
Abstract
Objective:
Yacon is a perennial plant forming a clump of >20 big, edible underground tubers. Yacon, which originates from South America, has become increasingly popular in the Japanese diet for tubers have a lower caloric value and a high fiber content. Recent studies have suggested that yacon feeding ameliorates diabetes as indicated by reduced blood glucose.
Methods:
We fed male Zucker fa/fa rats for 5 weeks with isocaloric normal chow diet containing from 6.5% control aroid or 6.5% yacon. Insulin sensitivity was evaluated by euglycemic-hyperinsulinemic clamp study.
Results:
Body weight was comparable between yacon- and aroid-fed rats. In the basal state, yacon feeding had an effect to lower fasting glucose levels from 184.1±4.1 to 167.8±2.7 mg dl−1 (P<0.01), as well as basal hepatic glucose output (HGO) from 9.9±0.4 to 7.4 ± 0.2 mg kg−1 per min (P<0.01). During the clamp studies, the glucose infusion rate required to maintain euglycemia was increased by 12.3% in yacon-fed rat. The insulin suppression of HGO was also increased in yacon-fed rats compared with control rats (85.3±2.4% vs 77.0±3.0%; P<0.05), whereas the glucose disposal rate was not different between the two groups. Consistent with the clamp data, the insulin-stimulated phosphorylation of Akt was significantly enhanced in liver but not in skeletal muscle. Furthermore, tribbles 3 (Trb3) expression, which is a negative regulator of Akt activity, was markedly reduced in the liver of yacon-fed rats compared with control rats.
Conclusion:
These results indicate that the effect of yacon feeding to reduce blood glucose is likely due to its beneficial effects on hepatic insulin sensitivity in the insulin resistant state.
Similar content being viewed by others

Introduction
Type 2 diabetes mellitus is characterized by insulin resistance and β-cell dysfunction, the balance between which varies widely between individuals. Insulin resistance, which is characterized by an impaired ability of insulin to inhibit glucose output from the liver and to promote glucose uptake in muscle,1 is frequently well established in individuals with impaired glucose intolerance. It is the concomitant presence of β-cell dysfunction that distinguishes those in whom glucose intolerance worsens from those in whom it remains stable. Interventions that decrease insulin resistance and preserve or improve β-cell function are likely to be effective in slowing progression from impaired glucose intolerance to diabetes, or even allowing reversion to normal glucose tolerance. Many interventions that improve insulin resistance might also affect insulin secretion. The Diabetes Prevention Program, a multicenter, randomized placebo-controlled, trial, examined the effect of two interventions to prevent or delay type 2 diabetes mellitus in people at high risk. This study found that the risk for developing type 2 diabetes was reduced by 58% and 31% in intensive lifestyle and metformin-treated groups, respectively, compared with the placebo-treated group.2 Intensive lifestyle intervention was more effective than metformin in slowing progression to diabetes, partly because lifestyle modification gave greater improvements in insulin sensitivity and β-cell functions.3 Changes in physical activity and diet (primarily, a reduced calorie intake from fat) predicted weight loss and weight loss, in turn, was associated with a reduced risk of developing type 2 diabetes mellitus. Thus, dietary interventions can be an effective tool to prevent or treat insulin resistance and type 2 diabetes mellitus.2
Yacon (Smallanthus sonchifolius, Asteraceae) is a perennial plant originating from South America that forms a clump of >20 big sweet-tasting underground tubers weighing from 100–500 g.4 In recent decades, yacon has gained increasing popularity in Japan and around the world for its low caloric value. Yacon tubers are composed mostly of water and very high levels of fructo-oligosaccharides (FOS) also called oligofructose or oligofructans. FOS are a type of sugar found naturally in many types of plants but unlike other oligosaccharides, they are able to resist the hydrolysis of enzymes in the saliva and upper gastrointestinal tract and leaves the body undigested. Thus, despite their sweet flavor, yacon tubers contain fewer calories than expected.4 Traditionally, yacon tubers have been recommended to people suffering from diabetes and various digestive diseases.5, 6, 7 Recently, yacon syrup, which is extracted and concentrated from the yacon tubers, has been shown to improve insulin resistance and reduce body weight in obese individuals.7 These studies raise the possibility that yacon feeding has beneficial effects in treating obesity-related insulin resistance and type 2 diabetes mellitus. However, they did not establish whether the effects of yacon on insulin sensitivity in obese individuals were primary or secondary to the decrease in body weight.
In the current study, we examined the metabolic effects of yacon-enriched diet in insulin resistant Zucker fa/fa rats. We show that yacon feeding effectively normalized hyperglycemia by lowing hepatic glucose production and increasing whole-body insulin sensitivity in these animals. Our data provide a physiological mechanism for how yacon dietary supplementation can improve diabetes in humans.
Materials and methods
Materials
Male Zucker fa/fa rats were procured from Charles River Laboratories Inc. (Kanagawa, Japan). Yacon and aroid were kindly provided from Fukushima Prefectural government. The catheter (Micro-Renathane MRE-033, 0.033 cm in outside diameter (OD) and 0.014 cm in inside diameter (ID)) was purchased from Braintree Scientific (Braintree, MA, USA). D-[3-3H] glucose was purchased from PerkinElmer Inc. (Waltham, MA, USA). Pentobarbital was purchased from Dainippon Sumitomo Pharma (Osaka, Japan). Insulin (Novolin R) was purchased from Novo Nordisk (Copenhagen, Denmark). The 50% dextrose was purchased from Otsuka Pharmaceutical Co. (Tokushima, Japan). The mouse/rat adiponectin enzyme-linked immunosorbent assay kit was purchased from B-Bridge International Inc. (San Jose, CA, USA). The rat insulin enzyme-linked immunosorbent assay kit was purchased from Crystal Chem Inc. (Chicago, IL, USA). Polyvinylidine difluoride transfer membranes were purchased from Millipore Corp. (Bedford, MA, USA). Anti-phospho-specific Akt (Ser473) and Akt antibodies were purchased from Cell Signaling Technology (Boston, MA, USA). Horseradish peroxidase-conjugated secondary antibody was purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). TRIzol reagent was purchased from Invitrogen Life Technologies (Carlsbad, CA, USA). The RNeasy kit was purchased from Qiagen Inc. (Valencia, CA, USA). iScript complementary DNA Synthesis Kit and iQ SYBR Green Supermix were purchased from Bio-Rad Laboratories (Richmond, CA, USA). All other reagents were purchased from Sigma (St. Louis, MO, USA).
Animal studies
Six-week-old male Zucker fa/fa rats (Charles River Laboratory) were selected for this study, because they display pronounced insulin resistance at this age. Rats were housed individually under controlled light/dark (12/12 h) and temperature conditions, and had free access to water and chow. The male Zucker fa/fa rats were fed an isocaloric normal chow diet containing from 6.5% aroid (control) or 6.5% yacon for 5 weeks. Rat received a fresh diet every 3 days, and food consumption rates and body weight gains were monitored every 3 days. All procedures were performed in accordance with the Guide for Care and Use of Laboratory Animals of the NIH and were approved by the Animal Subjects Committee of the Fukushima Medical University, Japan.
Euglycemic-hyperinsulinemic clamp procedure
The rats were implanted with three catheters (Micro-Renathane MRE-033, 0.033 cm in OD and 0.014 cm in ID; Braintree Scientific) after feeding the indicated chow for 4 weeks. Two catheters were placed into the right jugular vein, and another one was placed into the left carotid artery under single-dose anesthesia that pentobarbital 50 mg kg−1 (Dainippon Sumitomo Pharma) given intraperitoneally. Catheters were tunneled subcutaneously, exteriorized at the back of the neck and filled with heparinized saline. The jugular and carotid catheters were used for infusion and blood sampling, respectively.
On day 7 after surgery, insulin sensitivity was assessed using an euglycemic-hyperinsulinemic clamp. The rats were fasted for 12 h before the start of all experiments. The euglycemic-hyperinsulinemic clamp experiments began with a priming injection (2.5 μCi per 0.5 ml) and constant infusion (0.04 μCi per minute) of D-[3-3H] glucose (PerkinElmer Inc). After 120 min of tracer equilibration and basal sampling at time −10 min, and 0 min, glucose (50% dextrose, variable infusion; Otsuka Pharmaceutical Co.) and tracer (0.12 μCi min−1) plus insulin (25 mU kg min−1, Novolin R; Novo Nordisk) were infused into the jugular vein as previously described.8, 9
Small blood samples (60 μl) were drawn at 10-min intervals and immediately analyzed for glucose (Compact Electrode Blood Sugar Analyzer Antsense: Horiba Ltd, Kyoto, Japan) to maintain the integrity of the glucose clamp throughout the duration of the experiment. Blood samples were taken at −120 min (start of experiment), −10 min and 0 min (basal), and 110 min and 120 min (end of experiment), for determination of glucose-specific activity, and insulin content. To ensure accuracy, basal and terminal sampling was performed twice, at a 10-min interval. We confirmed that steady-state conditions were achieved at the end of the clamp before obtaining the terminal blood specimen by measuring blood glucose every 10 min and assuring that a steady state for glucose infusion and plasma glucose levels was maintained for a minimum of 20 min before final sampling. We define a steady-state blood glucose concentration as one where the glucose concentration and infusion rate fluctuate by 3 mg dl−1 or less and by 5% or less, respectively, over 10 min. All blood samples were immediately centrifuged, and plasma was stored at −80 °C for subsequent analysis. After terminal blood sampling at 120 min, animals were promptly euthanized with pentobarbital (180 mg kg−1). Tissues were taken and immediately frozen in liquid nitrogen and stored at −80 °C for subsequent metabolic analysis.
Analytical procedures
Total cholesterol, triglycerides and high-density lipoprotein-cholesterol were analyzed by a private laboratory (SRL Laboratory, Tokyo, Japan). Plasma adiponectin levels were determined using the Mouse/Rat Adiponectin enzyme-linked immunosorbent assay kit (B-Bridge International Inc.). Basal plasma insulin was quantified using a rat insulin enzyme-linked immunosorbent assay kit (Crystal Chem Inc.). Plasma glucose-specific activity was measured in duplicate after zinc sulfate and barium hydroxide deproteinization.
Immunoblotting analysis
Red quadriceps muscle and liver tissue were homogenized in liquid nitrogen and lyzed in buffer containing phosphatase and protease inhibitors. After a 10-min incubation, the lysates were clarified by centrifugation (10 000 g at 4 °C) and quantified for total protein. Samples were separated by SDS-polyacrylamide gel electrophoresis on 10% polyacrylamide gels. Proteins were transferred onto polyvinylidine difluoride membranes (Immobilon-P; Millipore) and blotted with Akt and phospho-Akt (Ser473) (Cell Signaling Technology) antibodies. Membranes were then incubated with horseradish peroxidase-conjugated secondary antibodies before chemiluminescence detection (Pierce, Rockford, IL, USA). Band intensities were quantified by densitometry using the Image-J software (NIH, Bethesda, MD, USA).
Quantitative real-time reverse-transcription PCR analysis
Total RNA samples were extracted from the liver tissues and the skeletal muscle with TRIzol reagent (Invitrogen Life Technologies), and further purified using the RNeasy kit with RNase-free DNase I treatment according to the manufacturer's instructions. Total RNA (1 μg) was reverse-transcribed with iScript complementary DNA Synthesis Kit according to the manufacturer’s instructions (Bio-Rad Laboratories). Quantitative real-time (qRT)-PCR was performed with a Bio-Rad system using iQ SYBR Green Supermix and specific primer pairs (Table 1) selected with Primer Express software (Applied Biosystems). The relative mass of specific RNAs was calculated by the comparative cycle of threshold detection method according to the manufacturer’s instructions.
Calculations
The homeostasis model assessment insulin resistance values were calculated from the fasting concentrations of insulin and glucose using the following formula: fasting plasma insulin (ng ml−1) × fasting plasma glucose (mg dl−1)/405.
Hepatic glucose output (HGO) and glucose disposal rate (GDR) were calculated for the basal period and steady-state portion of the glucose clamp using the Steele equation for steady-state conditions.10
Data calculation and statistical analysis were performed using the Stat View program (Abacus Concepts Inc., Berkley, CA, USA). Data are presented as mean±s.e.m.. Statistical differences between the two groups were calculated by the unpaired Student's test. All data are distributed to ensure that the use of means and t-test is appropriate. Statistical significance was defined as P<0.05.
Results
Overall animal characteristics
The effect of a yacon-enriched diet was examined in male Zucker fa/fa rat. Male Zucker fa/fa rats were fed isocaloric normal chow diet containing from 6.5% aroid (control) or 6.5% yacon for 5 weeks. Table 2 illustrates some of the general characteristics of the yacon and control groups in the basal state and at 5 weeks. Body weight and average daily food intake were not different between the two groups. In the fasting state, plasma glucose and insulin levels were significantly lower (P<0.01) in the yacon group than in the control group, whereas total cholesterol, triglycerides, high-density lipoprotein-cholesterol and adiponectin levels did not differ significantly between the two groups. Homeostasis model assessment insulin resistance, which evaluates the insulin sensitivity, was significantly decreased (P<0.01) in the yacon group compared with the control group, suggesting an improvement of insulin sensitivity after 5 weeks of yacon feeding.
Euglycemic-hyperinsulinemic clamp studies
To directly examine the metabolic impact of yacon feeding on insulin sensitivity, we next subjected both groups of rats to euglycemic-hyperinsulinemic clamps. These experiments were performed after 5 weeks of yacon feeding, at maximal (25 mU kg−1 per minute) insulin infusion rates. Steady-state glucose levels during the clamp studies were similar in the two groups, as shown in Table 2. During these studies, we measured the insulin stimulation of total body GDR and suppression of HGO. The glucose infusion rate required to maintain euglycemia was significantly increased by 12.3% (P<0.01) in yacon group, showing enhanced overall insulin sensitivity Table 2. To assess the insulin-stimulated component of glucose disposal, the GDR was measured. As seen in Figure 1b, the GDR was not significantly different between the two groups. As skeletal muscle accounts for the great majority of GDR, these results indicate that yacon feeding does not affect insulin sensitivity in skeletal muscle.

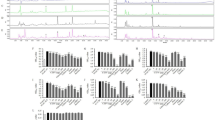
Effect of yacon diet on insulin sensitivity during euglycemic-hyperinsulinemic clamp studies in male Zucker fa/fa rats. (a) Data represent the basal HGO in rat fed with control (□; n=8) or yacon (▪; n=8) for 5 weeks. (b) Data represent the GDR during euglycemic-hyperinsulinemic clamp studies in rat fed with control (□; n=8) or yacon (▪; n=8) for 5 weeks. (c) Data represent the suppression of HGO during euglycemic-hyperinsulinemic clamp studies in rat fed with control (□; n=8) or yacon (▪; n=8) for 5 weeks. Values are means±s.e.m. of data obtained from the analysis of the rats fed with control or yacon. *: P<0.01; **: P<0.05 vs control rats.
Basal HGO values were significantly decreased by 25% (P<0.01) in the yacon group compared with the control group (Figure 1a). During the clamp studies, the insulin suppression of HGO is increased in yacon feeding rats compared with control rats (Figure 1c; 85.3±2.4% vs 77.0±3.0%; P<0.05), suggesting that yacon feeding improves hepatic insulin resistance.
Insulin signaling studies
To assess the potential cellular mechanisms of yacon feeding-induced increase in insulin sensitivity, we obtained skeletal muscle and liver tissue samples in the basal state and at the end of the euglycemic-hyperinsulinemic clamp. These latter samples, which represent the fully insulinized state at the termination of the glucose clamp study, were homogenized, and protein lysates were subjected to immunoblotting.
We measured Akt phosphorylation (Ser473), which is the most important molecule of the insulin signaling, in skeletal muscle and liver samples obtained from rats in the basal state and at the end of the euglycemic-hyperinsulinemic experiments. As shown in Figure 2, insulin led to a marked stimulation of Akt phosphorylation in skeletal muscle and liver from control rats. This effect of Akt phosphorylation at Ser473 was augmented by 50% (P<0.01) in liver, but not in skeletal muscle of the yacon feeding rats.

Effect of yacon diet on Akt phosphorylation (Ser473) in skeletal muscle (a) and liver (b) samples. Basal and insulin clamp-stimulated skeletal muscle and liver samples of control group (□; basal: n=8, clamp: n=8) and yacon group (▪; basal: n=8, clamp: n=8) were obtained and homogenized as described in Methods. After fraction of the resulting homogenates by 10% SDS-polyacrylamide gel electrophoresis, proteins were transferred onto polyvinylidine difluoride membranes, Membranes were probed with phosphorylated Akt (Ser473) and Akt antibodies. The bar graphs show data quantification by NIH-image for results in skeletal muscle and liver. Values are means±s.e.m. of data obtained from the analysis of the rats fed with control or yacon. Data are expressed relative to basal control values. *: P<0.01 vs control rats.
Gluconeogenetic gene studies in liver
To assess the molecular mechanisms of yacon feeding-induced increase in insulin sensitivity, we performed qRT-PCR analysis on total RNA from liver tissue samples at the end of the euglycemic-hyperinsulinemic clamp. We measured the expression levels of phosphoenolpyruvate carboxykinase 1 and glucose-6-phophatase, which are key enzymes in gluconeogenetic pathway. Consistent with the decreased HGO, the expression levels of gluconeogenetic genes, phosphoenolpyruvate carboxykinase 1 and glucose-6-phophatase, were significantly decreased by 49% and 64%, respectively (Figures 3a and b).

Effect of yacon diet on gluconeogenesis gene (phosphoenolpyruvate carboxykinase 1 (Pepck) (a) and glucose-6-phophatase (G6pase) (b)) genes expression during euglycemic-hyperinsulinemic clamp studies in liver samples. Total RNAs extracted from liver tissues (control group (□; clamp: n=8) and yacon group (▪; clamp: n=8)) were used for gene expression analysis of Pepck (a) and G6pase (b). Levels of Cph were used for normalization of sample loading. Values are means±s.e.m. of data obtained from the analysis of the rats fed with control or yacon. Data are expressed relative to control values. *: P<0.01; vs control rats.
Furthermore, based on findings that the hepatic tribbles 3 (TRB3) protein is elevated in insulin resistance models11, 12 and that TRB3 contributes to insulin resistance by blocking insulin signaling by inhibiting Akt activation,11 we hypothesized that yacon feeding could regulate TRB3 expression in liver. We performed qRT-PCR analysis on total RNA from liver and skeletal muscle samples at the end of the euglycemic-hyperinsulinemic clamp. As shown in Figure 4a, Trb3 expression was markedly reduced by 43% (P<0.01) in the liver of yacon-fed rats compared with that of control rats, whereas Trb3 expression in skeletal muscle did not change significantly in either group. Furthermore, we measured the expression of Pgc-1α, which is upstream of TRB3 in liver in the signaling cascade.12 As shown in Figure 4b, Pgc-1α expression in either liver nor skeletal muscle was not altered by yacon feeding.

Effect of yacon diet on Trb3 (a) and Pgc-1α (b) genes expression during euglycemic-hyperinsulinemic clamp studies in liver samples. Total RNAs extracted from liver tissues and skeletal muscle of control group (□; clamp: n=8) and yacon group (▪; clamp: n=8) were used for gene expression analysis of Trb3 (a) and Pgc-1α (b). Levels of Cph were used for normalization of sample loading. Values are means±s.e.m. of data obtained from the analysis of the rats fed with control or yacon. Data are expressed relative to control values. *: P<0.01; vs control rats.
Discussion
Over the past few decades, yacon tubers and yacon products such as yacon syrup and tea have been recommended to dieters and people suffering from diabetes and various digestive diseases for their low-calorie, nutritional and prebiotic value. Recent studies have demonstrated that yacon syrup consumption leads to weight loss and improved insulin sensitivity in obese individuals.7 Whether the beneficial effect on insulin sensitivity is a result of weight reduction or an independent effect of yacon supplementation has not been clarified. In the present studies, using a rat model, we find that yacon feeding leads to a reduction in fasting glucose and insulin levels. This effect is independent of body weight as body weight was not different between the control and yacon groups after 5 weeks of feeding. We assessed the physiological mechanisms underlying this glucose-lowering effect and found that a yacon-enriched diet leads to insulin sensitization exerted predominantly at the level of the liver.
Euglycemic-hyperinsulinemic clamp studies were performed in yacon and aroid-fed insulin-resistant male Zucker fa/fa rats. Yacon feeding led to an improvement in insulin sensitivity in the Zucker fa/fa rats, as demonstrated by a 12.3% increase in the glucose infusion rate necessary to maintain euglycemia in these animals. Using tracer methodologies, we measured the ability of insulin to suppress HGO. Compared with control animals, yacon-fed Zucker fa/fa rats displayed enhanced insulin-mediated suppression of HGO, indicative of increased hepatic insulin sensitivity. We found a comparable effect on GDR for both groups. Together, these data show that yacon feeding leads to an improvement in hepatic insulin resistance but had no effect to ameliorate skeletal muscle insulin resistance.
Yacon FOS, which are soluble non-digestible carbohydrates, effectively increase stool bulk. FOS are classified as prebiotics because they are fermented by the microflora in the large intestine, leading to a modulation in the composition of natural ecosystem. Yacon FOS are reported to have the potential to be fermented by bifidobacteria and lactobacilli, making yacon roots a novel source of prebiotics.13 Probiotics are foods that contain microorganisms that modulate the intestinal microbiota and aid the functioning of the gastrointestinal tract, and thus possibly prevent disease occurrence. Meanwhile, prebiotics are food containing substances resistant to enzymatic breakdown, which stimulate the proliferation or activity of certain bacteria in the intestinal microbiota, thus acting as selective substrate in the colon. Foods that contain both probiotics and prebiotics are called synbiotics.14 Within this context, fermented foods containing probiotics and prebiotics can be important diet components, because of their nutritional characteristics and ability to reduce the risk of chronic inflammation diseases such as insulin resistance.15 More recently, it has been reported that yacon can prevent enteric infection by improving the immunological intestinal barrier.16 Increased levels of circulating bacteria or bacterial products, which are derived from the microbiota, have been associated with insulin resistance.17
Moreover, our cellular measurements shed light on potential mechanisms of action of yacon supplementation and suggest the involvement of TRB3. TRB3 is a pesudokinase that modulates several signaling pathways, including the phosphatidylinositol 3-kinase/Akt cascade. TRB3 is induced in the liver during fasting and in diabetic mice, where it inhibits insulin signaling by binding to and preventing activation of Akt.11, 12 The serine-threonine kinase Akt is a principal target of insulin signaling that inhibits HGO when glucose is available from food. Therefore, pathological overexpression of TRB3 in the fed state contributes to insulin resistance and promotes hyperglycemia. Consistent with our clamp data, we found a uniform increase in Akt phosphorylation and decrease in Trb3 expression in the liver of yacon-fed rats compared with controls. Furthermore, peroxisome proliferator-activated receptor-γ coactivator-1α, a coactivator induced by fasting, glucagon, glucocorticoids and adrenergic stimuli that results in activation of gluconeogenic enzymes and increased HGO, has been shown to be an upstream modulator of Trb3 expression in the liver, an effect mediated by peroxisome proliferator-activated receptor-α.12 We examined the peroxisome proliferator-activated receptor-γ coactivator-1α expression in liver and found that yacon feeding did not alter its expression.
We observed in that feeding a yacon-enriched diet for 5-weeks did not decrease body weight in Zucker fa/fa rats. This enabled us to conclude that yacon feeding effect on blood glucose and insulin sensitivity was independent of weight loss. However, this is not to say that yacon dietary supplementation cannot decrease body weight in rats in the long-term, as documented in humans.7 Although one cannot extrapolate or compare rat studies to human studies, several points could account for the apparent difference: the length of the dietary intervention (5 weeks in rats vs 4 months in human); the gender and age of study population (young male rats vs pre-menopausal women); the type and amount of yacon supplementation (diet with 6.5% of yacon incorporated in chow preparation for rats vs 0.14 g kg−1 body weight per day of yacon syrup).
In conclusion, we show that in insulin resistant rats, a yacon-enriched diet improves insulin resistance in liver that is associated with reduced hepatic Trb3 expression and increased Akt activation. We thus provide a novel mechanistic explanation for the reported glucose-lowering beneficial effects of yacon tubers and yacon products in individuals with diabetes.
References
Olefsky JM, Glass CK . Macrophages, inflammation, and insulin resistance. Annu Rev Physiol 2010; 72: 219–246.
Knowler WC, Barrett-Connor E, Fowler SE, Hamman RF, Lachin JM, Walker EA et al. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N Engl J Med 2002; 346: 393–403.
Kitabchi AE, Temprosa M, Knowler WC, Kahn SE, Fowler SE, Haffner SM et al. Role of insulin secretion and sensitivity in the evolution of type 2 diabetes in the diabetes prevention program: effects of lifestyle intervention and metformin. Diabetes 2005; 54: 2404–2414.
Valentova K, Ulrichova J . Smallanthus sonchifolius and Lepidium meyenii - prospective Andean crops for the prevention of chronic diseases. Biomed Pap Med Fac Univ Palacky Olomouc Czech Repub 2003; 147: 119–130.
Yan X, Suzuki M, Ohnishi-Kameyama M, Sada Y, Nakanishi T, Nagata T . Extraction and identification of antioxidants in the roots of yacon (Smallanthus sonchifolius). J Agric Food Chem 1999; 47: 4711–4713.
Geyer M, Manrique I, Degen L, Beglinger C . Effect of yacon (Smallanthus sonchifolius) on colonic transit time in healthy volunteers. Digestion 2008; 78: 30–33.
Genta S, Cabrera W, Habib N, Pons J, Carillo IM, Grau A et al. Yacon syrup: beneficial effects on obesity and insulin resistance in humans. Clin Nutr 2009; 28: 182–187.
Satoh H, Nguyen MTA, Miles PDG, Imamura T, Usui I, Olefsky JM . Adenovirus-mediated chronic "hyper-resistinemia" leads to in vivo insulin resistance in normal rats. J Clin Invest 2004; 114: 224–231.
Satoh H, Nguyen MT, Trujillo M, Imamura T, Usui I, Scherer PE et al. Adenovirus-mediated adiponectin expression augments skeletal muscle insulin sensitivity in male Wistar rats. Diabetes 2005; 54: 1304–1313.
Steele R . Influences of glucose loading and of injected insulin on hepatic glucose output. Ann N Y Acad Sci 1959; 82: 420–430.
Du K, Herzig S, Kulkarni RN, Montminy M . TRB3: a tribbles homolog that inhibits Akt/PKB activation by insulin in liver. Science 2003; 300: 1574–1577.
Koo SH, Satoh H, Herzig S, Lee CH, Hedrick S, Kulkarni R et al. PGC-1 promotes insulin resistance in liver through PPAR-alpha-dependent induction of TRB-3. Nat Med 2004; 10: 530–534.
Pedreschi R, Campos D, Noratto G, Chirinos R, Cisneros-Zevallos L . Andean yacon root (Smallanthus sonchifolius Poepp. Endl) fructooligosaccharides as a potential novel source of prebiotics. J Agric Food Chem 2003; 51: 5278–5284.
Bengmark S, Martindale R . Prebiotics and synbiotics in clinical medicine. Nutr Clin Pract 2005; 20: 244–261.
Cavallini DC, Suzuki JY, Abdalla DS, Vendramini RC, Pauly-Silveira ND, Roselino MN et al. Influence of a probiotic soy product on fecal microbiota and its association with cardiovascular risk factors in an animal model. Lipids Health Dis 2011; 10: 126.
Velez E, Castillo N, Meson O, Grau A, Bibas Bonet ME, Perdigon G . Study of the effect exerted by fructo-oligosaccharides from yacon (Smallanthus sonchifolius) root flour in an intestinal infection model with Salmonella Typhimurium. Br J Nutr 2012. e-pub ahead of print 9 Nov 2012; doi.org/10.1017/S0007114512004230.
Burcelin R, Garidou L, Pomie C . Immuno-microbiota cross and talk: the new paradigm of metabolic diseases. Semin Immunol 2012; 24: 67–74.
Acknowledgements
We thank Fukushima prefectural government for kindly providing the powder of yacon and aroid, and Hiroko Ohashi and Atsuko Hashimoto for their excellent technical assistance. This work was supported in part by the Salt Science Research Foundation, grant no. 08C4 (HS), a Kanae Foundation for Life & Soci-Medical Science grant (HS), a Takeda Science Foundation grant (HS), a Grant-in-Aid for Challenging Exploratory Research, and a Grant-in-Aid for Scientific Research (C) from the Ministry of Education, Culture, Sports, Science and Technology of Japan (HS), and Banyu Life Science Foundation International (TW).
Author contributions
HS conducted the experiments, and analysis of the data; AK assisted the collection and analysis of qRT-PCR data; HS designed the experiments and wrote the manuscript; and NM and TW contributed to manuscript preparation. All authors have contributed to the final approval of the version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/3.0/
About this article
Cite this article
Satoh, H., Audrey Nguyen, M., Kudoh, A. et al. Yacon diet (Smallanthus sonchifolius, Asteraceae) improves hepatic insulin resistance via reducing Trb3 expression in Zucker fa/fa rats. Nutr & Diabetes 3, e70 (2013). https://doi.org/10.1038/nutd.2013.11
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nutd.2013.11
Keywords
This article is cited by
-
Daily Intake of Smallanthus sonchifolius (Yacon) Roots Reduces the Progression of Non-alcoholic Fatty Liver in Rats Fed a High Fructose Diet
Plant Foods for Human Nutrition (2022)
