
Abstract
Dysregulated activation of the CDK4/6 kinases is a hallmark of most mammary-derived carcinomas. ATP-competitive inhibitors against this complex have been recently advanced in the clinic and have shown significant activity, particularly against tumors driven by the estrogen receptor (ER). However, resistance to these compounds has begun to emerge often months to years after their initiation. We investigated potential mechanisms of resistance using cell line models that are highly sensitive to this class of drugs. After prolonged exposure to the selective and potent CDK4/6 inhibitor LY2835219, clones emerged and several were found to harbor amplification of the CDK6 kinase. Amplification of CDK6 resulted in a marked increase in CDK6 expression and reduced response of the CDK4/6 target, phospho-Rb (pRb), to CDK4/6 inhibitors. Knockdown of CDK6 restored drug sensitivity, while enforced overexpression of CDK6 was sufficient to mediate drug resistance. Not only did CDK6 overexpression mediate resistance to CDK4/6 inhibitors but it also led to reduced expression of the ER and progesterone receptor (PR), and diminished responsiveness to ER antagonism. The reduced ER/PR expression after CDK4/6 inhibitor resistance was additionally observed in tumor biopsy specimens from patients treated with these drugs. Alternative mechanisms of resistance to CDK4/6 inhibitors such as loss of pRb and cyclin E1 overexpression also exhibited decreased hormone responsiveness, suggesting that the clinical paradigm of sequential endocrine-based therapy may be ineffective in some settings of acquired CDK4/6 resistance.
Similar content being viewed by others

Introduction
Estrogen-driven cell cycle progression in breast cancer is mediated, in part, through transcriptional regulation of cyclin D1 along with suppression of cell cycle inhibitors, such as p27 and p21.1, 2, 3 These factors also receive inputs from growth factor signaling cascades such as the phosphoinositide 3-kinase pathway and it has been speculated that such signals might stabilize cyclin D expression and thereby potentiate resistance to antiestrogen therapy in breast cancer.4, 5 As such, kinase inhibition of the cyclin D–CDK4/6 complex was developed as a therapeutic approach to hormone receptor-expressing breast cancer.6, 7, 8 This strategy has shown activity in the form of combinations of ATP-competitive inhibitors of CDK4/6 together with aromatase inhibition (letrozole) or estrogen receptor (ER) antagonism (fulvestrant).9, 10, 11 Despite the benefit of this approach clinically, tumor resistance develops in most patients in the metastatic setting and the basis for this resistance is unknown. In this study, we investigated mechanisms of acquired resistance to CDK4/6 inhibition. We established cell lines that were chronically exposed to the CDK4/6 inhibitor, LY2835219 (LY5219, abemaciclib), and identified amplification of CDK6 as a recurrent event. Overexpression of CDK6 promoted resistance to several CDK4/6 inhibitors as well as inhibitors of the ER. The data suggest that mechanisms that bypass the G1–S checkpoint may mediate resistance to both direct inhibitors of CDK4/6 and inhibitors whose targets primarily function through activation of cyclin D–CDK4/6.
Results
Generation of CDK4/6 inhibitor-resistant cells
To identify potential mechanisms of resistance to CDK4/6 inhibitors, we initially characterized the selectivity of the CDK4/6 inhibitor, LY5219, in cells. The compound has been previously demonstrated to be a potent and selective inhibitor of the CDK4 (half maximal inhibitory concentration (IC50)=2 nm) and CDK6 (IC50=9.9 nm) kinases, with CDK9 (57 nm), HIPK2 (31 nm) and DYRK2 (61 nm) being the only other kinases found to be inhibited below 100 nm.8 Given the well-established requirement for intact Rb expression for the pharmacologic efficacy of this class of drugs,12, 13 we compared the effects of the drug in Rb mutant and wild-type cells. We found potent inhibition of growth of the Rb wild-type cells MCF-7 (32 nm) and T47D (60 nm) compared with the Rb-null MDA-MB-468 cells (644 nm) (Supplementary Figure 1). These data identified LY5219 as a selective inhibitor of CDK4/6 kinases in cells at doses below 100 nm.
To generate ER+ cells resistant to CDK4/6 inhibitors, we exposed MCF-7 cells to LY5219 over a span of 21 weeks and collected the cells that grew out. These cells (designated MCF-7 resistant (MR)) had a significantly higher IC50 (231 nm) for inhibition of proliferation compared with parental cells (27.2 nm) (Figure 1a). To further confirm the reduced potency of LY5219 in these cells, we analyzed the cell cycle distribution of drug-treated cells by fluorescence-activated cell sorting assay. As expected, parental MCF-7 cells had a marked decrease in the proportion of cells in the S phase after 48 h exposure to 100 nm LY5219. By contrast, there was virtually no effect of the drug on the S-phase content of MR cells (Figure 1b). To ascertain if the resistance of the MR cells was specific to the LY5219 compound, we tested the growth response of parental and MR cells to other selective CDK4/6 inhibitors, PD0332991 (palbocicilib) and LEE011 (ribociclib). In both cases, MR cells required significantly higher drug concentrations (6–8-fold) for growth suppression (Supplementary Figure 2).

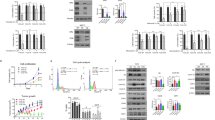
Derivation of ER+ cells resistant to CDK4/6 inhibitors. (a) MCF-7 and MR cells were treated with different concentrations of CDK4/6 inhibitor (LY2835219) for 5 days. Cell viability was measured via the alamarBlue assay. Inhibition of proliferation (%) is plotted against log drug concentration. Each data point represents the average of values obtained from three independent experiments. (b) MCF-7 and MR cells were treated with dimethyl sulfoxide (DMSO) or LY2835219 (100 nm) for 48 h and measured by fluorescence-activated cell sorting (FACS) cell cycle analysis. Results are reported as mean percent cell cycle distribution with standard errors (*P<0.05, **P<0.01, Student’s t-test). (c) MCF-7 and MR cells were treated with 100 nm LY2835219 and collected at indicated times. Immunoblots were performed with the indicated antibodies. (d) MCF-7 and MR cells were treated with increasing concentrations of LY2835219 and harvested after 24 h; lysates were then subjected to immunoblotting.
To understand whether the resistance of MR cells was due to changes in the response of CDK4/6 kinases to the drug or a downstream, Rb-independent mechanism, we examined the effect of drug on Rb phosphorylation. Parental MCF-7 cells showed decreases in phospho-Rb (pRb) (S780 and S795) as early as 4 h with complete loss by 24 h in 100 nm LY5219 (Figure 1c). Meanwhile, MR cells were unaffected up to 8 h post-dosing and only showed minor decreases at 24–48 h. Corresponding to these data, cyclin A2 and cyclin E2 showed decreases in expression by 24 h in parental but not in MR cells. In accordance with these results, parental cells showed complete inhibition of pRb (S780 and S795) with doses as low as 100 nm LY5219, whereas doses up to 1 μm did not block Rb phosphorylation in MR cells (Figure 1d). Taken together, the data suggest that MR cells are resistant to ATP-competitive CDK4/6 inhibitors in a manner that allows persistent Rb phosphorylation despite high levels of drug.
High CDK6 expression in CDK4/6 inhibitor-resistant cells
As MR cells displayed no change in basal pRb but impaired sensitivity to CDK4/6 inhibitors, we speculated a component of the CDK4/6–cyclin D1 complex may be altered in these cells. We initially performed next- generation sequencing (MSK-IMPACT; Supplementary Table 1) of 450 cancer-associated genes in the MR and parental cells.14 Nearly all exonic mutations were shared between the two cell lines with no obvious acquired missense mutation that might promote drug resistance. We next examined the levels of expression of components of the CDK4/6–cyclin D1 complex along with known signaling complexes responsible for its activation in these cells. Most strikingly, we observed a sevenfold increase in CDK6 mRNA levels in the MR cells, with concomitant increases in CDK6 protein levels as compared with parental cells (Figures 2a and b). To understand the basis for the increase in CDK6 expression, we examined the copy number of the CDK6 locus using fluorescent in situ hybridization (FISH) with probes against CDK6 and the chromosome 7 centromere. The analysis revealed that within the population of MR cells, a fraction showed increased CDK6 copy number (3–21 copies) as compared with parental cells (3–6 copies) where no such high-level amplification was observed (Figure 2c). Altogether, the data are consistent with an acquired amplification in the drug target, CDK6, being present in the MR cells. To ascertain whether this phenomenon was unique to MR cells, we generated CDK4/6 inhibitor-resistant cells in the T47D breast cancer model. As observed for the MR cells, selective doses of CDK4/6 inhibitors had limited effects on proliferation and cell cycle progression in T47D-resistant (TR) cells. Once again, elevated levels of CDK6 mRNA and protein were also identified in TR cells in this case together with decreases in Rb1 expression (Supplementary Figure 3), further supporting the notion that CDK6 may be susceptible to overexpression in ER+ breast cancers treated with CDK4/6 inhibitors.

Elevated CDK6 levels in CDK4/6 inhibitor-resistant cells. (a) MCF-7 and MR cells were cultured in normal media without LY2835219 for 24 h. The mRNA levels for the indicated genes were measured by quantitative RT–PCR analysis. Data are shown as mean±s.d. of three independent experiments (**P<0.01, Student’s t-test). (b) MCF-7 and MR cells were treated with LY2835219 and collected at 0, 4, 8, 24 and 48 h. Cell lysates were immunoblotted with indicated antibodies. (c) MCF-7 and MR cells were analyzed by FISH, using a CDK6 probe (Spectrum Red) and centromeric repeat probe for chromosome 7 (Spectrum Green). Representative images of metaphase and interphase cells are shown. The yellow arrow denotes CDK6 high cells.
Acquired CDK6 amplification promotes resistance to CDK4/6 inhibitors
In some instances, overexpression or amplification of a kinase has been associated with heightened sensitivity to inhibitors of that kinase. However, in this case, CDK6 overexpression was ‘acquired’ in the context of long-term exposure to CDK4/6 inhibitors. To determine if overexpression of CDK6 contributed to drug resistance, we suppressed CDK6 levels in these resistant cells using short hairpin RNAs (shRNAs) against CDK6. Greater than 90% CDK6 knockdown was confirmed by quantitative reverse transcription–PCR (RT–PCR) (Figure 3a). We found that reducing CDK6 levels close to those found in parental cells restored the sensitivity of MR cells to LY5219, resulting in comparable IC50s for proliferation (MCF-7=31 nm, MR-sh negative control=218 nm, MR-sh CDK6=17 nm) (Figure 3b). Moreover, the reduction of CDK6 levels in MR cells was associated with restored sensitivity of pRb (S795 and S780) to LY5219 (Figure 3c). These data suggest that CDK6 overexpression is necessary for MR cells to resist LY5219 treatment.

CDK6 overexpression is necessary for MR drug resistance. (a) CDK6 mRNA levels are detected by quantitative RT–PCR in MCF-7, MR-sh NC (MR cells stably transfected with control plasmid) and MR-sh CDK6 (MR cells stably transfected with CDK6-shRNA containing plasmid) cells. Data represent the mean of three independent experiments (**P<0.01, Student’s t-test). (b) Inhibition of proliferation by CDK4/6 inhibitor for MCF-7, MR-sh NC and MR-sh CDK6 cells. % Inhibition of proliferation is plotted against the log concentration of the CDK4/6 inhibitor LY2835219. Each data point represents the mean value of six biological replicates obtained from three independent experiments. (c) MCF-7, MR-sh NC and MR-sh CDK6 cells were treated with LY2835219 (100 nm) and collected at indicated times. Lysates were immunoblotted with the designated antibodies.
To ascertain whether CDK6 overexpression is sufficient to promote resistance to CDK4/6 inhibitors, we enforced expression of CDK6 through the use of a constitutive long terminal repeat promoter in three different ER+ cell lines: MCF-7, T47D and CAMA-1. After selecting for cells with stable CDK6 overexpression, we examined their response to CDK4/6 inhibitor treatment in cell proliferation assays. High levels of CDK6 resulted in a 4–11-fold increase in IC50 (LY5219) and also led to a reduced pRb response (S780 and S795) to the drug. Moreover, overexpression of CDK6 in T47D cells (T47D-CDK6-N1) prevented dephosphorylation of Rb by 100 nm LY5219 after 48 h (Figures 4a–d and Supplementary Figure 4). We further noted that overexpression of CDK6 was associated with suppression of cyclin D1 levels while drug treatment was associated with greater increases in cyclin D1 levels in parental cells consistent with CDK4/6- regulating cyclin D1 expression in these cells. We went on to examine clones expressing different levels of CDK6 and found that higher CDK6 levels correlated with higher LY5219 IC50s. For instance, T47D-CDK6-N2 expressed five times more protein than parental T47D, while T47D-CDK6-N1 expressed 20 times more protein. Correspondingly, the IC50 of LY5219 for T47D-CDK6-N2 cells was 143.6 nm, while the IC50 for T47D-CDK6-N1 cells was 960.4 nm (Figures 4e and f). The data from these ER+ cell lines is consistent with CDK6 overexpression directly promoting resistance to CDK4/6 inhibitors.

CDK6 overexpression promotes resistance to CDK4/6 inhibitors. (a and c) MCF-7 (a) and T47D (c) cells stably transfected with CDK6 overexpression vector or empty control vector were treated with increasing concentrations of LY2835219 for 5 days. Cell viability was measured by the alamarBlue assay. Inhibition of proliferation (%) is plotted against log drug concentration. Each data point represents the average of values±s.d. obtained from three independent experiments. (b) MCF-7-control and MCF-7 CDK6-N2 cells were treated with different concentrations of LY2835219 for 24 h and collected with lysates subject to immunoblotting. (d) T47D-control and T47D-CDK6-N1 were treated with 100 nm LY2835219 and collected at indicated times. Immunoblots were performed with antibodies to the indicated proteins. (e) T47D cells stably transfected with either CDK6 expression vector or empty vector were treated with different concentrations of LY2835219 for 5 days. Cell proliferation was measured by the alamarBlue assay and plotted as % inhibition of proliferation against log concentration of inhibitor. Inset table shows the IC50 for each cell line. T-CDK6-N1, T-CDK6-N2 and T-CDK6-N3 represent individual stable cell lines. (f) T47D-control, T47D-CDK6-N1, T47D-CDK6-N2 and T47D-CDK6-N3 cells were harvested and lysates immunoblotted for CDK6 and β-actin. The results from three independent experiments were quantified to determine relative intensity of CDK6 protein expression calculated as the ratio of T47D-CDK6/T47D-control in the graph.
Acquired CDK6 amplification and reduced ER/PR expression
One of the major drivers of cyclin D1–CDK4/6 activity in breast cancer cells is hormone-mediated activation of the ER.15 This induction of the CDK4/6 kinases has been shown to be essential for estrogen-driven cell proliferation.16, 17 Previous reports have suggested that E2F may be a key regulator of expression of ER.18, 19 Given the role of CDK4/6 in regulating free E2F levels, we sought to determine whether there might be changes in ER or PR expression in the resistant cells where the G1–S checkpoint was abrogated. In the MR cells, mRNA and protein levels of ER and PR were significantly reduced compared with the parental cells. Moreover, the expression of ER-regulated genes, such as XBP1 and MYC, was also markedly suppressed (Figures 5a and c). Similar ER and PR loss was further detected in the TR cells compared with parental T47D cell lines (Figures 5b and d).

CDK4/6 inhibitor resistance is associated with diminished ER/PR signaling. (a and b) mRNA from MCF-7 and MR (a) and T47D and TR (b) cells were analyzed by quantitative RT–PCR for levels of ESR1 and ER-regulated genes PGR, GREB1, XBP1, MYCand TFF1. Data shown are from triplicate experiments and reported as the mean±s.d. (*P<0.05, **P<0.01, Student’s t-test). (c and d) MCF-7 and MR (c) and T47D and TR (d) cells were treated with increasing concentrations of LY2835219 for 24 h and collected for immunoblotting of ER, PR and CDK6. (e) Quantitative RT–PCR analysis comparing mRNA levels for indicated genes from MCF-7-control and MCF-7 CDK6 cells. Data are shown as mean±s.d. of three independent experiments (*P<0.05, **P<0.01, Student’s t-test). (f) MCF-7-control and MCF-7 CDK6-N2 cells were treated with different concentrations of LY2835219 and collected after 24 h. Cell lysates were immunoblotted with the indicated antibodies. Data shown are representative of three independent experiments. (g) CAMA-1 and CR cells were cultured in normal media without LY2835219 for 24 h. The mRNA levels for the indicated genes were measured by real-time PCR analysis. Data are shown as mean±s.d. of three independent experiments (*P<0.05, **P<0.01, Student’s t-test). (h) MCF-7 and MR cells were cultured in LY2835219-free medium for 24 h. ChIP was performed with anti-polymerase II (Pol II) antibody or control immunoglobulin G (IgG). Primers to amplify the Pol II binding regions of ER promoters were used in qPCR to determine fold enrichment relative to IgG. Error bars represent s.d. of three independent experiments with three technical replicates each (**P<0.01, Student’s t-test).
To exclude the possibility that this was an MR-specific off-target effect because of exposure to drug, we examined the kinetics of loss of ER and noted that neither transient drug inhibition nor transient CDK6 overexpression led to declines in ER or PR expression in MCF-7 cells (Supplementary Figures 1A and 5A). Next, we assessed the ER and PR levels in our models engineered to overexpress CDK6 including MCF-7 (Figures 5e and f), T47D (Supplementary Figure 5B) and CAMA-1 (Supplementary Figure 5C). In all cases, we observed diminished ESR1 and PGR expression. By contrast, overexpression of CDK4 did not result in significant changes in PGR or ESR1 expression levels (Supplementary Figure 5D). It thus appeared that prolonged exposure to high levels of CDK6, associated with drug resistance, was necessary for ER downregulation.
To understand if these effects were unique to the CDK6-mediated models of resistance, we assessed ER and PR levels in another independent LY5219 resistance model, which was derived from CAMA-1 cells (CR) and was found to have overexpression of CCNE1 (Figure 5g). This model also showed reduced mRNA levels of ESR1 and PGR, suggesting that reduction of these receptors may be related to a variety of mechanisms that converge on inhibiting the G1/S checkpoint. Finally, to further understand the basis of loss of ER expression, we examined occupancy of RNA polymerase II at the ESR1 promoter using chromatin immunoprecipitation (ChIP) against Pol II (Figure 5h). We observed MR cells to have markedly reduced RNA Pol II occupancy at the ESR1 promoter compared with parental MCF-7 cells, suggesting that transcriptional repression of ER may underlie its diminished expression.
Acquired CDK6 amplification promotes hormone resistance
We next investigated whether the changes in ER/PR expression might be associated with a change in the dependence of these cells on the ER for growth. We examined the effect of the pure antiestrogen fulvestrant (ICI182, 780) on MCF-7 parental and MR cells and found that the resistant cells required significantly higher levels of ICI182 (IC50=48 nm) compared with parental cells (IC50=0.23 nm) to block the growth (Figure 6a). Moreover, selective ER modulators, such as 4-hydroxytamoxifen (4-OHT), also showed a higher IC50 (2030 nm) in MR cells compared with MCF-7 parental cells (113.2 nm) (Figure 6b). To further understand the basis for this insensitivity, we examined the effect of fulvestrant on ER/PR expression and activity in the MR and MCF-7 cells (Figure 6c). Increasing doses of ICI182 was able to downregulate ER and PR expression in both parental and MR cells; however, this downregulation was not associated with the same magnitude of pRb loss in the MR cells as in the parental cells.

CDK4/6 inhibitor resistance promotes diminished ER expression and activity. (a and b) Proliferation of MCF-7 and MR cells in the presence of different concentrations of ICI (a) or 4-hydroxytamoxifen (4-OHT) (b) was measured by the alamarBlue assay and plotted as % inhibition of proliferation against log concentration of inhibitor. Each data point represents the average value±s.d. of six replicates obtained in three independent experiments. (c) MCF-7 and MR cells were treated with different concentrations of ICI, collected after 24 h and then immunoblotted with the indicated antibodies.
To determine whether this loss of ER/PR expression and dependence might occur in patients, we examined a series of patients with ER+ breast cancer who received treatment with a CDK4/6 inhibitor and had tumor biopsies performed both before and after exposure to drug. We examined ER and PR levels using a conventional immunohistochemical scoring system used for clinical assessment of ER/PR expression (Table 1 and Supplementary Figure 6). Strikingly, in the small number of patients examined, it appeared that several patients had tumors that changed from ER+ to ER− or from PR+ to PR−. We unfortunately could not assess whether there was a loss of ER dependence in these patients as none of them underwent single-agent hormone therapy. Nevertheless, the data raise the possibility that a subset of patients who develop resistance to CDK4/6 inhibitors might manifest with associated tumoral changes in ER/PR levels and resistance to CDK4/6 inhibitors, as well as endocrine therapies such as fulvestrant.
Discussion
A number of clinically effective breast cancer therapies have been developed that work, in part, by indirectly reducing the activity of the CDK4/6–cyclin D complex.20 Foremost among these are hormonal therapies where ER regulation of cyclin D1 has been demonstrated to be critical to their effectiveness.21 Clinical resistance to these forms of therapy appears to involve reactivation of CDK4/6–cyclin D either through reactivation of ER (e.g. ESR1 mutation) or through other growth factor signaling pathways (e.g. ERBB2 amplification).22, 23, 24 Therefore, direct pharmacologic inhibition of the G1–S checkpoint kinases, CDK4 and CDK6, has long been considered a promising strategy.4, 6, 15 Selective inhibitors of CDK4/6 indeed have proven highly effective in large clinical trials of patients with advanced ER+ breast cancer; however, resistance to therapy has also frequently emerged.9, 10, 25, 26, 27 The basis for resistance in the clinical setting is not well understood, nor is there consensus on how to effectively manage patients after cancer progression on CDK4/6 inhibitor therapy. In this study, we sought to uncover mechanisms that might engender resistance to ER+ breast cancer cells previously sensitive to CDK4/6 inhibitors and identified alterations that not only led to CDK4/6 kinase inhibitor resistance but also resistance to hormonal therapies.
To identify alterations that promote resistance to CDK4/6 inhibitors, we exposed two different hormone-sensitive cell line models to the potent CDK4/6 kinase inhibitor, LY5219.8, 27 Long-term exposure of the models to drug led to multiple independent clones that had evidence for amplification of the CDK6 kinase. This was observed in two different cell lines and in experimental replicates conducted months apart. Although other changes might have also occurred in these cells over time, the sufficiency of CDK6 to promote resistance was clear from experiments in which CDK6 was knocked down in the resistant cells and in which CDK6 was overexpressed in the parental cells. In both cases, CDK6 overexpression was clearly correlated with a requirement for higher doses of inhibitor to cause cell growth inhibition.
How CDK6 overexpression promotes resistance to CDK4/6 inhibition is not yet clear. CDK6 is known to have kinase-independent functions;28, 29 however, the close correlation between the dose of kinase inhibitor causing loss of pRb and the dose blocking cell proliferation supports the premise that the kinase function of CDK6 is essential for resistance. Interestingly, overexpression of CDK4 was never observed in the models and enforced overexpression of CDK4 did not promote inhibitor resistance. In actuality, we often observed decreases in CDK4 expression in the resistant cells. One speculation is that the partner cyclin or other components of the complex influence inhibitor response. For instance, CDK6 may preferentially bind to cyclin D3 and this complex proves more drug resistant than the cyclin D1–CDK4 complex. Nevertheless, the data raise the possibility that compounds with even greater potency against CDK6 might be of clinical interest, particularly in the setting of acquired resistance.
In addition to amplification of CDK6, we also identified two models: one with mutation in Rb as well as CDK6 overexpression, and another with cyclin E1 overexpression. Both of them were well correlated with inactivity of CDK4/6 inhibitors. This observation was in line with prior reports on the essential role of pRb and CCNE1 in mediating the effects of CDK4/6 inhibitors.12, 30, 31 Taken together with our models demonstrating CDK6 amplification, the findings suggest that alterations that uncouple the G1–S checkpoint from hormone or growth factor regulation may prove to be a major route to drug resistance. One prediction that might then follow is that tumors that develop resistance to CDK4/6 inhibitors will also prove resistant to inhibition of the major pathway driving CDK4/6 in those cells—in this case, the ER pathway. This appeared to be the case in our cells, although unexpectedly we also observed a loss of ER/PR signaling.
Cells that were chronically exposed to LY5219 exhibited a marked loss in ER and PR mRNA and protein expression. This was observed to occur in models with different mechanisms of resistance to CDK4/6 inhibition potentially implying the basis is related to constitutive bypass of the G1–S checkpoint. We speculate that one potential effector of this response may be the E2F family of transcription factors that would be more persistently liberated from Rb and function as known regulators of ER expression.18, 19 Irrespectively, this loss of ER/PR expression was confirmed to occur in patients receiving these drugs who had biopsies after progression on treatment. Although our numbers of postprogression samples are quite limited, it is uncommon to observe true loss of ER+/PR+ status clinically and thus these individual events portend a real possibility that a subset of the patients who develop resistance to CDK4/6 inhibitors will further manifest with decreases in ER/PR expression. With respect to ER dependence, loss of ER expression in the models was strongly associated with an acquired insensitivity to hormonal agents such as the ER antagonists—fulvestrant and tamoxifen. These findings are consistent with a recently published study32 showing that CDK6 overexpression is associated with resistance to fulvestrant. Our results point to an uncoupling of cell cycle progression from ER signaling occurring, in part, through a loss of ESR1 transcription. This loss of dependence may certainly occur even without changes in expression; however, our models appeared to manifest with both loss of expression and dependence.
The implications for a loss of hormone receptor signaling and dependence are likely to be quite significant. Patients with ER+ breast cancer are routinely given sequential endocrine-based therapy on the premise that the target pathway remains the driver. Patients may be treated with multiple lines of therapy including aromatase inhibitor, selective ER degrader and combination aromatase inhibitor with mTORC1 inhibition (everolimus). The value of such therapies in the context of ER− and ER-independent disease is likely to be low. This finding, if validated in large clinical cohorts, would place a premium on post-therapy biopsies to confirm the ongoing role and presence of the driver pathway. More importantly, alternative targeting approaches could be considered for tumors understood to be at risk for developing such mechanisms of resistance, potentially forestalling the development of an ER− phenotype.
Materials and methods
Reagents
LY2835219 was provided by Eli Lilly and Company (Indianapolis, IN, USA). PD0332991, LEE011, fulvestrant (ICI182 780) and tamoxifen (4-hydroxytamoxifen) were purchased from Selleck Chemicals (Houston, TX, USA). 4-Hydroxytamoxifen was dissolved in ethanol, whereas all other drugs were reconstituted in dimethyl sulfoxide.
Phospho-Rb (Ser780) (D59B7, no. 8180), phospho-Rb (Ser795) (no. 9301), Rb (4H1, no. 9309), cyclin D1 (92G2, no. 2978), CDK6 (DCS83, no. 3136), CDK4 (D9G3E, no. 12790), CDK2 (78B2, no. 2546), E2F1 (no. 3742), cyclin A2 (BF683, no. 4656), cyclin E2 (no. 4132), GAPDH (D16H11, no. 5174) and progesterone receptor A/B (D8Q2J, no. 8757) were all purchased from Cell Signaling Technology (Danvers, MA, USA); Mouse anti-cyclin D1 (M20) and anti-ER (F-10) were obtained from Santa Cruz (Dallas, TX, USA); β-actin was obtained from Sigma-Aldrich (St Louis, MO, USA).
Cell culture and resistant cell line establishment
All breast cancer cell lines used were obtained from the American Type Culture Collection (Manassas, VA, USA) and maintained in a humidified atmosphere with 5% CO2 at 37 °C. T47D cells were cultured in RPMI, MCF-7 in DMEM (Dulbecco's modified Eagle's medium) high glucose and CAMA-1 in DMEM-F12 with non-essential amino acids. All media were supplemented with 10% heat-inactivated fetal bovine serum (Corning, NY, USA), 100 μg/ml streptomycin and 4 mm glutamine. The 293T cell line was obtained from the laboratory of Ping Chi and cultured in DMEM with high glucose.
Drug-resistant cells were established by continuous selection using methods similar to those previously described.33 Briefly, MCF-7, T47D and CAMA-1 cells were cultured in medium containing LY5219 at the concentration of 500 nm (T47D) or 100 nm (MCF-7 and CAMA-1) with fresh media and drug replenished every 3 days. Cells were subcultured every 2–3 weeks with a 25% increase in drug concentration. The resistant cells were established after 4 months (final concentration: T47D 1 μm, MCF-7 200 nm, CAMA-1 100 nm) and maintained in the presence of 100 nm LY5219 and named as MR, TR and CAMA-1 resistant (CR).
Plasmids and generation of stable cell lines
pDONR223-CDK6, pDONR223-CDK4 (a gift from William Hahn and David Root; Addgene (Cambridge, MA, USA); plasmid nos. 23688 and 23778)34 and pDONR223-GFP (a gift from the Yu Chen Lab) was delivered in the MSCV-N-Flag-HA-IRES-PURO plasmid using the Gateway LR Clonase II Enzyme Mix (Invitrogen, Waltham, MA, USA) and named as MSCV-N-CDK6, MSCV-N-CDK4 and MSCV-N-control. Retroviral vector pSIREN-RetroQ-CDK6-shRNA was a gift from Dr Judy Lieberman (Addgene; plasmid no. 25789).35 pSIREN-RetroQ-shN was used as a negative control (a gift from Joe Landry; Addgene; plasmid no. 73665). Retroviral infections were performed as described previously.36
Human tumor specimens
Patients with metastatic ER+ breast cancer were prospectively enrolled on an institutional review board-approved protocol for treatment regimens either including abemaciclib or ribociclib. Among these patients, those who had routine standard of care biopsies of their breast cancer both before and after protocol therapy and agreed (signed informed consent) to have their specimens analyzed for further research were included. Routine analyses of ER/PR immunohistochemistry were performed using Food Drug Administration-approved antibodies and scored by the clinical pathologist.
Cell proliferation
Cells (2.5 × 104/ml) were seeded into 96-well plates in DMEM and treated with different concentrations of indicated drugs. On days 0, 3, 5 and 7, 25 μl of resazurin (R&D Systems, Minneapolis, MN, USA) was added into each well according to the manufacturer’s instructions. After 4 h, fluorescence in the plate was measured using a microplate reader (SpectraMax M5; Molecular Devices, Sunnyvale, CA, USA) with an excitation and emission of 560 and 590 nm. Results were normalized to blank media with no cells. Cell growth data from day 5 was then plotted as the percentage inhibition against the log concentration of indicated drug. IC50 was determined using a sigmoidal regression model using GraphPad Prism 6.0 (GraphPad Software, La Jolla, CA, USA) and was defined as the concentration of drug required for a 50% reduction in growth. Each experiment was repeated at least three times with six replicates.
Immunoblotting
After treatment, cells were washed two times in cold phosphate-buffered saline and collected on ice. Cells were lysed and immunobloted as described previously.36 Equal amounts of total protein (30 μg) was separated by 4–12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis minigel electrophoresis and transferred onto nitrocellulose membranes. Membranes were sequentially probed with indicated primary antibodies and corresponding mouse or rabbit horseradish peroxidase-conjugated secondary antibodies (Sigma-Aldrich). Blots were then developed using enhanced chemiluminescence (Merck Millipore, Billerica, MA, USA).
Quantitative RT–PCR
Taqman primers for CDK4, CDK6, CCND1, CCNE1, CCNE2, CCNA2, E2F1, RB1, ESR1, PGR, GREB1, XBP1, TFF1 and MYC were used to detect mRNA levels by quantitative RT–PCR. Total cellular RNA was extracted from cells using the RNeasy Mini Kit (Qiagen, Venlo, Netherlands) according to the manufacturer's protocol. cDNA was synthesized from 1 μg of RNA with the qScript cDNA SuperMix (Quanta Biosciences, Beverly, MA, USA) and 2 μl of cDNA was added to the Taqman PCR Master Mix (Applied Biosystems, Foster City, CA, USA) along with the appropriate primer. Relative quantification for each mRNA was performed using the comparative CT method with a ViiA 7 Real-Time PCR System (Applied Biosystems). The mRNA expression of each gene was normalized to that of RPLP0. TaqMan primers were all purchased from Applied Biosystems. Each experiment was repeated at least three times with three replicates.
Fluorescence-activated cell sorting
Cells (2 × 104 cells) were treated with drug or dimethyl sulfoxide for 48 h. Both adherent and floating cells were collected. Nuclei were isolated and stained with ethidium bromide. The DNA content (G0/G1, S and G2/M) was determined by flow cytometry as described previously.37
ChIP analysis
ChIP assay was performed as described before.38 Briefly, cells were crosslinked and collected. Chromatin was broken down into 200–1000 bp fragments through 30 min sonication. One microgram of antibody was added into the lysate and incubated overnight at 4 °C. Twenty microliters of ChIP grade Protein A/G magnetic beads were added into each IP tube and incubated for 2–4 h. IP samples were washed and crosslinks reversed by adding proteinase K and incubating overnight at 65 °C. DNA was then recovered using the Qiagen PCR Purification Kit (Qiagen). RNA polymerase II antibody (05-623; EMD Millipore, Billerica, MA, USA) was used in this study. Primer sequences used for RNA polymerase II ChIP were as follows: ESR1 pol, forward: 5′-TTGTGCCTGGAGTGATGTTT -3′ and reverse: 5′-GCATTACAAAGGTGCTGGAG-3′.
Fluorescence in situ hybridization
Cell lines were cultured with colcemid (0.1 μg/ml) and fixed in methanol acetic acid (3:1). FISH analysis was performed on the fixed cells according to standard protocols with a two-color CDK6/Cen7 probe. The probe mix consisted of bacterial artificial chromosome clones containing the full-length CDK6 gene (clones RP11-809H24 and RP11-1102K14; labeled with Red dUTP) and a centromeric repeat plasmid for chromosome 7 served as the control (clone p7t1; labeled with Green dUTP). Slides were scanned and representative regions were selected for imaging. For each cell line at least 100 discrete interphase nuclei and 10 metaphases were analyzed. Amplification was defined as CDK6:Cen7 (control) ratio of ⩾1.2, >6 copies of CDK6 (independent of control locus) or at least one small cluster of CDK6 (⩾4 signals resulting from tandem repeat/duplication).
Statistical methods
Statistical analysis was performed using two-sided standard Student's t-tests and probability values of P<0.05 were regarded as statistically significant. Analysis was conducted using GraphPad Prism v.6.0c (GraphPad Software).
References
Zwijsen RM, Wientjens E, Klompmaker R, van der Sman J, Bernards R, Michalides RJ . CDK-independent activation of estrogen receptor by cyclin D1. Cell 1997; 88: 405–415.
Sicinski P, Donaher JL, Parker SB, Li T, Fazeli A, Gardner H et al. Cyclin D1 provides a link between development and oncogenesis in the retina and breast. Cell 1995; 82: 621–630.
Cariou S, Donovan JC, Flanagan WM, Milic A, Bhattacharya N, Slingerland JM . Down-regulation of p21WAF1/CIP1 or p27Kip1 abrogates antiestrogen-mediated cell cycle arrest in human breast cancer cells. Proc Natl Acad Sci USA 2000; 97: 9042–9046.
O’Sullivan CC . Overcoming endocrine resistance in hormone-receptor positive advanced breast cancer-the emerging role of CDK4/6 inhibitors. Int J Cancer Clin Res 2015; 2: pii:029.
Liang J, Slingerland JM . Multiple roles of the PI3K/PKB (Akt) pathway in cell cycle progression. Cell Cycle 2003; 2: 336–342.
Sherr CJ, Beach D, Shapiro GI . Targeting CDK4 and CDK6: from discovery to therapy. Cancer Discov 2016; 6: 353–367.
Finn RS, Dering J, Conklin D, Kalous O, Cohen DJ, Desai AJ et al. PD 0332991, a selective cyclin D kinase 4/6 inhibitor, preferentially inhibits proliferation of luminal estrogen receptor-positive human breast cancer cell lines in vitro. Breast Cancer Res 2009; 11: R77.
Gelbert LM, Cai S, Lin X, Sanchez-Martinez C, Del Prado M, Lallena MJ et al. Preclinical characterization of the CDK4/6 inhibitor LY2835219: in vivo cell cycle-dependent/independent anti-tumor activities alone/in combination with gemcitabine. Invest New Drugs 2014; 32: 825–837.
Finn RS, Crown JP, Lang I, Boer K, Bondarenko IM, Kulyk SO et al. The cyclin-dependent kinase 4/6 inhibitor palbociclib in combination with letrozole versus letrozole alone as first-line treatment of oestrogen receptor-positive, HER2-negative, advanced breast cancer (PALOMA-1/TRIO-18): a randomised phase 2 study. Lancet Oncol 2015; 16: 25–35.
Patnaik A, Rosen LS, Tolaney SM, Tolcher AW, Goldman JW, Gandhi L et al. (eds). LY2835219, a novel cell cycle inhibitor selective for CDK4/6, in combination with fulvestrant for patients with hormone receptor positive (HR+) metastatic breast cancer. ASCO Annual Meeting Proceedings, 2014.
Cristofanilli M, Turner NC, Bondarenko I, Ro J, Im S-A, Masuda N et al. Fulvestrant plus palbociclib versus fulvestrant plus placebo for treatment of hormone-receptor-positive, HER2-negative metastatic breast cancer that progressed on previous endocrine therapy (PALOMA-3): final analysis of the multicentre, double-blind, phase 3 randomised controlled trial. Lancet Oncol 2016; 17: 425–439.
Dean JL, Thangavel C, McClendon AK, Reed CA, Knudsen ES . Therapeutic CDK4/6 inhibition in breast cancer: key mechanisms of response and failure. Oncogene 2010; 29: 4018–4032.
Dean JL, McClendon AK, Hickey TE, Butler LM, Tilley WD, Witkiewicz AK et al. Therapeutic response to CDK4/6 inhibition in breast cancer defined by ex vivo analyses of human tumors. Cell Cycle 2012; 11: 2756–2761.
Cheng DT, Mitchell TN, Zehir A, Shah RH, Benayed R, Syed A et al. Memorial Sloan Kettering-Integrated Mutation Profiling of Actionable Cancer Targets (MSK-IMPACT): a hybridization capture-based next-generation sequencing clinical assay for solid tumor molecular oncology. J Mol Diagn 2015; 17: 251–264.
VanArsdale T, Boshoff C, Arndt KT, Abraham RT . Molecular pathways: targeting the cyclin D–CDK4/6 axis for cancer treatment. Clin Cancer Res 2015; 21: 2905–2910.
Watts CK, Brady A, Sarcevic B, deFazio A, Musgrove EA, Sutherland RL . Antiestrogen inhibition of cell cycle progression in breast cancer cells in associated with inhibition of cyclin-dependent kinase activity and decreased retinoblastoma protein phosphorylation. Mol Endocrinol 1995; 9: 1804–1813.
Yu Q, Sicinska E, Geng Y, Ahnstrom M, Zagozdzon A, Kong Y et al. Requirement for CDK4 kinase function in breast cancer. Cancer Cell 2006; 9: 23–32.
Macaluso M, Cinti C, Russo G, Russo A, Giordano A . pRb2/p130-E2F4/5-HDAC1-SUV39H1-p300 and pRb2/p130-E2F4/5-HDAC1-SUV39H1-DNMT1 multimolecular complexes mediate the transcription of estrogen receptor-alpha in breast cancer. Oncogene 2003; 22: 3511–3517.
Umemura S, Shirane M, Takekoshi S, Kusakabe T, Itoh J, Egashira N et al. Overexpression of E2F-5 correlates with a pathological basal phenotype and a worse clinical outcome. Br J Cancer 2009; 100: 764–771.
Miller TW, Balko JM, Fox EM, Ghazoui Z, Dunbier A, Anderson H et al. ERalpha-dependent E2F transcription can mediate resistance to estrogen deprivation in human breast cancer. Cancer Discov 2011; 1: 338–351.
Wilcken NR, Prall OW, Musgrove EA, Sutherland RL . Inducible overexpression of cyclin D1 in breast cancer cells reverses the growth-inhibitory effects of antiestrogens. Clin Cancer Res 1997; 3: 849–854.
Karnik PS, Kulkarni S, Liu X-P, Budd GT, Bukowski RM . Estrogen receptor mutations in tamoxifen-resistant breast cancer. Cancer Res 1994; 54: 349–353.
Houston S, Plunkett T, Barnes D, Smith P, Rubens R, Miles D . Overexpression of c-erbB2 is an independent marker of resistance to endocrine therapy in advanced breast cancer. Br J Cancer 1999; 79: 1220.
Jansen MP, Foekens JA, van Staveren IL, Dirkzwager-Kiel MM, Ritstier K, Look MP et al. Molecular classification of tamoxifen-resistant breast carcinomas by gene expression profiling. J Clin Oncol 2005; 23: 732–740.
Schwartz GK, LoRusso PM, Dickson MA, Randolph SS, Shaik MN, Wilner KD et al. Phase I study of PD 0332991, a cyclin-dependent kinase inhibitor, administered in 3-week cycles (Schedule 2/1). Br J Cancer 2011; 104: 1862–1868.
Infante JR, Shapiro G, Witteveen P, Gerecitano JF, Ribrag V, Chugh R et al. (eds). A phase I study of the single-agent CDK4/6 inhibitor LEE011 in pts with advanced solid tumors and lymphomas. ASCO Annual Meeting Proceedings, 2014.
Patnaik A, Rosen LS, Tolaney SM, Tolcher AW, Goldman JW, Gandhi L et al. Efficacy and safety of abemaciclib, an inhibitor of CDK4 and CDK6, for patients with breast cancer, non-small cell lung cancer, and other solid tumors. Cancer Discov 2016; 6: 740–753.
Tigan AS, Bellutti F, Kollmann K, Tebb G, Sexl V . CDK6—a review of the past and a glimpse into the future: from cell-cycle control to transcriptional regulation. Oncogene 2016; 35: 3083–3091.
Grossel MJ, Hinds PW . Beyond the cell cycle: a new role for Cdk6 in differentiation. J Cell Biochem. 2006; 97: 485–493.
Herrera-Abreu MT, Palafox M, Asghar U, Rivas MA, Cutts RJ, Garcia-Murillas I et al. Early adaptation and acquired resistance to CDK4/6 inhibition in estrogen receptor-positive breast cancer. Cancer Res 2016; 76: 2301–2313.
Konecny GE, Winterhoff B, Kolarova T, Qi J, Manivong K, Dering J et al. Expression of p16 and retinoblastoma determines response to CDK4/6 inhibition in ovarian cancer. Clin Cancer Res 2011; 17: 1591–1602.
Alves CL, Elias D, Lyng MB, Bak M, Kirkegaard T, Lykkesfeldt AE et al. High CDK6 protects cells from fulvestrant-mediated apoptosis and is a predictor of resistance to fulvestrant in estrogen receptor-positive metastatic breast cancer. Clin Cancer Res 2016, pii: clinccanres.1984.2015.
McDermott M, Eustace AJ, Busschots S, Breen L, Crown J, Clynes M et al. In vitro development of chemotherapy and targeted therapy drug-resistant cancer cell lines: a practical guide with case studies. Front Oncol 2014; 4: 40.
Johannessen CM, Boehm JS, Kim SY, Thomas SR, Wardwell L, Johnson LA et al. COT drives resistance to RAF inhibition through MAP kinase pathway reactivation. Nature 2010; 468: 968–972.
Navarro F, Gutman D, Meire E, Caceres M, Rigoutsos I, Bentwich Z et al. miR-34a contributes to megakaryocytic differentiation of K562 cells independently of p53. Blood 2009; 114: 2181–2192.
Toy W, Shen Y, Won H, Green B, Sakr RA, Will M et al. ESR1 ligand-binding domain mutations in hormone-resistant breast cancer. Nat Genet 2013; 45: 1439–1445.
She QB, Chandarlapaty S, Ye Q, Lobo J, Haskell KM, Leander KR et al. Breast tumor cells with PI3K mutation or HER2 amplification are selectively addicted to Akt signaling. PLoS One 2008; 3: e3065.
Bosch A, Li Z, Bergamaschi A, Ellis H, Toska E, Prat A et al. PI3K inhibition results in enhanced estrogen receptor function and dependence in hormone receptor-positive breast cancer. Sci Transl Med 2015; 7: 283ra51.
Acknowledgements
This work was generously supported by funds from the Shen Family Foundation, Susan G Komen and the Breast Cancer Research Foundation. The Marie-Josée and Henry R Kravis Center for Molecular Oncology as well as NCI Cancer Center Support Grant (CCSG, P30 CA08748) additionally supported this work. We thank Weiyi Toy for her critical feedback.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
Dr Chandarlapaty has received research support from Novartis and Lilly. The other authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on the Oncogene website
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Yang, C., Li, Z., Bhatt, T. et al. Acquired CDK6 amplification promotes breast cancer resistance to CDK4/6 inhibitors and loss of ER signaling and dependence. Oncogene 36, 2255–2264 (2017). https://doi.org/10.1038/onc.2016.379
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2016.379
This article is cited by
-
Molecular profiling of pediatric and young adult colorectal cancer reveals a distinct genomic landscapes and potential therapeutic avenues
Scientific Reports (2024)
-
The paradox of senescent-marker positive cancer cells: challenges and opportunities
npj Aging (2024)
-
MYC induces CDK4/6 inhibitors resistance by promoting pRB1 degradation
Nature Communications (2024)
-
CDK4/6i-treated HR+/HER2- breast cancer tumors show higher ESR1 mutation prevalence and more altered genomic landscape
npj Breast Cancer (2024)
-
The CDK4/6 inhibitor revolution — a game-changing era for breast cancer treatment
Nature Reviews Clinical Oncology (2024)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}