for written consent, the average time between birth and storage was 13 h. Maternal samples were all stored within 4 h of collection and consistently taken between 0800 and 1000 h.
Total fatty acids were measured in RBC membranes within 4 wk of collection. RBC rather than plasma fatty acids were measured to reflect relatively stable long-term status over the last trimester of gestation (23) when bone mineralization would have been maximal (24,25). Total lipid was extracted using a modified method of Folch (26). The RBC membrane sample (250 L) was slowly added to 2.7 mL methanol and vortexed. Chloroform (3.3 mL) with 0.02% butylated hydroxytoluene and an internal standard, C17, were then added and the sample was centrifuged. The solvent layer was transferred and washed again with 1.5 mL NaCl (0.73%) and 2 mL chloroform and centrifuged followed by two more washes. The solvent layer was again removed, evaporated under nitrogen, dissolved in 1 mL toluene, and transmethylated by adding 1.2 mL of 3N methanolic HCL (Supelco, Bellefonte, PA) heating at 80°C for 1 h. Fatty acid methyl esters were recovered by the sequential addition of water and petroleum ether with final reconstitution in hexane. Fatty acid methyl esters were then separated using gas chromatography (Varian Star 3400, Mississauga, Canada) with hydrogen as the carrier gas. The gas chromatograph was equipped with a 30-m capillary column (DB225, 25% cyanopropylphenyl, J&W Scientific, Folsom, CA), 8100 autosampler, integrator, and flame ionization detector. Samples were injected at an initial temperature of 180°C and the column temperature was increased to a final temperature of 220°C at a rate of 3°C/ min. The detector temperature was 300°C. Fatty acid methyl esters were identified by comparison with retention times of Supelco 37 component fatty acid methyl esters mixture (Supelco). Fatty acids from carbon 14:0 to 24:1 were quantified and LC PUFA expressed as grams per 100 g total fatty acids.
Anthropometry and bone mass.
Birth weight and gestational age (based on ultrasound dating between 8 and 13 wk) were obtained from the medical record as were maternal age at time of delivery, weight gain during pregnancy, and height. Mothers were asked to self-identify their ethnicity.
At a follow-up visit (within 15 d of birth), infant weight was measured to the nearest gram on an electric scale (SB 32000; Mettler-Toledo Inc, Greifensee, Switzerland). DXA (Hologic QDR 4500A, Waltham, MA) was used to measure whole-body, lumbar spine 1–4, and total left femur BMC in array mode. The infants wore a light sleeper with no metal or plastic, a diaper, and were wrapped in a receiving blanket. BMC was expressed as absolute grams. Only scans with no movement artifacts were accepted for analysis as per Koo et al. (27), and, thus, the sample size for whole body, spine, and femur may not equal 30.
Sample size and statistical analysis.
Because there are no data upon which to base a sample size estimate for a relationship between infant BMC and LC PUFA status, a sample size of 30 was used to provide for a reasonable estimate of normal variation within the population (28). In total, 84 cord and maternal samples were collected. Those not used were inadequate in vitamin D (n = 27), as reported upon elsewhere (29), or if bone mass was not able to be measured within 15 d of birth due to rescheduling (n = 13) or sample attrition (n = 14).
Descriptive data are expressed as mean ± SD unless otherwise indicated. Relationships between maternal and cord blood LC PUFA and between maternal or cord RBC LC PUFA and BMC were conducted using Pearson correlation analysis. A p value <0.05 was taken as indicative of a probable relationship.
To determine whether LC PUFA status contributes to prediction of BMC, linear regression models were constructed using backward step-wise regression analyses. Predictor variables for BMC included infant weight at time of follow-up plus maternal or infant LC PUFA status. Maternal and infant LC PUFA status were run separately. Variables entered to represent LC PUFA status were RBC AA, DHA, EPA, AA:EPA, AA:DHA, and AA:EPA+DHA, because these LC PUFA have previously been studied for relationships to bone in animals. No variables were forced into the model. A p value of <0.05 was considered to indicate a probable relationship between BMC and the predictor variables.
RESULTS
Descriptive data.
The average gestational age at birth of the 30 infants (n = 16 females, n = 14 males) was 39.3 ± 1.1 wk, with an average birth weight of 3433 ± 430 g. Overall, the weights ranged from the 10th to 95th percentiles for sex (21). By design, all infants had adequate vitamin D status, with an average serum 25(OH)D of 45.8 ± 11.7 nmol/L. The average BMC was 73.5 ± 10.7 g for whole body (n = 22), 2.34 ± 0.37 g for lumbar spine 1–4 (n = 29), and 2.83 ± 0.56 g for femur (n = 29). These infants were born to mothers aged 29.3 ± 6.6 y who were on average 162 ± 7 cm in height with an average pregnancy weight gain of 16.4 ± 12.6 kg. The majority of mothers were white (n = 27), with one each self-identifying as Asian, East Indian, and First Nations (native Canadian).
The average RBC LC PUFA from cord blood and maternal blood are reported in Table 1. Maternal AA was not correlated with cord AA (r = 0.26, p = 0.17), nor was maternal EPA with cord EPA (r = 0.13, p = 0.49). However, maternal DHA correlated with cord DHA (r = 0.64, p = 0.0002).
Correlation analyses.
Correlation analysis of cord RBC LC PUFA with infant BMC yielded three significant relationships: AA with whole-body BMC (r = 0.61, p = 0.0032); AA:EPA with lumbar spine 1–4 BMC (r = 0.44, p = 0.0206) and femur BMC (r = 0.39, p = 0.044). Only maternal RBC AA was significantly related to whole-body BMC (r = 0.52, p = 0.014).
Regression analyses.
Infant whole-body BMC was best predicted by the regression equation: BMC = –47.467 + 20.0 (weight in kilograms) + 3.413 (cord RBC AA g/100 g fatty acids), n = 22, R2 = 0.76. No maternal LC PUFA contributed to prediction of infant whole-body BMC after accounting for weight: BMC = –8.270 + 24.0 (weight in kilograms), n = 22, R2 = 0.65.
For infant lumbar spine 1–4 BMC, infant weight did not contribute to the regression line such that BMC = 1.843 + 0.004 (cord RBC AA:EPA ratio), n = 29, R2 = 0.19. Maternal DHA plus infant weight, however, better predicted infant lumbar spine BMC = 1.252 + 0.487 (weight in kg) – 0.161 (maternal RBC DHA g/100 g fatty acids), n = 29, R2 = 0.33. This relationship is demonstrated in Figure 1a using lumbar spine BMC expressed as grams per kilogram body weight relative to maternal DHA.

Relationship between maternal RBC membrane DHA and infant lumbar spine (a) or femur (b) BMC expressed as grams per kilogram body weight, n = 29 for spine (r = –0.46, p = 0.012), n = 28 for femur (r = –0.45, p = 0.018).
Femur BMC was not predicted by any cord RBC LC PUFA: BMC = 0.613 + 0.640 (weight in kilograms), n = 29, R2 = 0.20. After accounting for weight, maternal DHA contributed to prediction of femur BMC such that BMC = 1.092 + 0.769 (weight in kilograms) – 0.245 (maternal RBC DHA g/100 g fatty acids), n = 29, R2 = 0.36. This relationship is demonstrated in Figure 1b using femur BMC expressed as grams per kilogram body weight relative to maternal DHA.
DISCUSSION
This is the first report of a relationship between infant bone mass and LC PUFA status of mother or infant after healthy full-term gestation. Cord RBC AA was related to whole-body BMC, and cord AA:EPA was related to lumbar spine BMC in both correlation analysis plus regression analysis. In addition, femur BMC was correlated with cord RBC AA:EPA. These relationships are similar to those previously reported in piglets (n = 12), whereby liver total AA was associated with whole-body BMC (r = 0.75, p < 0.01), lumbar spine 1–4 (r = 0.71, p < 0.01) and femur BMC (r = 0.67, p < 0.01) (5) The relationship between cord RBC AA:EPA with lumbar spine 1–4 and femur BMC in the infants may be indicative of an imbalance between n-6 and n-3 fatty acids with consequences to bone. A high AA:EPA ratio (associated with higher BMC) may arise from relatively more AA or relatively less EPA and thus the relationship to bone is unclear. However, too little EPA may limit development of BMC as observed recently in piglets (n = 36) fed varied amounts of AA (0.5–2.0% of dietary fat) and DHA (0.1–0.4% of dietary fat) (30). For example, liver AA:EPA negatively correlated with whole-body (r = –0.49, p = 0.009), lumbar spine 1–4 (r = –0.39, p = 0.04) and femur BMC (r = –0.48, p = 0.01). In the piglet study, no EPA was added to the diet, resulting in very low EPA status but high AA in the 2.0% of dietary fat group. In other animals, feeding high amounts of EPA is detrimental to bone biomechanics in rabbits (9) and rats (10). In regular chow-fed rats weaned from dams fed a high n-3 diet during late gestation and throughout lactation, femur BMC is reduced compared with rats of dams fed a balanced n-6 and n-3 diet (11). Regarding the present study, it seems as well that maternal n-3 LC PUFA is associated with lower BMC human infants. For example, maternal DHA was negatively associated with infant spine and femur BMC after accounting for infant body weight in the regression analyses. Thus, just as for growing animals (9,11), it seems that higher n-3 LC PUFA status in pregnant humans is not associated with advantages to neonatal BMC.
By design, the infants and mothers in this study were all healthy, giving rise to expected measurements of infant weight and BMC. The whole-body BMC value of 73.5 g was basically identical to that reported for other Canadian infants (31). No Canadian values exist for lumbar spine 1–4 or femur BMC, but values obtained in the United States for lumbar spine 1–4 were similar, at 2.14 g for infants born between 27 and 42 wk gestation (20). Because the infants in the present study were all healthy and of similar gestational age at birth, the range in BMC values was relatively narrow compared with that expected if small and large weight for date infants were also studied. Inclusion of small infants was ruled out for the present study because such infants are known to have low bone mass (32) at birth and also lower LC PUFA status (33). Both would have confounded the results.
Regarding cord and maternal RBC LC PUFA measured as grams per 100 g total fatty acids, few datasets exist for RBC values in both cord and maternal samples with which to compare. However, the values for cord blood and maternal RBC LC PUFA were similar to those reported at delivery by Montgomery et al. (34). The relatively low DHA status was expected, inasmuch as Canadian women have a low intake of DHA, ranging from 24 to 524 mg DHA daily (35). In this study, the highest RBC DHA value was only 5.5 g/100 g fatty acids. Thus, whether higher maternal intakes of DHA or DHA status are associated with neonatal BMC is unknown. If the regression equations are used to calculate infant BMC using the mean values for infant weight (3.419 kg) and maternal DHA (3.7 g/100 g RBC fatty acids), the contribution of a 1–2 g elevation (i.e., 4.7–5.7 g/100 g RBC fatty acids) in maternal DHA would translate into a 7–14% reduction (–0.161 to –0.322 g) in lumbar spine BMC and a 9–18% reduction (–0.245 to –0.490 g) in femur BMC. For both spine and femur, such elevation in maternal DHA would place the infant 0.44–0.88 SD below the mean. Few studies have quantified the effect of maternal complications or diet on lumbar spine BMC and none have examined femur BMC. However, the size of this effect is similar to a low (<600 mg/d) versus high calcium (∼3 g/d) maternal diet between 22 wk gestation and term delivery, whereby infant spine BMC is 15% lower at birth in the low calcium group (2).
Although assessment of maternal dietary intake of LC PUFA was not conducted in the present study, other researchers document that 4 wk of maternal supplementation of n-3 alpha-linolenic acid gives rise to higher plasma EPA but not DHA (36). Higher maternal intake of DHA through high-DHA eggs (37), fish oil (23), or semi-purified AA+DHA supplements (38) leads to elevated maternal DHA status. Total RBC DHA can be elevated from 4.69 to 7.15 g/100 g fatty acids in pregnant women after 9 wk of supplementation using sardines and fish oil providing 2.6 g n-3 fatty acids daily (23). In another study where women were supplemented with fish oil (4 g/d ∼ 3.4 g n-3 fatty acids) from 20 to 37 wk of pregnancy, total maternal RBC DHA was elevated from 6.9 to 11.0 g/100 g fatty acids, but AA status was compromised in both the mother and neonate (14). If the balance between AA and EPA or DHA is important to acquisition of BMC in utero, it is important to provide supplementation of both n-6 and n-3 LC PUFA. When DHA (0.57 g/d) and AA (0.26 g/d) are supplemented together, no decline in maternal AA is observed in plasma or RBC membranes (38). Given the associations between the various LC PUFA and BMC in this study, it is important to test for the optimal balance among n-6 and n-3 LC PUFA in the diet of pregnant women to support whole-body development. In theory, elevations in maternal DHA in the order of 4 g/100 g fatty acids, as reported by others (14), could limit bone mass in the neonate by ∼30%. Randomized controlled studies are required to test this possibility.
In summary, this study of healthy term-born infants reveals for the first time a relationship between perinatal maternal and infant LC PUFA status and neonatal BMC. Similar relationships have also been observed in growing animals, although not at birth. The women participating in this study did not undergo any intervention and thus the results require confirmation under controlled conditions including assessment of dietary LC PUFA intake. Maternal n-3 LC PUFA intake has been implicated in neurodevelopment of the infant until 4 y of age (39). The associations described in this report justify investigation of the safety, with respect to neonatal bone mass, of an increased maternal intake of AA and DHA in pregnancy. Such investigation should be extended through infancy to establish or rule out long-term implications to bone mass. Until then, whether optimizing maternal and neonatal LC PUFA status can offer a new element to the primary prevention of osteoporosis remains unknown.
Abbreviations
- AA:
-
arachidonic acid
- BMC:
-
bone mineral content
- EPA:
-
eicosapentaenoic acid
- LC PUFA:
-
long-chain polyunsaturated fatty acid
- RBC:
-
red blood cell
References
Godfrey K, Walker-Bone K, Robinson S, Taylor P, Shore S, Wheeler T, Cooper C 2001 Neonatal bone mass: influence of parental birthweight, maternal smoking, body composition, and activity during pregnancy. J Bone Miner Res 16: 1694–1703
Koo WW, Walters JC, Esterlitz J, Levine RJ, Bush AJ, Sibai B 1999 Maternal calcium supplementation and fetal bone mineralization. Obstet Gynecol 94: 577–582
Jones G, Riley MD, Dwyer T 2000 Maternal diet during pregnancy is associated with bone mineral density in children: a longitudinal study. Eur J Clin Nutr 54: 749–756
Jones G, Riley M, Dwyer T 1999 Maternal smoking during pregnancy, growth, and bone mass in prepubertal children. J Bone Miner Res 14: 146–151
Weiler HA 2000 Dietary supplementation of arachidonic acid is associated with higher whole body weight and bone mineral density in growing pigs. Pediatr Res 47: 692–697
Blanaru JL, Kohut JR, Fitzpatrick-Wong SC, Weiler HA 2004 Dose response of bone mass to dietary arachidonic acid in piglets fed cow milk-based formula. Am J Clin Nutr 79: 139–147
Watkins BA, Shen CL, Allen KG, Seifert MF 1996 Dietary (n-3) and (n-6) polyunsaturates and acetylsalicylic acid alter ex vivo PGE2 biosynthesis, tissue IGF-1 levels, and bone morphometry in chicks. J Bone Miner Res 11: 1321–1332
Watkins BA, Shen CL, McMurtry JP, Xu H, Bain SD, Allen KG, Seifert MF 1997 Dietary lipids modulate bone prostaglandin E2 production, insulin-like growth factor-1 concentration and formation rate in chicks. J Nutr 127: 1084–1091
Judex S, Wohl GR, Wolff RB, Leng W, Gillis AM, Zernicke R 2000 Dietary fish oil supplementation adversely affects cortical bone morphology and biomechanics in growing rabbits. Calcif Tissue Int 66: 443–448
Sirois I, Cheung AM, Ward WE 2003 Biomechanical bone strength and bone mass in young male and female rats fed a fish oil diet. Prostaglandins Leukot Essent Fatty Acids 68: 415–421
Korotkova M, Ohlsson C, Hanson LA, Strandvik B 2004 Dietary n-6:n-3 fatty acid ratio in the perinatal period affects bone parameters in adult female rats. Br J Nutr 92: 643–648
Wang L, Banu J, McMahan CA, Kalu DN 2001 Male rodent model of age-related bone loss in men. Bone 29: 141–148
Larque E, Demmelmair H, Berger B, Hasbargen U, Koletzko B 2003 In vivo investigation of the placental transfer of (13)C-labeled fatty acids in humans. J Lipid Res 44: 49–55
Dunstan JA, Mori TA, Barden A, Beilin LJ, Holt PG, Calder PC, Taylor AL, Prescott SL 2004 Effects of n-3 polyunsaturated fatty acid supplementation in pregnancy on maternal and fetal erythrocyte fatty acid composition. Eur J Clin Nutr 58: 429–437
Lewiecki EM, Kendler DL, Kiebzak GM, Schmeer P, Prince RL, El-Hajj Fuleihan G, Hans D 2004 Special report on the official positions of the International Society for Clinical Densitometry. Osteoporos Int 15: 779–784
Rigo J, De Curtis M, Picaud JC, Nyamugabo K, Senterre J 1998 Whole body calcium content in term and preterm neonates. Eur J Pediatr 157: 259–260
Kurl S, Heinonen K, Jurvelin JS, Lansimies E 2002 Lumbar bone mineral content and density measured using a Lunar DPX densitometer in healthy full-term infants during the first year of life. Clin Physiol Funct Imaging 22: 222–225
Salle BL, Braillon P, Glorieux FH, Brunet J, Cavero E, Meunier PJ 1992 Lumbar bone mineral content measured by dual energy X-ray absorptiometry in newborns and infants. Acta Paediatr 81: 953–958
Braillon PM, Salle BL, Brunet J, Glorieux FH, Delmas PD, Meunier PJ 1992 Dual energy x-ray absorptiometry measurement of bone mineral content in newborns: validation of the technique. Pediatr Res 32: 77–80
Koo WW, Hockman EM 2000 Physiologic predictors of lumbar spine bone mass in neonates. Pediatr Res 48: 485–489
Growth Curves 2000 Centers for Disease Control web site. Available at: http://www.cdc.gov/growthcurves/. Accessed February 14, 2004
Institute of Medicine 1997 Dietary Reference Intakes for Calcium, Phosphorus, Magnesium, Vitamin D, and Fluoride. National Academy Press, Washington, DC, pp 259
Connor WE, Lowensohn R, Hatcher L 1996 Increased docosahexaenoic acid levels in human newborn infants by administration of sardines and fish oil during pregnancy. Lipids 31: S183–S187
Ryan S, Congdon PJ, James J, Truscott J, Horsman A 1988 Mineral accretion in the human fetus. Arch Dis Child 63: 799–808
Ziegler EE, O'Donnell AM, Nelson SE, Fomon SJ 1976 Body composition of the reference fetus. Growth 40: 329–341
Folch J, Lees MSloane ST, Stanley GH 1957 A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem 226: 497–509
Koo WW, Walters J, Bush AJ 1995 Technical considerations of dual-energy X-ray absorptiometry-based bone mineral measurements for pediatric studies. J Bone Miner Res 10: 1998–2004
Whitley E, Ball J 2002 Statistics review 2: samples and populations. Crit Care 6: 143–148
Weiler H, Fitzpatrick-Wong S, Veitch R, Kovacs H, Schellenberg J, McCloy U, Yuen CK 2005 Vitamin D deficiency and whole-body and femur bone mass relative to weight in healthy newborns. CMAJ 172: 757–761
Mollard RC, Kovacs HR, Fitzpatrick Wong SC, Weiler HA 2005 Low levels of dietary arachidonic and docosahexaenoic acid improve bone mass in neonatal piglets, but higher levels provide no benefit. J Nutr 135: 505–512
Atkinson SA, Randall-Simpson J 2000 Factors influencing body composition of premature infants at term-adjusted age. Ann N Y Acad Sci 904: 393–399
Chunga Vega F, Gomez de Tejada MJ, Gonzalez Hachero J, Perez Cano R, Coronel Rodriguez C 1996 Low bone mineral density in small for gestational age infants: correlation with cord blood zinc concentrations. Arch Dis Child Fetal Neonatal Ed 75: F126–F129
Cetin I, Giovannini N, Alvino G, Agostoni C, Riva E, Giovannini M, Pardi G 2002 Intrauterine growth restriction is associated with changes in polyunsaturated fatty acid fetal-maternal relationships. Pediatr Res 52: 750–755
Montgomery C, Speake BK, Cameron A, Sattar N, Weaver LT 2003 Maternal docosahexaenoic acid supplementation and fetal accretion. Br J Nutr 90: 135–145
Innis SM, Elias SL 2003 Intakes of essential n-6 and n-3 polyunsaturated fatty acids among pregnant Canadian women. Am J Clin Nutr 77: 473–478
Francois CA, Connor SL, Bolewicz LC, Connor WE 2003 Supplementing lactating women with flaxseed oil does not increase docosahexaenoic acid in their milk. Am J Clin Nutr 77: 226–233
Smuts CM, Borod E, Peeples JM, Carlson SE 2003 High-DHA eggs: feasibility as a means to enhance circulating DHA in mother and infant. Lipids 38: 407–414
Otto SJ, van Houwelingen AC, Hornstra G 2000 The effect of supplementation with docosahexaenoic and arachidonic acid derived from single cell oils on plasma and erythrocyte fatty acids of pregnant women in the second trimester. Prostaglandins Leukot Essent Fatty Acids 63: 323–328
Helland IB, Smith L, Saarem K, Saugstad OD, Drevon CA 2003 Maternal supplementation with very-long-chain n-3 fatty acids during pregnancy and lactation augments children's IQ at 4 years of age. Pediatrics 111: e39–e44
Author information
Authors and Affiliations
Corresponding author
Additional information
This research was supported by grants from the Thorlakson Foundation Fund (University of Manitoba), Canadian Institutes of Health Research, and the Manitoba Health Research Council. H.W. is in receipt of a New Investigator Salary Award from the Canadian Institutes of Health Research. U.M. was in receipt of postdoctoral fellowship and H.K. a graduate scholarship from the Manitoba Institute of Child Health.
The study was conceived and manuscript written by H.W. S.F.-W. was responsible for the majority of the LC PUFA analysis. J.S., U.M., R.V., and H.K. were responsible for recruitment, sample collection, and follow-up measurements. J.K. was responsible for testing the stability of LC PUFA under the study conditions and was involved in the initial stages of the study. C.K.Y. participated in the development of the study and advised on recruitment methodology.
Rights and permissions
About this article
Cite this article
Weiler, H., Fitzpatrick-Wong, S., Schellenberg, J. et al. Maternal and Cord Blood Long-Chain Polyunsaturated Fatty Acids Are Predictive of Bone Mass at Birth in Healthy Term-Born Infants. Pediatr Res 58, 1254–1258 (2005). https://doi.org/10.1203/01.pdr.0000185129.73971.74
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/01.pdr.0000185129.73971.74
