
Abstract
The TET (ten–eleven translocation) family of α-ketoglutarate (α-KG)-dependent dioxygenases catalyzes the sequential oxidation of 5-methylcytosine (5mC) to 5-hydroxymethylcytosine (5hmC), 5-formylcytosine and 5-carboxylcytosine, leading to eventual DNA demethylation. The TET2 gene is a bona fide tumor suppressor frequently mutated in leukemia, and TET enzyme activity is inhibited in IDH1/2-mutated tumors by the oncometabolite 2-hydroxyglutarate, an antagonist of α-KG, linking 5mC oxidation to cancer development. We report here that the levels of 5hmC are dramatically reduced in human breast, liver, lung, pancreatic and prostate cancers when compared with the matched surrounding normal tissues. Associated with the 5hmC decrease is the substantial reduction of the expression of all three TET genes, revealing a possible mechanism for the reduced 5hmC in cancer cells. The decrease of 5hmC was also observed during tumor development in different genetically engineered mouse models. Together, our results identify 5hmC as a biomarker whose decrease is broadly and tightly associated with tumor development.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 50 print issues and online access
$259.00 per year
only $5.18 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
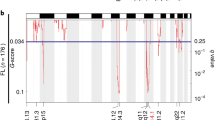
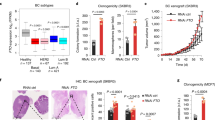
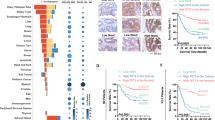
References
Amary MF, Bacsi K, Maggiani F, Damato S, Halai D, Berisha F et al. (2011a). IDH1 and IDH2 mutations are frequent events in central chondrosarcoma and central and periosteal chondromas but not in other mesenchymal tumours. J Pathol 224: 334–343.
Amary MF, Damato S, Halai D, Eskandarpour M, Berisha F, Bonar F et al. (2011b). Ollier disease and Maffucci syndrome are caused by somatic mosaic mutations of IDH1 and IDH2. Nat Genet 43: 1262–1265.
Dang L, White DW, Gross S, Bennett BD, Bittinger MA, Driggers EM et al. (2009). Cancer-associated IDH1 mutations produce 2-hydroxyglutarate. Nature 462: 739–744.
Delhommeau F, Dupont S, Della VV, James C, Trannoy S, Masse A et al. (2009). Mutation in TET2 in myeloid cancers. N Engl J Med 360: 2289–2301.
Figueroa ME, Abdel-Wahab O, Lu C, Ward PS, Patel J, Shih A et al. (2010). Leukemic IDH1 and IDH2 mutations result in a hypermethylation phenotype, disrupt TET2 function, and impair hematopoietic differentiation. Cancer Cell 18: 553–567.
Haffner MC, Chaux A, Meeker AK, Esopi DM, Gerber J, Pellakuru LG et al. (2011). Global 5-hydroxymethylcytosine content is significantly reduced in tissue stem/progenitor cell compartments and in human cancers. Oncotarget 2: 627–637.
He YF, Li BZ, Li Z, Liu P, Wang Y, Tang Q et al. (2011). Tet-mediated formation of 5-carboxylcytosine and its excision by TDG in mammalian DNA. Science 333: 1303–1307.
Hemerly JP, Bastos AU, Cerutti JM . (2010). Identification of several novel non-p.R132 IDH1 variants in thyroid carcinomas. Eur J Endocrinol 163: 747–755.
Ito S, D'Alessio AC, Taranova OV, Hong K, Sowers LC, Zhang Y . (2010). Role of tet proteins in 5mC to 5hmC conversion, ES-cell self-renewal and inner cell mass specification. Nature 466: 1129–1133.
Ito S, Shen L, Dai Q, Wu SC, Collins LB, Swenberg JA et al. (2011). Tet proteins can convert 5-methylcytosine to 5-formylcytosine and 5-carboxylcytosine. Science 333: 1300–1303.
Jones PA, Baylin SB . (2007). The epigenomics of cancer. Cell 128: 683–692.
Langemeijer SM, Kuiper RP, Berends M, Knops R, Aslanyan MG, Massop M et al. (2009). Acquired mutations in TET2 are common in myelodysplastic syndromes. Nat Genet 41: 838–842.
Mardis ER, Ding L, Dooling DJ, Larson DE, McLellan MD, Chen K et al. (2009). Recurring mutations found by sequencing an acute myeloid leukemia genome. N Engl J Med 361: 1058–1066.
Mullighan CG . (2009). TET2 mutations in myelodysplasia and myeloid malignancies. Nat Genet 41: 766–767.
Murugan AK, Bojdani E, Xing M . (2010). Identification and functional characterization of isocitrate dehydrogenase 1 (IDH1) mutations in thyroid cancer. Biochem Biophys Res Commun 393: 555–559.
Noushmehr H, Weisenberger DJ, Diefes K, Phillips HS, Pujara K, Berman BP et al. (2010). Identification of a CpG island methylator phenotype that defines a distinct subgroup of glioma. Cancer Cell 17: 510–522.
Pansuriya TC, van Eijk R, d'Adamo P, van Ruler MA, Kuijjer ML, Oosting J et al. (2011). Somatic mosaic IDH1 and IDH2 mutations are associated with enchondroma and spindle cell hemangioma in Ollier disease and Maffucci syndrome. Nat Genet 43: 1256–1261.
Parsons DW, Jones S, Zhang X, Lin JC, Leary RJ, Angenendt P et al. (2008). An integrated genomic analysis of human glioblastoma multiforme. Science 321: 1807–1812.
Tahiliani M, Koh KP, Shen Y, Pastor WA, Bandukwala H, Brudno Y et al. (2009). Conversion of 5-methylcytosine to 5-hydroxymethylcytosine in mammalian DNA by MLL partner TET1. Science 324: 930–935.
Wu SC, Zhang Y . (2010). Active DNA demethylation: many roads lead to Rome. Nat Rev Mol Cell Biol 11: 607–620.
Xu W, Yang H, Liu Y, Yang Y, Wang P, Kim SH et al. (2011). Oncometabolite 2-hydroxyglutarate is a competitive inhibitor of alpha-ketoglutarate-dependent dioxygenases. Cancer Cell 19: 17–30.
Zhao S, Lin Y, Xu W, Jiang W, Zha Z, Wang P et al. (2009). Glioma-derived mutations in IDH1 dominantly inhibit IDH1 catalytic activity and induce HIF-1alpha. Science 324: 261–265.
Acknowledgements
We thank the members of the Fudan MCB laboratory for discussions and support throughout this study, and Eric Oermann for reading the manuscript. This work was supported by MOST 973 (No. 2009CB918401, No. 2011CB910600), NSFC (Grant No. 30600112, 30871255, 31071192). This work was also supported by the 985 Program, Shanghai key project (Grant No. 09JC1402300), the Shanghai Leading Academic Discipline Project (project number B110), and NIH grants (to YX and KLG).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on the Oncogene website
Supplementary information
Rights and permissions
About this article
Cite this article
Yang, H., Liu, Y., Bai, F. et al. Tumor development is associated with decrease of TET gene expression and 5-methylcytosine hydroxylation. Oncogene 32, 663–669 (2013). https://doi.org/10.1038/onc.2012.67
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/onc.2012.67
Keywords
This article is cited by
-
Transcending frontiers in prostate cancer: the role of oncometabolites on epigenetic regulation, CSCs, and tumor microenvironment to identify new therapeutic strategies
Cell Communication and Signaling (2024)
-
TET2 Is Downregulated in Early Esophageal Squamous Cell Carcinoma and Promotes Esophageal Squamous Cell Malignant Behaviors
Digestive Diseases and Sciences (2024)
-
Diagnosing and monitoring pancreatic cancer through cell-free DNA methylation: progress and prospects
Biomarker Research (2023)
-
StableDNAm: towards a stable and efficient model for predicting DNA methylation based on adaptive feature correction learning
BMC Genomics (2023)
-
Loss of 5-hydroxymethylcytosine induces chemotherapy resistance in hepatocellular carcinoma via the 5-hmC/PCAF/AKT axis
Cell Death & Disease (2023)
