
Abstract
Objective:
Experimental endotoxaemia induces subcutaneous adipose tissue inflammation and systemic insulin resistance in lean subjects. Glyceroneogenesis, by limiting free fatty acids (FFA) release from adipocytes, controls FFA homoeostasis and systemic insulin sensitivity. The roles of subcutaneous adipose tissue (SAT) and visceral adipose tissue (VAT) in metabolic deregulation are intrinsically different. We compared the effect of lipopolysaccharide (LPS) on the inflammation profiles of SAT and VAT explants from lean women, as well as on glyceroneogenesis, to test whether these two fat depots have intrinsically different responses to this metabolic endotoxin.
Design:
Abdominal SAT and VAT explants from eight lean women were treated in vitro with LPS. Their inflammatory status was evaluated by cytokine gene expression and secretion; glyceroneogenesis was evaluated by cytosolic phosphoenolpyruvate carboxykinase activity and FFA vs glycerol release.
Results:
In the basal state, the cytokine status and expression of macrophage markers were lower in SAT than VAT. In the presence of 100 ng ml−1 LPS, SAT exhibited a strong inflammatory response (increased interleukin-6 and tumor necrosis factor-α expression) and increased release of FFA due to inhibition of glyceroneogenesis, whereas VAT was only mildly affected. The effects of LPS on both tissues were blocked by the nuclear factor-κB (NF-κB) inhibitor, parthenolide. A significant effect of LPS on VAT occurred only at 1 μg ml−1 LPS.
Conclusion:
SAT explants from lean women are more sensitive to LPS-induced NF-κB activation than are VAT explants, leading to a depot-specific dysfunction of FFA storage. As SAT is the major player in FFA homoeostasis, this SAT dysfunction could be associated with visceral fat hypertrophy and systemic lipid disorders.
Similar content being viewed by others
Introduction
The roles of subcutaneous adipose tissue (SAT) and visceral adipose tissue (VAT) in metabolic deregulation are intrinsically different. In mouse models, SAT protects against insulin resistance and metabolic disorders, whereas VAT does not.1 In humans, however, the picture is less clear. VAT accumulation is associated with increased mortality and risk for diabetes and cardiovascular complications, whereas SAT hypertrophy is generally found to be neutral,2, 3 although other studies find no difference between SAT and VAT.4 Thus, it remains uncertain whether and how different human fat depots influence the occurrence of metabolic disease.5
Inflammation is a common feature of abnormal adipose tissue—whether hypertrophic in obesity6 or atrophied in lipodystrophic patients7, 8—and inflammation is linked to insulin resistance.9 Previous studies have shown an increased number of macrophages in adipose tissue from obese patients with both M1 and M2 phenotypes.10, 11 Upregulation of immune response-related gene expression in human SAT and VAT from obese subjects was found to be associated with downregulation of most of the adipocyte functions.4
Immune system homoeostasis is challenged by external insults, such as saturated free fatty acid (FFA)-rich diets.12 Moreover, in mice, a high-fat diet and/or acute fat load increases plasma lipopolysaccharide (LPS), the main component of the outer membrane of Gram-negative bacteria.13 Such ‘metabolic endotoxaemia’ can trigger an innate immune response, particularly in adipose tissue, which can result in the release of proinflammatory cytokines that interfere with glucose and lipid metabolism and induce insulin resistance.14, 15 A similar association between metabolic diseases and endotoxaemia seems to apply in humans too. Some obese and type 2 diabetes patients present with elevated levels of endotoxaemia16 together with alterations in their gut microflora,17, 18 and experimental endotoxaemia induces adipose tissue inflammation and systemic insulin resistance in lean subjects.19 Thus, LPS is potentially an endogenous inflammatory agent that might affect adipose tissue, whereas not being the sole mediator in obesity. Thus far, the effect of LPS on adipose tissue in humans has been analysed only in SAT,19 and any differential effect of LPS on SAT and VAT has not been investigated.
The primary role of adipose tissue is to store triglycerides, allowing the disposal of FFA according to the energy needed by the body. SAT is the major player in the systemic homoeostasis of FFA.20 Inducing high plasma FFA concentrations experimentally can reproduce some obesity-linked metabolic abnormalities in lean, healthy individuals. Conversely, lowering FFA concentrations can improve the metabolic health of android or diabetic, obese individuals.21
During fasting, the release of FFA into the blood is initiated by lipolysis of triglycerides stored in adipose tissue, but is limited by a simultaneous FFA re-esterification process in adipose tissue. This FFA–triglyceride recycling requires the de novo synthesis of glycerol-3 phosphate from lactate, a non-carbohydrate source, by glyceroneogenesis. The cytosolic form of phosphoenolpyruvate carboxykinase (PEPCKc) is the key enzyme that controls this pathway.22 A functional link between glyceroneogenesis and insulin sensitivity has been established in mice: the adipocyte-specific inactivation in mice of the gene encoding PEPCKc, PCK1, increased systemic insulin resistance associated with increased circulating FFA.23 By contrast, mice that overexpress PCK1 in adipocytes have reduced circulating FFA levels and normal insulin sensitivity, which has been associated with increased fat depots due to enhanced glyceroneogenesis and FFA re-esterification.24
We previously reported a progressively decreasing efficiency of glyceroneogenesis in controlling FFA output in human SAT explants associated with increasing body mass index of the sample donors, leading to a parallel increase in the release of FFA into the culture medium.25 These ‘ex vivo’ data suggested that glyceroneogenesis might be downregulated in adipocytes from overweight subjects and might thus contribute to the systemic increase in FFA in these individuals. We also reported that glyceroneogenesis is inhibited by anti-HIV drugs that activate the nuclear factor-κB (NF-κB) signalling pathway.26 This inflammation-linked inhibition of glyceroneogenesis with its concomitant increase in FFA release occurred in explants of SAT, but not in the paired VAT explants from the same individuals, suggesting that SAT and VAT might be fundamentally different in their responses to activators of inflammation and immunity. To test this hypothesis, we compared here the effect of LPS on markers of inflammation and FFA homoeostasis in SAT and VAT explants from lean subjects.
Materials and methods
Biopsies
Paired biopsies from abdominal SAT (periumbilical) and VAT (greater omental) were obtained from eight premenopausal women (38.3±6.3 years old) with body mass indices of 20.5–23.5 kg m−2 (average body mass index=21.9±1.5 kg m−2) undergoing elective exploratory laparotomy for endometriosis at Cochin hospital. Subjects were otherwise healthy and were receiving no medication or hormonal supplements, except for contraceptive pills. All subjects gave written informed consent, and all procedures were approved by the ethics committee of Cochin hospital. Samples, removed at the beginning of the surgery, were immediately transferred to the laboratory in Dulbecco’s modified Eagle’s medium containing 10% fetal bovine serum and 12.5 mM glucose.
Culture of adipose tissue explants
The tissue was carefully dissected to remove visible blood vessels and connective tissue, and then cut into small pieces of 20–35 mg wet weight, distributed in 12-well culture plates with about 200 mg per well and incubated in 2.5 ml of Dulbecco’s modified Eagle’s medium containing 10% fetal bovine serum and 5 mM glucose, biotin, pantothenate and antibiotics as described previously25 in a humidified atmosphere of 10% CO2 at 37 °C. After 4 h, explants were incubated with serum-free medium containing 0.5% (w:v) low-fat bovine serum albumin supplemented (or not) with LPS (100 ng ml−1) for 14 h (LPS from Escherichia coli 0111:B4; ref. L-4391 from Sigma, Tafkirchen, Germany). When mentioned in the main text, parthenolide (Alomone Labs, Jerusalem, Israel) was added to the medium to a final concentration of 30 μM 1 h before treatment with LPS. After incubation overnight, media were collected for the determination of cytokines, and explants were frozen in liquid nitrogen before RNA extraction.
Explants used to assay PEPCKc activity and lipolytic rate were incubated for 24 h. At 4 h before the analysis of FFA release, the medium was replaced with glucose- and serum-free Dulbecco’s modified Eagle’s medium containing 3% (w:v) low-fat bovine serum albumin to eliminate residual glycolytic and glycogenolytic intermediates. Then, the medium was replaced with 3 ml of the same medium supplemented with 2.5 mM pyruvate and 1 μM isoproterenol for 1 h. At the end of the incubation, the medium was collected for the estimation of FFA and glycerol released by lipolysis (below), and the corresponding tissue fragments were frozen in liquid nitrogen before the determination of PEPCKc activity.
Determination of PEPCKc-specific activity and biochemical analyses
PEPCKc activity was estimated in a post-mitochondrial fraction, prepared from adipose tissue previously homogenized in 10 mM Tris-HCl, pH 7.4, containing 1 mM dithiothreitol. PEPCKc activity was determined as described previously27 by the radioactive method of Chang and Lane,28 with saturating substrates concentration and 1:30 isotope dilution of [14C]NaHCO3; the reaction was performed for 30 min at 37 °C. Protein content was determined by using a BCA protein assay kit from Sigma-Aldrich. Concentrations of glycerol and FFA released in the incubation medium were monitored using free glycerol reagent (Sigma-Aldrich) and an acyl-CoA oxidase kit from Roche Applied Science (Meylan, France), respectively. In our experimental protocol, which was designed previously to analyse the efficiency of glyceroneogenesis in human SAT, the FFA:glycerol ratio was shown to be dependent on FFA re-esterification by glyceroneogenesis by using a PEPCKc inhibitor, 3-mercaptopicolinate.25
Levels of cytokines secreted into the culture medium
Cytokine analysis kits (Bioplex Magnetic Assay, BioRad, Marnes-la-Coquette, France) were used to quantify concentrations of interleukin (IL)-6 and IL-10 in the medium; Millipore multiscreen 96-well filtration plates were used for the assays (Millipore, Billerica, MA, USA). All samples were tested in duplicate with a Bioplex 200 system (Bio-Rad). All analyses were performed according to the manufacturer’s protocol. Dilutions of 1:100 and 1:1000 were needed for the quantification of IL-6 in SAT or VAT incubation media, respectively. Quantification of IL-10 secretion did not require dilution of the medium.
RNA and reverse transcription-polymerase chain reaction
RNA extraction was performed by using an RNeasy Lipid kit (Qiagen, Courtaboeuf, France), and the quality (meaning integrity) of the RNA was assessed by gel electrophoresis. In all, 1 μg of RNA was reverse transcribed using a high-capacity cDNA reverse transcription kit (Applied Biosystems, Carlsbad, CA, USA), and quantitative reverse transcription-polymerase chain reaction measurements were performed with various quantities of cDNA (5–20 ng), to control the linearity of the amplification, and 500 nM primers, using SYBR Green and a Roche thermocycler (Roche Diagnostics, Meylan, France). Variations in mRNA levels were estimated relative to those of 18S rRNA (ΔCT=CT gene of interest−CT 18S) obtained with 5 ng cDNA. The primer sequences were as follows: hPCK1 sense, 5′-GCTCTGAGGAGGAGAATGG-3′, hPCK1 antisense, 5′-TGCTCTTGGGTGACGATAAC-3′; hIL6 sense, 5′-CAAATTCGGTACATCCTCGAC-3′, hIL6 antisense, 5′-TTTCACCAGGCAAGTCTCCT-3′; hTNF-α sense, 5′-AATAGGCTGTTCCCATGTAGC-3′, hTNF-α antisense, 5′-AGAGGCTCAGCAATGAGTGA-3′; PPARγ sense, 5′-CAGTGGGGATGTCTCATAA-3′, PPARγ antisense, 5′-CTTTTGGCATACTCTGTGAT-3′; and h18S sense, 5′-GAGCGAAAGCATTTGCCAAG-3′, h18S antisense, 5′-GGCATCGTTTAGGTCGGAA-3′.
Statistical analysis
Data are presented as means±s.d., and the statistical significance of the difference between two means (untreated SAT vs untreated VAT, or treated SAT or VAT vs respective controls) were performed by the paired t-test. Significant differences were assumed for P-values <0.05.
Results
SAT and VAT explants from lean women have different inflammatory profiles
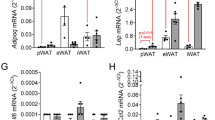
Pairs of abdominal SAT and omental VAT explants from eight individual, lean women were evaluated for their expression of inflammatory cytokines IL-6 and tumor necrosis factor (TNF)-αmRNAs (Figure 1a). The amount of IL-6 mRNA in VAT was 10.6-fold greater than in SAT and the amount of TNF-α mRNA was 1.6-fold greater (Figure 1a). Also, the amount of IL-6 secreted by VAT explants was markedly higher than that secreted by SAT explants (Figure 1b), as previously reported for TNF-α.26 We also evaluated the level of expression of the M1 macrophage marker CD68 and observed 3.7-fold more in VAT than in SAT; this is consistent with the large number of macrophages in this tissue, even in lean individuals29 and children.30 As human-resident adipose macrophages have both M2 and M1 phenotypic traits, we also evaluated the expression of two M2 markers CD163 and the anti-inflammatory cytokine IL-10. The level of CD163 mRNA was 1.4-fold greater in VAT than in SAT, and the secretion of IL-10 was 15-fold greater in VAT than in SAT (Figure 1).

Abdominal SAT and VAT explants from lean women have distinct inflammatory profiles. Paired adipose tissue explants were incubated for 14 h in Dulbecco’s modified Eagle’s medium containing 0.5% bovine serum albumin. (a) Relative mRNA expression normalized to 18s rRNA (N=8). Values are the means of VAT:SAT ratio±s.d. ΔCT vs 18S being for IL-6: 14.1±1.1 in SAT and 10.7±0.5 in VAT; for CD68: 13.7±0.2 in SAT and 11.8±0.6 in VAT; for CD163: 18.1±0.4 in SAT and 17.6±0.2 in VAT; and for TNFα: 20.1±0.6 in SAT and 19.4±0.2 in VAT. (b) Cytokine secretion into the incubation media. The data are expressed as mean ng g−1 of adipose tissue±s.d. (N=8). **P<0.01 and ***P<0.001.
These data suggest that, even in lean subjects, VAT contains more macrophages than does SAT, as it has higher levels of M1 and M2 markers. This likely reflects a local low-grade inflammation of VAT, indicated by high IL-6 and TNF-α gene expression and secretion, although this may be tempered by the high level of anti-inflammatory IL-10 secretion.
Depot-specific effects of LPS on human adipose tissue inflammation and FFA release by glyceroneogenesis
To analyse the effect of LPS on inflammation and FFA homoeostasis in SAT and VAT explants from lean subjects, we treated explants with LPS (100 ng ml−1) for 14 h; this concentration was chosen because it was shown previously to maximally increase TNF-α mRNA expression in macrophage cell lines.31 LPS treatment of SAT markedly increased IL-6 and TNF-α mRNA levels (by 16- and 12-fold, respectively), whereas LPS treatment of VAT had no significant effect on IL-6 expression and TNF-α expression increased by only threefold (Figure 2a). Consistent with the mRNA levels, secretion of IL-6 and TNF-α was significantly increased by LPS treatment of SAT (by 1.7- and 1.8-fold, respectively), but not by LPS treatment of VAT (both IL-6 and TNF-α expression increased by 1.1-fold, which was not statistically significant).

Depot-specific effects of LPS on human adipose tissue inflammation and FFA release by glyceroneogenesis. SAT and VAT explants were incubated with 100 ng ml−1 LPS for 14 h before gene expression analysis (a and b) and for 24 h for functional analysis of glyceroneogenesis (c). (a and b) Gene expression of cytokines or adipocyte markers: mRNA amounts were first normalized to the amount of 18S rRNA in each sample and then presented as a ratio vs the control, untreated SAT. (c) LPS effect on glyceroneogenesis: on the left, measure of the PEPCKc activity in each fat depot, in basal conditions. Data are the means of eight samples±s.d. On the right, measure of FFA and glycerol released in the incubation medium after treatment with isoproterenol. Data are the mean ratios of FFA:Gly±s.d. (N=8). Explants+LPS vs respective untreated explant: *P<0.05, **P<0.01 and ***P<0.001. Untreated VAT vs untreated SAT: #P<0.05 and ##P<0.01.
In parallel with our analysis of cytokines, we also determined the effect of LPS on metabolic genes involved in glyceroneogenesis; in response to LPS, the PPARγ and PCK1 mRNAs in SAT were substantially decreased (by 92% and 79%, respectively), whereas they were not significantly affected in VAT (Figure 2b). This effect of LPS on glyceroneogenesis was further investigated by assaying the specific activity of PEPCKc in the two fat depots and by measuring the release of FFA and glycerol into the medium after isoproterenol-induced lipolysis. At 24 h after LPS treatment, PEPCKc activity was reduced in SAT but not in VAT. Accordingly, the ratio of FFA:glycerol released (reflecting FFA re-esterification via glyceroneogenesis) increased only in the SAT explants (Figure 2c). We found no significant effect of LPS on the lipolytic index of either tissue, as determined from the amount of glycerol released (SAT: 1.42±0.17; LPS-SAT: 1.58±0.22; VAT: 2.37±0.32; LPS-VAT: 2.48±0.42 μmol h−1 g−1 of AT), whereas LPS increased the FFA release from SAT (SAT: 2.30±0.28; LPS-SAT: 4,31±0.53; VAT: 4.53±0.51; LPS-VAT: 5.26±0.77 μmol h−1 g−1 of AT).
These data show that human abdominal SAT from lean women is more sensitive to LPS-induced inflammation than is their VAT; this is associated with less expression of adipocyte genes involved in glyceroneogenesis and a corresponding decrease of PEPCKc activity, as well as increased release of FFA in conditions of acute lipolysis. By contrast, LPS (100 ng ml−1) does not affect glyceroneogenesis by VAT nor does it modulate FFA output.
LPS-induced inflammation and decreased glyceroneogenesis in human SAT are both mediated by NF-κB
As LPS acts by activating the NF-κB pathway, we used an inhibitor of NF-κB activation, parthenolide, to investigate whether the LPS effect on inflammation and glyceroneogenesis results from NF-B activation (Figure 3). When SAT explants were pre-treated with parthenolide 1 h before LPS treatment, the activating effect of LPS on IL-6 and TNF-α mRNA expression was prevented (Figure 3a). Accordingly, parthenolide significantly reduced LPS-induced secretion of IL-6 and TNF-α into the culture medium (data not shown). Likewise, in the presence of parthenolide the inhibitory effect of LPS on PPARγ and PCK1 gene expression was reduced (Figure 3b). Similarly, the glyceroneogenesis activity was restored by parthenolide: the specific activity of PEPCKc and the FFA:glycerol ratio remained at the level of untreated explants (Figure 3c).

LPS-induced inflammation and changes in glyceroneogenesis are linked to NF-κB activation in human SAT. SAT and VAT explants were incubated with 100 ng ml−1 LPS supplemented, or not, with 30 μM parthenolide (P), for 14 h for gene expression analysis (a and b) and 24 h for functional analysis of glyceroneogenesis (c). Gene expression and glyceroneogenesis were determined as in Figure 2, but the data are here presented as the fold change compared with their respective controls. LPS+P vs LPS: *P<0.05, **P<0.01 and ***P<0.001.
As for SAT, pre-treatment of VAT explants with parthenolide before LPS treatment reduced the small increase in inflammatory cytokine gene expression seen with LPS alone (Figure 3a), but had no significant effect on metabolic gene expression and glyceroneogenesis (Figures 3b and c). Parthenolide alone had no significant effect on the basal inflammation status of either fat depot (data not shown), indicating that the higher level of proinflammatory cytokines in VAT as compared with SAT seen in Figure 1 did not result from constitutive activation of the NF-κB pathway.
These data show that both LPS-induced inflammation and the LPS-induced decrease in glyceroneogenesis observed in SAT depend on the activation of the NF-κB pathway.
VAT is more resistant than SAT to the effects of LPS
We have reported previously that more IL-6 is required to inhibit the activity of PEPCKc in VAT than to inhibit the activity in SAT from lean women.26 We therefore tested here the effect of a 10-fold higher concentration of LPS than that used in the experiment in Figure 2 (that is, 1 μg ml−1) on VAT (Figure 4). This higher concentration of LPS induced secretion of IL-6 by VAT, which was not observed with 100 ng ml−1 LPS (Figure 4a). Likewise, the PEPCKc activity in VAT was decreased and the FFA:glycerol ratio in the medium was increased by the higher concentration of LPS (Figures 4b and c). These effects were mediated by the NF-κB pathway as they were reversed by the NF-κB inhibitor parthenolide (Figures 4b and c).

VAT is more resistant than SAT to the effects of LPS. VAT explants were treated with 100 ng ml−1 or 1 μg ml−1 LPS±30 μM parthenolide for 24 h. (a) IL-6 secretion was determined in the media after overnight incubation of the explants with LPS or excipient and calculated per gram of adipose tissue. (b) PEPCKc-specific activity was measured in nmol min−1 mg−1 of protein. (c) The FFA:glycerol ratio was calculated in each culture medium 1 h after the addition of isoproterenol to induce lipolysis. The data are presented as the fold change compared with their respective controls±s.d.: **P<0.01 and ***P<0.001.
These data confirm that NF-κB activation inhibits PEPCKc activity in VAT as it does in SAT. Nevertheless, they suggest that the NF-κB pathway in VAT requires a higher concentration of LPS to be activated than does the same pathway in SAT. Thus, VAT is more resistant than SAT to the effect of LPS on IL-6 secretion and glyceroneogenesis, but very high LPS concentrations can induce similar effects to those seen at lower concentrations in SAT.
Discussion
In this study, we describe the differential effect of a bacterial endotoxin LPS on abdominal subcutaneous and visceral fat depots from lean human subjects. We demonstrate a greater sensitivity of the SAT than VAT to LPS-induced NF-κB activation, which leads to a depot-specific decrease in glyceroneogenesis and increase of FFA release.
We studied samples from lean individuals because we wanted to analyse the effect of LPS, a potential initiator of inflammation and associated lipid disorders, under basal non-inflammatory conditions. Most previous studies, particularly studies of VAT, considered only adipose tissues from obese subjects, that is, hypertrophied tissues.
Our comparison of the basal inflammatory status of SAT and VAT in lean women confirmed that VAT had more resident macrophages than SAT, as first observed in lean subjects by Harman-Boehm et al.29 This was associated with increased expression and secretion of the proinflammatory cytokines IL-6 and TNF-α and of the anti-inflammatory cytokine IL-10. The increased expression of markers of M1 and M2 macrophages (CD68 or CD163) in VAT as compared with SAT suggests that VAT of lean women has both a higher basal inflammatory status than SAT and also a higher anti-inflammatory status. These data accord with the dual phenotype proposed for macrophages resident in adipose tissue in humans, which have both M1 and M2 markers.10, 29, 32 Interestingly, we found that the treatment of VAT with an NF-κB inhibitor, parthenolide, did not reduce this ‘basal’ inflammation in VAT, indicating that the low-level inflammation observed did not result from constitutive activation of the NF-κB pathway.
A number of recent studies point to the role of circulating LPS in triggering metabolic complications and insulin resistance.33, 34 In previous studies, LPS was found to stimulate in vitro proinflammatory cytokine secretion from adipocytes of SAT taken from lean human subjects.16 Injection of a small dose of LPS (3 ng kg−1) into lean subjects induced systemic insulin resistance that was preceded by stimulation of inflammatory status and downregulation of insulin signalling pathways in SAT. VAT was not evaluated in these studies, however, because the tissue was not available.19 The only published data about the effect of LPS on human visceral adipocytes used biological samples from the adipose tissue of obese subjects with a body mass index of 30–40 kg m−2;35 it found increased IL-6 secretion induced by LPS, with no significant change in the expression of PPARγ and fatty acid synthase mRNAs. The comparative responses of subcutaneous and visceral fat depots to experimental endotoxaemia has been studied only in mice and cats, where endotoxaemia led to a major increase in the expression of inflammatory adipokine genes in fat, with a precocious or higher inflammatory response in SAT than VAT.36, 37
We decided to investigate the release of FFA from subcutaneous and visceral fat depots because these lipids have an important role in the lipotoxicity process. To do so, we evaluated glyceroneogenesis, measured by PEPCKc activity and FFA:glycerol release, under basal conditions and after the induction of lipolysis, respectively. We found that LPS causes a substantial decrease in both PPARγ and PEPCKc gene expression, with a subsequent decrease in PEPCKc activity, as was recently observed in mice.38 Consequently, the amount of FFA released during lipolysis was increased by LPS-induced inflammation in the fat from lean women, and the SAT was far more sensitive than the VAT.
Our findings are in line with our previous observations that some antiretroviral protease inhibitors affect glyceroneogenesis in SAT but not VAT from lean subjects.26 Indeed, this effect of antiretroviral protease inhibitors, like the effect of LPS, requires NF-κB activation and correlates with increased local production of proinflammatory cytokines by SAT.26 The higher concentration of LPS required to affect VAT is reminiscent of our previous finding that IL-6 inhibits glyceroneogenesis in human VAT only at a 10-fold higher concentration than in SAT.26 We also confirmed the inhibitory effect of NF-κB activation on glyceroneogenesis in human fat, as recently observed in mice.39 Taken together, our data suggest that NF-κB-induced increased FFA release from SAT might favour ectopic FFA deposition in other tissues, including VAT, and therefore, potentially participating in the expansion of VAT.
Our experimental approach treating adipose tissue from normal, lean subjects with LPS simulates metabolic endotoxaemia, but does not recapitulate human obesity. A recent study4 that compared abdominal SAT and VAT gene expression in overweight and lean women, however, suggests that our findings could be relevant for obese subjets. In comparison with the lean women, the overweight group, characterized by increased VAT/total fat area and by hyperinsulinaemia, presented with increased inflammation in SAT, which was higher than in their VAT.
Our data show that the inflammatory agent LPS selectively increases FFA release from SAT, the major fat depot involved in FFA homoeostasis, whereas mildly affecting VAT. Considering that ectopic lipid accumulation precedes metabolic complications,40 we propose that damage of the FFA-buffering function of SAT by an inflammatory process (here, in response to LPS or by anti-human immunodeficiency virus drugs26) might participate in the expansion of VAT. Thus, our data provide novel insights into the differential contributions of abdominal subcutaneous and visceral human fat depots to metabolic complications.
References
Tran TT, Yamamoto Y, Gesta S, Kahn CR . Beneficial effects of subcutaneous fat transplantation on metabolism. Cell Metab 2008; 7: 410–420.
Despres JP, Lemieux I . Abdominal obesity and metabolic syndrome. Nature 2006; 444: 881–887.
Iozzo P . Viewpoints on the way to the consensus session: where does insulin resistance start? The adipose tissue. Diabetes Care 2009; 32 (Suppl 2): S168–S173.
Klimcakova E, Roussel B, Marquez-Quinones A, Kovacova Z, Kovacikova M, Combes M et al. Worsening of obesity and metabolic status yields similar molecular adaptations in human subcutaneous and visceral adipose tissue: decreased metabolism and increased immune response. J Clin Endocrinol Metab 2011; 96: E73–E82.
Gesta S, Tseng YH, Kahn CR . Developmental origin of fat: tracking obesity to its source. Cell 2007; 131: 242–256.
Wellen KE, Hotamisligil GS . Inflammation, stress, and diabetes. J Clin Invest 2005; 115: 1111–1119.
Jan V, Cervera P, Maachi M, Baudrimont M, Kim M, Vidal H et al. Altered fat differentiation and adipocytokine expression are inter-related and linked to morphological changes and insulin resistance in HIV-1-infected lipodystrophic patients. Antivir Ther 2004; 9: 555–564.
Gandotra S, Le Dour C, Bottomley W, Cervera P, Giral P, Reznik Y et al. Perilipin deficiency and autosomal dominant partial lipodystrophy. N Engl J Med 2011; 364: 740–748.
Hotamisligil GS . Inflammation and metabolic disorders. Nature 2006; 444: 860–867.
Bourlier V, Zakaroff-Girard A, Miranville A, De Barros S, Maumus M, Sengenes C et al. Remodeling phenotype of human subcutaneous adipose tissue macrophages. Circulation 2008; 117: 806–815.
Aron-Wisnewsky J, Tordjman J, Poitou C, Darakhshan F, Hugol D, Basdevant A et al. Human adipose tissue macrophages: m1 and m2 cell surface markers in subcutaneous and omental depots and after weight loss. J Clin Endocrinol Metab 2009; 94: 4619–4623.
Dasu MR, Jialal I . Free fatty acids in the presence of high glucose amplify monocyte inflammation via Toll-like receptors. Am J Physiol Endocrinol Metab 2011; 300: E145–E154.
Cani PD, Bibiloni R, Knauf C, Waget A, Neyrinck AM, Delzenne NM et al. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008; 57: 1470–1481.
Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007; 56: 1761–1772.
Serino M, Luche E, Gres S, Baylac A, Berge M, Cenac C et al. Metabolic adaptation to a high-fat diet is associated with a change in the gut microbiota. Gut 2012; 61: 543–553.
Creely SJ, McTernan PG, Kusminski CM, Fisher M, Da Silva NF, Khanolkar M et al. Lipopolysaccharide activates an innate immune system response in human adipose tissue in obesity and type 2 diabetes. Am J Physiol Endocrinol Metab 2007; 292: E740–E747.
Ley RE, Turnbaugh PJ, Klein S, Gordon JI . Microbial ecology: human gut microbes associated with obesity. Nature 2006; 444: 1022–1023.
Wu X, Ma C, Han L, Nawaz M, Gao F, Zhang X et al. Molecular characterisation of the faecal microbiota in patients with type II diabetes. Curr Microbiol 2010; 61: 69–78.
Mehta NN, McGillicuddy FC, Anderson PD, Hinkle CC, Shah R, Pruscino L et al. Experimental endotoxemia induces adipose inflammation and insulin resistance in humans. Diabetes 2010; 59: 172–181.
Jensen MD . Gender differences in regional fatty acid metabolism before and after meal ingestion. J Clin Invest 1995; 96: 2297–2303.
Jensen MD . Adipose tissue metabolism—an aspect we should not neglect? Horm Metab Res 2007; 39: 722–725.
Beale EG, Hammer RE, Antoine B, Forest C . Disregulated glyceroneogenesis: PCK1 as a candidate diabetes and obesity gene. Trends Endocrinol Metab 2004; 15: 129–135.
Millward CA, Desantis D, Hsieh CW, Heaney JD, Pisano S, Olswang Y et al. Phosphoenolpyruvate carboxykinase (Pck1) helps regulate the triglyceride/fatty acid cycle and development of insulin resistance in mice. J Lipid Res 2010; 51: 1452–1463.
Franckhauser S, Munoz S, Pujol A, Casellas A, Riu E, Otaegui P et al. Increased fatty acid re-esterification by PEPCK overexpression in adipose tissue leads to obesity without insulin resistance. Diabetes 2002; 51: 624–630.
Leroyer SN, Tordjman J, Chauvet G, Quette J, Chapron C, Forest C et al. Rosiglitazone controls fatty acid cycling in human adipose tissue by means of glyceroneogenesis and glycerol phosphorylation. J Biol Chem 2006; 281: 13141–13149.
Leroyer S, Vatier C, Kadiri S, Quette J, Chapron C, Capeau J et al. Glyceroneogenesis is inhibited through HIV protease inhibitor-induced inflammation in human subcutaneous but not visceral adipose tissue. J Lipid Res 2011; 52: 207–220.
Ballard FJ, Hanson RW . Purification of phosphoenolpyruvate carboxykinase from the cytosol fraction of rat liver and the immunochemical demonstration of differences between this enzyme and the mitochondrial phosphoenolpyruvate carboxykinase. J Biol Chem 1969; 244: 5625–5630.
Chang HC, Lane MD . The enzymatic carboxylation of phosphoenolpyruvate. II. Purification and properties of liver mitochondrial phosphoenolpyruvate carboxykinase. J Biol Chem 1966; 241: 2413–2420.
Harman-Boehm I, Bluher M, Redel H, Sion-Vardy N, Ovadia S, Avinoach E et al. Macrophage infiltration into omental versus subcutaneous fat across different populations: effect of regional adiposity and the comorbidities of obesity. J Clin Endocrinol Metab 2007; 92: 2240–2247.
Tam CS, Heilbronn LK, Henegar C, Wong M, Cowell CT, Cowley MJ et al. An early inflammatory gene profile in visceral adipose tissue in children. Int J Pediatr Obes 2011; 6: e360–e363.
Shi H, Kokoeva MV, Inouye K, Tzameli I, Yin H, Flier JS . TLR4 links innate immunity and fatty acid-induced insulin resistance. J Clin Invest 2006; 116: 3015–3025.
Zeyda M, Farmer D, Todoric J, Aszmann O, Speiser M, Gyori G et al. Human adipose tissue macrophages are of an anti-inflammatory phenotype but capable of excessive pro-inflammatory mediator production. Int J Obes (Lond) 2007; 31: 1420–1428.
Lassenius MI, Pietilainen KH, Kaartinen K, Pussinen PJ, Syrjanen J, Forsblom C et al. Bacterial endotoxin activity in human serum is associated with dyslipidemia, insulin resistance, obesity, and chronic inflammation. Diabetes Care 2011; 34: 1809–1815.
Pussinen PJ, Havulinna AS, Lehto M, Sundvall J, Salomaa V . Endotoxemia is associated with an increased risk of incident diabetes. Diabetes Care 2011; 34: 392–397.
Bassols J, Ortega FJ, Moreno-Navarrete JM, Peral B, Ricart W, Fernandez-Real JM . Study of the proinflammatory role of human differentiated omental adipocytes. J Cell Biochem 2009; 107: 1107–1117.
Leuwer M, Welters I, Marx G, Rushton A, Bao H, Hunter L et al. Endotoxaemia leads to major increases in inflammatory adipokine gene expression in white adipose tissue of mice. Pflugers Arch 2009; 457: 731–741.
Osto M, Zini E, Franchini M, Wolfrum C, Guscetti F, Hafner M et al. Subacute endotoxemia induces adipose inflammation and changes in lipid and lipoprotein metabolism in cats. Endocrinology 2011; 152: 804–815.
Feingold KR, Moser A, Shigenaga JK, Grunfeld C . Inflammation inhibits the expression of phosphoenolpyruvate carboxykinase in liver and adipose tissue. Innate Immun 2012; 18: 231–240.
Zhang J, Henagan TM, Gao Z, Ye J . Inhibition of glyceroneogenesis by histone deacetylase 3 contributes to lipodystrophy in mice with adipose tissue inflammation. Endocrinology 2011; 152: 1829–1838.
Unger RH . Minireview: weapons of lean body mass destruction: the role of ectopic lipids in the metabolic syndrome. Endocrinology 2003; 144: 5159–5165.
Acknowledgements
This study was sponsored by Assistance Publique—Hôpitaux de Paris (AP-HP) with a Contrat d'Initiation à la Recherche Clinique and by INSERM. CV is the recipient of a PhD grant from the Conseil Régional d’Ile de France (Cardiovasculaire-Obésité-Diabète Domaine d’Intérêt Majeur) and SK is a recipient of a PhD grant from the Ministère de l’Education Nationale et de la Recherche. BA is a CNRS researcher. We thank the IFR 65 plateforme microdosages for help with cytokine determination.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under the Creative Commons Attribution-NonCommercial-No Derivative Works 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Vatier, C., Kadiri, S., Muscat, A. et al. Visceral and subcutaneous adipose tissue from lean women respond differently to lipopolysaccharide-induced alteration of inflammation and glyceroneogenesis. Nutr & Diabetes 2, e51 (2012). https://doi.org/10.1038/nutd.2012.29
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/nutd.2012.29
Keywords
This article is cited by
-
Are gut dysbiosis, barrier disruption, and endotoxemia related to adipose tissue dysfunction in metabolic disorders? Overview of the mechanisms involved
Internal and Emergency Medicine (2023)
-
Pu’erh tea extract-mediated protection against hepatosteatosis and insulin resistance in mice with diet-induced obesity is associated with the induction of de novo lipogenesis in visceral adipose tissue
Journal of Gastroenterology (2017)
-
Lipopolysaccharide challenge significantly influences lipid metabolism and proteome of white adipose tissue in growing pigs
Lipids in Health and Disease (2015)
-
The limited storage capacity of gonadal adipose tissue directs the development of metabolic disorders in male C57Bl/6J mice
Diabetologia (2015)
-
Histological analysis of mammary gland remodeling caused by lipopolysaccharide in lactating mice
Cell and Tissue Research (2013)
