
Abstract
Secretory immunoglobulin A (sIgA), the predominant antibody fraction of human milk, represents a major protective factor against neonatal infection. Until now, sIgA had been identified only in the humoral fraction of human milk. The aim of this study was to assess whether in human milk sIgA is also associated with the milk fat globule (MFG). Using anti-sIgA-agglutinated human MFG and immune fluorescence microscopy, we could demonstrate that sIgA is, in fact, associated with human MFG. In human MFG membranes separated by Western blotting, sIgA bands were specifically stained, suggesting that sIgA is strongly associated with the human MFG membrane. This may be of physiologic relevance, inasmuch as earlier we could show that a quantity of undigested and functional human MFG are in fact found in the stools of the newborn. This would allow an additional extension of the protective mechanisms of sIgA throughout the whole intestine.
Similar content being viewed by others
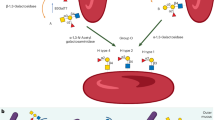


Main
Among the humoral anti-infectious factors, the immunoglobulins contained in human milk represent the major group for the protection of the neonate(1). Within this group, the largest share is made up of sIgA, which passes through the child's gastrointestinal tract virtually undigested and thus lends its protective effect to the entire intestinal passage(2). The fat component of human milk consists of numerous fat globules which are individually coated by a membrane. Generally accepted is the theory on the genesis of these fat globules, according to which they are produced by apocrine extrusion from the mammary gland cell. Briefly, fat globules are believed to be generated within the mammary gland epithelial cell, surrounded by a piece of cell membrane that buds out and finally separates from the cell into the duct(3). This theory finds support in the observation that after electrophoretic separation the milk fat globule membrane proteins and the plasma proteins of the mammary epithelial cell show a similar pattern(4).
We have recently demonstrated that the HMFGM not only serves as a vessel for milk fat, but it also exerts protective functions, inasmuch as through interaction between carbohydrates and bacterial lectins pathogenic microorganisms can be bound by the HMFGM and thus prevented from mucosal epithelial cell adhesion(5,6).
For bovine milk, immunoglobulins have been demonstrated both within the aqueous phase and bound to the milk fat globule membrane(7). The aim of this study was to explore whether sIgA is bound to HMFGM and thus may contribute to the protective function of HMFGM.
METHODS
Isolation of milk fat globules from human colostrum. Milk was collected from healthy mothers of term babies on the first to fifth day of lactation. The milk was diluted in an equal volume of PBS and centrifuged at 750 × g for 10 min at 4°C to obtain cream. The cream was washed between two and six times in 10 vol of sucrose-PBS (0.25 M sucrose containing 10% PBS, pH 7.4) to isolate washed HMFG.
Preparation of HMFGM. Freshly collected human breast milk from the first 5 days of lactation was pooled and skimmed by centrifugation at 3 000 × g for 60 min at 4°C. The cream was washed in 5 vol of 10 mM Tris buffer, pH 7.2, by centrifugation under the same conditions. The washed cream was suspended in 4 vol of 10 mM Tris buffer, pH 7.2, and shaken on a laboratory shaker at a temperature of 4°C for 60 min. Under continued shaking, the suspension was allowed to warm up to room temperature for 60 min. Finally, it was incubated at 40°C until butter had formed. The membranes were recovered as a pellet after centrifugation at 30 000 × g for 30 min at 30°C and washed twice under the same conditions before freeze-drying(8,9).
Microscopic plate agglutination. Washed HMFG prepared as described above were diluted in sucrose-PBS to an optical density of 1.5 (measured at 605 nm in a 1-cm cuvette). Twenty microliters of different antisera (for source of antisera see Table 1) was added to 2.5 µL of the HMFG suspension. After incubation for 120 min on glass slides in a humid chamber, the degree of agglutination was assessed microscopically.
Indirect immunofluorescence. HMFG were diluted in sucrose-PBS plus 0.5% BSA (sucrose-PBS-BSA). Ten microliters of different antisera (for source of antisera see Table 2, primary antibody) was added to 80 µL of the HMFG suspension. After incubation for 60 min at room temperature on a slow rotary shaker, the suspension was washed in 1 mL of sucrose-PBS-BSA by centrifugation at 200 × g for 5 min at room temperature. Twenty microliters of FITC-labeled antiserum (for source of antisera see Table 2, secondary antibody) was added to 200 µL of the preincubated HMFG suspension, and after 60 min of incubation in a dark humid chamber and washing as above, the degree of fluorescence was assessed microscopically.
Electrophoresis, Coomassie blue staining, and immunoblotting. For electrophoresis, the samples [10 mg/mL freeze-dried HMFGM, 1 mg/mL lactoferrin (Sigma Chemical Co., St. Louis, MO), 1 mg/mL purified human sIgA, prepared as described previously(10)] were preincubated in sample buffer (0.3% SDS for 10 min) or for denaturation and reduction in 0.3% SDS and 3% 2-mercaptoethanol for 10 min at 90°C. Proteins were separated in polyacrylamide gels (3-13%) in the presence of SDS (0.1%) at 40 mA. Gels were fixed and stained with Coomassie blue. Electroblotting onto nitrocellulose sheets (0.45 µm) was performed at 250 mA in Tris (25 mM)-glycine (192 mM)-methanol (20%) at pH 8.3. To block residual surface activity, the nitrocellulose sheets were immersed in PBS containing 5% BSA for 90 min at room temperature. For immunostaining the sheets were incubated for 60 min at 4°C with sheep anti-human sIgA (1:1 000 in PBS) (Nordic, Tilburg, The Netherlands) or with mouse anti-human SC (1:1 000 in PBS) which binds to the SC only (Sigma Chemical Co.). Alkaline phosphatase conjugated donkey anti-sheep IgG (1:10 000 in PBS; Sigma Chemical Co.) or rabbit anti-mouse IgG (1:10 000 in PBS; Sigma Chemical Co.) was added for 30 min after washing the samples in PBS-BSA. After four washing steps, the sheets were developed with 5-bromo-4-chloro-3-indoxyl phosphate (sodium salt) and nitro-blue-tetrazolium-chloride (Serva, Heidelberg, Germany) for 15 min at 37°C(11).
RESULTS
Agglutination. The glass slide agglutination studies showed significant agglutination of HMFG with antisera against sIgA and IgA (Table 1 and Fig. 1). Even increasing the number of washing steps before addition of the antibody did not alter the amount of agglutination observed. These additional washing steps were performed to rule out a contribution of loosely adsorbed protein from the aqueous phase of human milk.

a, HMFG, no antibody, 1:500 magnification. b, HMFG, incubated with sheep anti-human sIgA, 1:500 magnification.
There was no agglutination with the control antisera, which were targeted against other immunoglobulin classes or nonhuman immunoglobulin (Table 1 and Fig. 1).
Indirect immunofluorescence. Under the fluorescence microscope the fat globules stained markedly fluorescence-positive when labeled with antibodies targeted against IgA or sIgA. This fluorescence was not observed in negative controls with different antisera of other specificities (Table 2 and Fig. 2).

HMFG, 1:500 magnification, labeled with sheep anti-human sIgA and rabbit anti-sheep Ig (FITC-labeled).
Immunoblot and Coomassie blue staining. After Coomassie blue staining of the gels and Ponceau S staining of the blot and subsequent reduction, both the sIgA and the HMFGM lanes showed corresponding bands (figure not shown). These bands stained positive for anti-sIgA in the immunoblot (Fig. 3). The bands at approximately 180 kD in the sIgA lane should correspond to monomeric IgA (Fig. 3b). Three bands corresponding to the light chain with joining chain, the α-chain, and the SC could be identified (Fig. 3a). Similarly, the immunoblot with anti-SC showed identical bands with HMFGM and sIgA (Fig. 4).

SDS-PAGE immunoblot of sIgA, HMFGM, native (b) and under reducing conditions (a), immunodetection with sheep anti-human sIgA and donkey anti-sheep IgG, alkaline phosphatase conjugated.

SDS-PAGE immunoblot of sIgA and HMFGM, native, with mouse anti-human SC and rabbit anti-mouse IgG, alkaline phosphatase conjugated.
DISCUSSION
We showed not only that the sIgA of human milk is contained in the aqueous phase and is loosely associated with the HMFG, while originating from the aqueous phase, but that it is strongly bound to the HMFGM and can still be detected after membrane separation.
The human milk used was obtained between d 1 to 5 of lactation and processed immediately to get a maximal proportion of milk fat globule membranes and to avoid storage-induced artifacts.
For the detection of sIgA on the HMFG surface, a glass slide assay with different antisera was performed. In this assay, a specific agglutination with anti-sIgA antibodies was demonstrated. These results were highly reproducible. No agglutination was seen with sera against other antigens.
In the past, an association of sIgA with bovine milk fat globules has been demonstrated(7). Laven et al.(12) stained noncellular globules containing lipids in human milk with fluorochrome-labeled reagents specific for IgA and SC. In retrospect it appears likely that at least some of these elements were milk fat globules.
The glass slide agglutination assay was chosen because milk fat globules can be well demonstrated under the light microscope at a 100-fold magnification and because after incubation with specific antisera these fat globules form microscopically visible agglutinates.
To rule out an influence of non-membrane-bound components from the lipid phase, these were removed by copious washing of the lipid phase before the addition of antibody. This was accomplished by suspending the fat globules in iso-osmotic sucrose-PBS, slow spinning, and resuspending in new sucrose-PBS solution. The agglutination of milk fat globules by anti-sIgA was never affected, even after repeated washing.
The agglutination test with anti-human IgM failed, just as did the test with anti-human IgG. For IgG the result is explained by the low concentrations of IgG in human milk(13). Similarly, the findings for IgM can possibly be explained by its 3-fold lower concentration in comparison with IgA, by a steric inhibition due to the molecular structure of IgM, or by a combination thereof. Presumably there is also a different mode of secretion whereby IgM is not bound to the HMFGM. IgM is predominantly secreted by plasma cells, and it mostly lacks the SC, which is known to be bound by a membrane-integrated receptor.
Indirect fluorescence staining with FITC was preferred over direct immunofluorescence for its greater sensitivity and to augment the signal intensity. In our experiments we used polyclonal primary antibodies against the SC, the IgA Fc-fragment and the α-chain of the human IgA molecule. The FITC-labeled secondary antibodies were directed against the respective immunoglobulin subclasses of the primary antibody donors.
To neutralize nonspecific antibody binding, in these experiments we added BSA to the iso-osmotic diluent, which markedly reduced background fluorescence.
To investigate further the localization of IgA within the HMFGM fraction, we initially separated the HMFGM from the fat contained within these globules, then separated the membrane components using SDS-PAGE.
When comparing the band pattern of purified sIgA and HMFGM before and after reduction, in both Coomassie and Ponceau S stains three bands with equal migration patterns were detected under reducing conditions.
In comparison with purified sIgA from colostrum, the immunoblots with anti-sIgA (Fig. 3) and anti-SC (Fig. 4) stained at identical molecular weights. On blots of the native HMFGM sample a slowly migrating band on top of the gel represents the dimeric sIgA molecule with a molecular weight of 390 kD.
Using what is known about the physiology and immunobiology of secretory glands, e.g. in the gastrointestinal or bronchial tracts(14), we propose a thesis on the origin of the sIgA lining of the HMFGM.
In their Golgi apparatuses and endoplasmic reticula, the mammary gland epithelial cells produce fat globules and shuttle them through the cells toward their apical poles. By exocytosis they become encased in membranes that have been cut out of the apical cell membrane(3).
At the same time, plasma cells localized in the mammary gland epithelium produce dimeric secretory IgA molecules. The SC (expressed at the basolateral mammary gland cell surface) binds to the joining chain of the dimeric sIgA; these sIgA molecules can be taken up into the mammary gland epithelial cell by endocytosis.
Together with the J-chain and the SC, the sIgA molecule is shuttled through the mammary gland cell to the apical pole(15,16), where it is in part expressed on the cell surface and in part secreted into the aqueous phase of the human milk.
During the described budding process of the fat globule coating from the apical membrane of the glandular cell, it appears not inconceivable that via the SC receptor, which in part remains an integral component of the cell membrane, membrane-bound sIgA molecules along with bits of mammary cell membrane become components of the HMFGM(17). In addition to sIgA binding by the SC receptor, the firm binding of the richly glycosylated sIgA to the HMFGM might also be mediated by glycosyl transferases contained in the membrane(18).
It is not possible from our data to estimate the quantity of sIgA bound to the HMFGM. From a teleologic standpoint, the fast association of sIgA and the HMFGM appears adaptive, because at least a share of the imbibed milk fat globules can traverse the whole gastrointestinal passage unaltered(5). Thus the sIgA-mediated adhesion of microorganisms to the milk fat globule surface can prevent adhesion of these germs to the gut mucosa(6). Further studies will elucidate whether this proposed mechanism of bacterial neutralization by HMFGM-bound sIgA has some bearing in vivo.
Abbreviations
- sIgA:
-
secretory immunoglobulin A
- HMFG:
-
human milk fat globule
- HMFGM:
-
human milk fat globule membrane
- SC:
-
secretory component
References
Goldman AS 1993 The immune system of human milk: antimicrobial antiinflammatory and immunomodulation properties. Pediatr Infect Dis J 12: 664–671.
Davidson LA, Lonnerdal B 1987 Persistence of human milk proteins in the breast-fed infant. Acta Paediatr Scand 76: 733–740.
Moyer-Mileur L, Chan GM 1989 Milk membranes: origin, content, changes during lactation, and nutritional importance. In: Lebenthal E (ed) Textbook of Gastroenterology and Nutrition in Infancy. Raven Press, New York, 151–155.
Ceriani R, Thompson K, Peterson JA 1977 Surface differentiation antigens of human mammary epithelial cells carried on the milk fat globule. Proc Natl Acad Sci U S A 74: 582–586.
Schroten H, Hanisch FG, Plogmann R, Hacker J, Uhlenbruck G, Nobis-Bosch R, Wahn V 1992 Inhibition of adhesion of S-fimbriated Escherichia coli to buccal epithelial cells by human milk fat globule membrane components: a novel aspect of the protective function of mucins in the nonimmunoglobulin fraction. Infect Immun 60: 2893–2899.
Schroten H 1998 The benefits of human milk fat globule membranes against infection. Nutrition 14: 52–53.
Honkanen-Buzalski T, Sandholm M 1981 Association of bovine secretory immunoglobulins with milk fat globule membranes. Comp Immunol Microbiol Infect Dis 4: 329–342.
Dowben RM, Brunner JR, Philpott DE 1967 Studies on milk fat globule membranes. Biochim Biophys Acta 135: 1–10.
Imam A, Laurence JR, Neville AM 1981 Isolation and characterization of a major glycoprotein from milk-fat-globule membrane of human breast milk. Biochem J 193: 47–54.
Mestecky J, Kilian M 1985 Immunoglobulin A. Methods Enzymol 116: 37–75.
Towbin H, Staehelin T, Gordon J 1979 Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci U S A 76: 4350–4354.
Laven GT, Crago SS, Kutteh WH, Mestecky J 1981 Hemolytic plaque formation by cellular and noncellular elements of human colostrum. J Immunol 127: 1967–1972.
Goldman AS, Goldblum RM 1986 Immunoglobulins in human milk. In: Board RG (ed) Natural Antimicrobial Systems. International Diary Federation, UK, Bath University Press, Bath, 1–17.
Brandtzaeg P, Halstensen TS, Kett K, Krajci P, Kvale D, Rognum TO, Scott H, Sollid LM 1989 Immunobiology and immunopathology of human gut mucosa: humoral immunity and intraepithelial lymphocytes. Gastroenterology 97: 1562–1584.
Bienenstock J, Befus AD 1980 Mucosal immunology. Immunology 41: 249–270.
Mestecky J, McGhee JR, Crago SS, Jackson S, Kilian M, Kijono H, Babb JL, Michalek SM 1980 Molecular-cellular interactions in the secretory IgA response. J Reticuloendothel Soc 28: 45–60.
Mestecky J, Lue C, Russel MW 1991 Selective transport of IgA. Gastroenterol Clin North Am 20: 441–471.
Kobayashi K, Mafune N, Narimatsu H, Nakao H, Taniguchi N 1996 Secretory component, the receptor for polymeric immunoglobulin, has nothing to do with -galactosyltransferase in human milk. Immunol Lett 50: 99–104.
Acknowledgements
The authors thank Jiri Mestecky for supplying sIgA samples and critically reading the manuscript.
Author information
Authors and Affiliations
Rights and permissions
About this article
Cite this article
Schroten, H., Bosch, M., Nobis-Bosch, R. et al. Secretory Immunoglobulin A Is a Component of the Human Milk Fat Globule Membrane. Pediatr Res 45, 82–86 (1999). https://doi.org/10.1203/00006450-199901000-00014
Received:
Accepted:
Issue Date:
DOI: https://doi.org/10.1203/00006450-199901000-00014
This article is cited by
-
Function and dysfunction of plasma cells in intestine
Cell & Bioscience (2019)
