
Key Points
-
Multiple myeloma is a currently incurable B-cell malignancy characterized by excess monotypic plasma cells in the bone marrow in association with an excess of monoclonal protein in serum and/or urine.
-
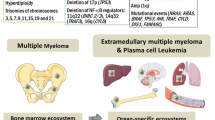
Multiple myeloma has complex heterogeneous cytogenetic abnormalities. Approximately 55–60% of patients have a hyperdiploid karyotype, which confers a better prognosis than those with non-hyperdiploid disease. Most non-hyperdiploid tumours have IgH translocations that involve several recurrent chromosomal loci, including 11q13 (cyclin D1), 6p21 (cyclin D3), 4p16 (FGFR3 and MMSET), 16q23 (MAF) and 20q11 (MAFB). Recent genomic and expression-profiling studies have both identified new therapeutic targets and provided the framework for a genetically based prognostic classification of multiple myeloma.
-
These constitutive genetic alterations in multiple myeloma cells and changes in gene-expression profiles mediate the protective effects of the bone marrow microenvironment on multiple myeloma cells.
-
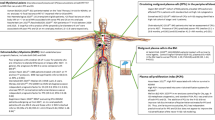
Multiple myeloma cells that home to the bone marrow have important functional sequelae. Specifically, the adhesion of multiple myeloma cells to extracellular matrix proteins confers cell adhesion-mediated drug resistance (CAMDR), and the binding of multiple myeloma cells to bone marrow accessory cells triggers the secretion of cytokines, which not only promote growth, survival and migration of multiple myeloma cells, but also confer resistance to conventional chemotherapy. Targeting these mechanisms offers a potential therapeutic strategy to overcome drug resistance.
-
Several factors, including MIP1α and RANKL, stimulate osteoclast activity; on the other hand, DKK1 inhibits osteoblastogenesis in multiple myeloma. This imbalance between bone formation and resorption results in osteolytic lesions, which are a hallmark of multiple myeloma.
-
New agents that target multiple myeloma cells, tumour–bone marrow interactions, or the bone marrow milieu, used alone or in combination, have shown promise in overcoming conventional drug resistance and improving patient outcome in multiple myeloma. Oncogenomics will allow for both patient selection and rational combination therapeutics.
Abstract
Multiple myeloma is a plasma cell malignancy characterized by complex heterogeneous cytogenetic abnormalities. The bone marrow microenvironment promotes multiple myeloma cell growth and resistance to conventional therapies. Although multiple myeloma remains incurable, novel targeted agents, used alone or in combination, have shown great promise to overcome conventional drug resistance and improve patient outcome. Recent oncogenomic studies have further advanced our understanding of the molecular pathogenesis of multiple myeloma, providing the framework for new prognostic classification and identifying new therapeutic targets.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others

References
Kyle, R. A. & Rajkumar, S. V. Monoclonal gammopathy of undetermined significance. Br. J. Haematol. 134, 573–589 (2006).
Ries, L. A. G. et al. (eds). SEER Cancer Statistics Review, 1975–2004 (National Cancer Institute, Bethesda, USA, 2007).
Kyle, R. A. & Rajkumar, S. V. Multiple myeloma. N. Engl. J. Med. 351, 1860–1873 (2004).
Singhal, S. et al. Antitumor activity of thalidomide in refractory multiple myeloma. N. Engl. J. Med. 341, 1565–1571 (1999).
Richardson, P. G. et al. Immunomodulatory drug CC-5013 overcomes drug resistance and is well tolerated in patients with relapsed multiple myeloma. Blood 100, 3063–3067 (2002).
Richardson, P. G. et al. A phase 2 study of bortezomib in relapsed, refractory myeloma. N. Engl. J. Med. 348, 2609–2617 (2003). Report of a phase III clinical trial that first demonstrated that a proteasome inhibitor, bortezomib, achieves responses in advanced multiple myeloma.
Richardson, P. G. et al. Bortezomib or high-dose dexamethasone for relapsed multiple myeloma. N. Engl. J. Med. 352, 2487–2498 (2005).
Gonzalez-Paz, N. et al. Tumor suppressor p16 methylation in multiple myeloma: biological and clinical implications. Blood 109, 1228–1232 (2007).
Pompeia, C. et al. Microarray analysis of epigenetic silencing of gene expression in the KAS-6/1 multiple myeloma cell line. Cancer Res. 64, 3465–3473 (2004).
Takahashi, T. et al. DNA methylation profiles of lymphoid and hematopoietic malignancies. Clin. Cancer Res. 10, 2928–2935 (2004).
Carrasco, D. R. et al. High-resolution genomic profiles define distinct clinico-pathogenetic subgroups of multiple myeloma patients. Cancer Cell 9, 313–325 (2006). The first high-resolution genomic analysis of multiple myeloma cell lines and patient samples. This work provides a classification based on aCGH profiles, and a list of potential oncogenes based on the merging of expression profiling with aCGH data.
Smadja, N. V. et al. Chromosomal analysis in multiple myeloma: cytogenetic evidence of two different diseases. Leukemia 12, 960–969 (1998). The first report on the existence of two patterns of cytogenetic abnormalities in multiple myeloma, hyperdiploid and non-hyperdiploid.
Debes-Marun, C. S. et al. Chromosome abnormalities clustering and its implications for pathogenesis and prognosis in myeloma. Leukemia 17, 427–436 (2003).
Chng, W. J. et al. Ploidy status rarely changes in myeloma patients at disease progression. Leuk. Res. 30, 266–271 (2006).
Fassas, A. B. et al. Both hypodiploidy and deletion of chromosome 13 independently confer poor prognosis in multiple myeloma. Br. J. Haematol. 118, 1041–1047 (2002).
Smadja, N. V., Bastard, C., Brigaudeau, C., Leroux, D. & Fruchart, C. Hypodiploidy is a major prognostic factor in multiple myeloma. Blood 98, 2229–2238 (2001).
Zhan, F. et al. The molecular classification of multiple myeloma. Blood 108, 2020–2028 (2006). This work provides a new classification of patients with multiple myeloma based on a comprehensive expression analysis of 414 newly diagnosed patients.
Avet-Loiseau, H. et al. Genetic abnormalities and survival in multiple myeloma: the experience of the Intergroupe Francophone du Myelome. Blood 109, 3489–3495 (2007).
Bergsagel, P. L. et al. Promiscuous translocations into immunoglobulin heavy chain switch regions in multiple myeloma. Proc. Natl Acad. Sci. USA 93, 13931–13936 (1996). A landmark paper reporting the first description of frequent translocations into the IgH switch regions in multiple myeloma.
Fonseca, R. et al. The recurrent IgH translocations are highly associated with nonhyperdiploid variant multiple myeloma. Blood 102, 2562–2567 (2003).
Bergsagel, P. L. & Kuehl, W. M. Chromosome translocations in multiple myeloma. Oncogene 20, 5611–5622 (2001). An excellent review of the mechanisms and consequences of chromosomal translocations in multiple myeloma.
Gabrea, A., Bergsagel, P. L., Chesi, M., Shou, Y. & Kuehl, W. M. Insertion of excised IgH switch sequences causes overexpression of cyclin D1 in a myeloma tumor cell. Mol. Cell 3, 119–123 (1999).
Chesi, M. et al. Dysregulation of cyclin D1 by translocation into an IgH γ switch region in two multiple myeloma cell lines. Blood 88, 674–681 (1996).
Shaughnessy, J. Jr. et al. Cyclin D3 at 6p21 is dysregulated by recurrent chromosomal translocations to immunoglobulin loci in multiple myeloma. Blood 98, 217–223 (2001).
Chesi, M. et al. Frequent translocation t(4;14)(p16. 3;q32. 3) in multiple myeloma: association with increased expression and activating mutations of fibroblast growth factor receptor 3. Nature Genet. 16, 260–264 (1997).
Chesi, M. et al. Frequent dysregulation of the c-maf proto-oncogene at 16q23 by translocation to an Ig locus in multiple myeloma. Blood 91, 4457–463 (1998).
Chesi, M. et al. The t(4;14) translocation in myeloma dysregulates both FGFR3 and a novel gene, MMSET, resulting in IgH/MMSET hybrid transcripts. Blood 92, 3025–3034 (1998).
Hurt, E. M. et al. Overexpression of c-maf is a frequent oncogenic event in multiple myeloma that promotes proliferation and pathological interactions with bone marrow stroma. Cancer Cell 5, 191–199 (2004).
Bergsagel, P. L. & Kuehl, W. M. Molecular pathogenesis and a consequent classification of multiple myeloma. J. Clin. Oncol. 23, 6333–6338 (2005).
Trudel, S. et al. Inhibition of fibroblast growth factor receptor 3 induces differentiation and apoptosis in t(4;14) myeloma. Blood 103, 3521–3528 (2004).
Trudel, S. et al. The inhibitory anti-FGFR3 antibody, PRO-001, is cytotoxic to t(4;14) multiple myeloma cells. Blood 107, 4039–4046 (2006).
Keats, J. J. et al. In multiple myeloma, t(4;14)(p16;q32) is an adverse prognostic factor irrespective of FGFR3 expression. Blood 101, 1520–1529 (2003).
Santra, M., Zhan, F., Tian, E., Barlogie, B. & Shaughnessy, J. Jr . A subset of multiple myeloma harboring the t(4;14)(p16;q32) translocation lacks FGFR3 expression but maintains an IGH/MMSET fusion transcript. Blood 101, 2374–2376 (2003).
Smadja, N. V. et al. Further cytogenetic characterization of multiple myeloma confirms that 14q32 translocations are a very rare event in hyperdiploid cases. Genes Chromosomes Cancer 38, 234–239 (2003).
Moreau, P. et al. Recurrent 14q32 translocations determine the prognosis of multiple myeloma, especially in patients receiving intensive chemotherapy. Blood 100, 1579–1583 (2002).
Fonseca, R. et al. Myeloma and the t(11;14)(q13;q32); evidence for a biologically defined unique subset of patients. Blood 99, 3735–3741 (2002).
Avet-Loiseau, H. et al. Cytogenetic, interphase, and multicolor fluorescence in situ hybridization analyses in primary plasma cell leukemia: a study of 40 patients at diagnosis, on behalf of the Intergroupe Francophone du Myelome and the Groupe Francais de Cytogenetique Hematologique. Blood 97, 822–825 (2001).
Fonseca, R. et al. Clinical and biologic implications of recurrent genomic aberrations in myeloma. Blood 101, 4569–4575 (2003).
Bergsagel, P. L. et al. Cyclin D dysregulation: an early and unifying pathogenic event in multiple myeloma. Blood 106, 296–303 (2005). This work provides a comprehensive analysis of the role of cyclin dysregulation in multiple myeloma, and proposes a multiple myeloma classification, the TC (translocation/cyclin D) classification, based on translocations and associated dysregulation of specific cyclins.
Hanamura, I. et al. Frequent gain of chromosome band 1q21 in plasma-cell dyscrasias detected by fluorescence in situ hybridization: incidence increases from MGUS to relapsed myeloma and is related to prognosis and disease progression following tandem stem-cell transplantation. Blood 108, 1724–1732 (2006).
Shou, Y. et al. Diverse karyotypic abnormalities of the c-myc locus associated with c-myc dysregulation and tumor progression in multiple myeloma. Proc. Natl Acad. Sci. USA 97, 228–233 (2000).
Kuehl, W. M. & Bergsagel, P. L. Multiple myeloma: evolving genetic events and host interactions. Nature Rev. Cancer 2, 175–187 (2002). An excellent review on the genetics of multiple myeloma.
Stewart, A. K. & Fonseca, R. Prognostic and therapeutic significance of myeloma genetics and gene expression profiling. J. Clin. Oncol. 23, 6339–6344 (2005).
Shaughnessy, J. D. Jr. et al. A validated gene expression model of high-risk multiple myeloma is defined by deregulated expression of genes mapping to chromosome 1. Blood 109, 2276–2284 (2007).
Jagannath, S. et al. Bortezomib appears to overcome the poor prognosis conferred by chromosome 13 deletion in phase 2 and 3 trials. Leukemia 21, 151–157 (2007).
Largo, C. et al. Identification of overexpressed genes in frequently gained/amplified chromosome regions in multiple myeloma. Haematologica 91, 184–191 (2006).
Intini, D. et al. Analysis of FGFR3 gene mutations in multiple myeloma patients with t(4;14). Br. J. Haematol. 114, 362–364 (2001).
Chesi, M. et al. Activated fibroblast growth factor receptor 3 is an oncogene that contributes to tumor progression in multiple myeloma. Blood 97, 729–736 (2001).
Corradini, P. et al. Mutational activation of N- and K- ras oncogenes in plasma cell dyscrasias. Blood 81, 2708–2713 (1993).
Bezieau, S. et al. High incidence of N and K-Ras activating mutations in multiple myeloma and primary plasma cell leukemia at diagnosis. Hum. Mutat. 18, 212–224 (2001).
Fonseca, R. et al. Genomic abnormalities in monoclonal gammopathy of undetermined significance. Blood 100, 1417–1424 (2002).
Zhan, F. et al. Gene-expression signature of benign monoclonal gammopathy evident in multiple myeloma is linked to good prognosis. Blood 109, 1692–700 (2007).
Keats, J. J., Reiman, T., Belch, A. R. & Pilarski, L. M. Ten years and counting: so what do we know about t(4;14)(p16;q32) multiple myeloma. Leuk. Lymphoma 47, 2289–2300 (2006).
Mitsiades, C. S. et al. In vitro and in vivo molecular profiling of multiple myeloma (MM) cell interaction with bone marrow (BM) microenvironment: insight into the role of novel anti-MM agents in counteracting BM-mediated drug-resistance in MM. Blood 102, 441a (2003).
Damiano, J. S., Cress, A. E., Hazlehurst, L. A., Shtil, A. A. & Dalton, W. S. Cell adhesion mediated drug resistance (CAM-DR): role of integrins and resistance to apoptosis in human myeloma cell lines. Blood 93, 1658–1667 (1999). This work demonstrates the concept of CAMDR, which overcomes conventional treatment in multiple myeloma.
Akiyama, M. et al. Cytokines modulate telomerase activity in a human multiple myeloma cell line. Cancer Res. 62, 3876–3882 (2002).
Chauhan, D. et al. Blockade of ubiquitin-conjugating enzyme CDC34 enhances anti-myeloma activity of Bortezomib/Proteasome inhibitor PS-341. Oncogene 23, 3597–3602 (2004).
Hideshima, T. et al. Perifosine, an oral bioactive novel alkylphospholipid, inhibits Akt and induces in vitro and in vivo cytotoxicity in human multiple myeloma cells. Blood 107, 4053–4062 (2006).
Mitsiades, C. S., Mitsiades, N. S., Munshi, N. C., Richardson, P. G. & Anderson, K. C. The role of the bone microenvironment in the pathophysiology and therapeutic management of multiple myeloma: interplay of growth factors, their receptors and stromal interactions. Eur. J. Cancer 42, 1564–1573 (2006).
Freund, G. G., Kulas, D. T. & Mooney, R. A. Insulin and IGF-1 increase mitogenesis and glucose metabolism in the multiple myeloma cell line, RPMI 8226. J. Immunol. 151, 1811 (1993).
Vanderkerken, K., Asosingh, K., Braet, F., Van Riet, I. & Van Camp, B. Insulin-like growth factor-1 acts as a chemoattractant factor for 5T2 multiple myeloma cells. Blood 93, 235–241 (1999).
Podar, K. et al. Vascular endothelial growth factor triggers signaling cascades mediating multiple myeloma cell growth and migration. Blood 98, 428–435 (2001).
Podar, K. et al. Vascular endothelial growth factor-induced migration of multiple myeloma cells is associated with beta 1 integrin- and phosphatidylinositol 3-kinase-dependent PKCα activation. J. Biol. Chem. 277, 7875–7881 (2002).
Hideshima, T. et al. The biological sequelae of stromal cell-derived factor-1α in multiple myeloma. Mol. Cancer Ther. 1, 539–544 (2002).
Mitsiades, C. S. et al. Inhibition of the insulin-like growth factor receptor-1 tyrosine kinase activity as a therapeutic strategy for multiple myeloma, other hematologic malignancies, and solid tumors. Cancer Cell 5, 221–230 (2004).
L'Hote, C. G. & Knowles, M. A. Cell responses to FGFR3 signalling: growth, differentiation and apoptosis. Exp. Cell Res. 304, 417–4131 (2005).
Otsuki, T. et al. Expression of fibroblast growth factor and FGF-receptor family genes in human myeloma cells, including lines possessing t(4;14)(q16. 3;q32. 3) and FGFR3 translocation. Int. J. Oncol. 15, 1205–1212 (1999).
Chauhan, D. et al. Multiple myeloma cell adhesion-induced interleukin-6 expression in bone marrow stromal cells involves activation of NF-κB. Blood 87, 1104–1112 (1996). This is the first study to demonstrate that transcription and secretion of IL6 from BMSCs are modulated by NFκB.
Chauhan, D. et al. A novel orally active proteasome inhibitor induces apoptosis in multiple myeloma cells with mechanisms distinct from Bortezomib. Cancer Cell 8, 407–419 (2005).
Moreaux, J. et al. BAFF and APRIL protect myeloma cells from apoptosis induced by interleukin 6 deprivation and dexamethasone. Blood 103, 3148–3157 (2004).
Hideshima, T. & Anderson, K. C. Molecular mechanisms of novel therapeutic approaches for multiple myeloma. Nature Rev. Cancer 2, 927–937 (2002).
Hideshima, T., Bergsagel, P. L., Kuehl, W. M. & Anderson, K. C. Advances in biology of multiple myeloma: clinical applications. Blood 104, 607–618 (2004).
Roodman, G. D. Pathogenesis of myeloma bone disease. Blood Cells Mol. Dis. 32, 290–292 (2004).
Roodman, G. D. New potential targets for treating myeloma bone disease. Clin. Cancer Res. 12, 6270s–6273s (2006).
Jakob, C. et al. Angiogenesis in multiple myeloma. Eur. J. Cancer 42, 1581–1590 (2006).
Ribatti, D., Nico, B. & Vacca, A. Importance of the bone marrow microenvironment in inducing the angiogenic response in multiple myeloma. Oncogene 25, 4257–4266 (2006).
De Clercq, E. Potential clinical applications of the CXCR4 antagonist bicyclam AMD3100. Mini Rev. Med. Chem. 5, 805–824 (2005).
Alsayed, Y. et al. Mechanisms of regulation of CXCR4/SDF-1 (CXCL12) dependent migration and homing in multiple myeloma. Blood 109, 2708–2717 (2006).
Landowski, T. H., Olashaw, N. E., Agrawal, D. & Dalton, W. S. Cell adhesion-mediated drug resistance (CAM-DR) is associated with activation of NFκB (RelB/p50) in myeloma cells. Oncogene 22, 2417–2421 (2003).
Damiano, J. S. & Dalton, W. S. Integrin-mediated drug resistance in multiple myeloma. Leuk. Lymphoma 38, 71–81 (2000).
Hazlehurst, L. A., Damiano, J. S., Buyuksal, I., Pledger, W. J. & Dalton, W. S. Adhesion to fibronectin via β1 integrins regulates p27kip1 levels and contributes to cell adhesion mediated drug resistance (CAM-DR). Oncogene 19, 4319–4327 (2000).
Hazlehurst, L. A. et al. Genotypic and phenotypic comparisons of de novo and acquired melphalan resistance in an isogenic multiple myeloma cell line model. Cancer Res. 63, 7900–7906 (2003).
Lwin, T. et al. Cell adhesion induces p27Kip1-associated cell-cycle arrest through down-regulation of the SCFSkp2 ubiquitin ligase pathway in mantle cell and other non-Hodgkin's B-cell lymphomas. Blood 14 May 2007 (epub ahead of print).
Sanderson, R. D., Turnbull, J. E., Gallagher, J. T. & Lander, A. D. Fine structure of heparan sulfate regulates syndecan-1 function and cell behavior. J. Biol. Chem. 269, 13100–13106 (1994).
Yang, Y. et al. Soluble syndecan-1 promotes growth of myeloma tumors in vivo. Blood 100, 610–617 (2002).
Yang, Y. et al. Heparanase enhances syndecan-1 shedding: a novel mechanism for stimulation of tumor growth and metastasis. J. Biol. Chem. 282, 13326–13333 (2007).
Hideshima, T., Chauhan, D., Schlossman, R. L., Richardson, P. R. & Anderson, K. C. Role of TNF-a in the pathophysiology of human multiple myeloma: therapeutic applications. Oncogene 20, 4519–4527 (2001).
Urashima, M. et al. Transforming growth factor b1: differential effects on multiple myeloma versus normal B cells. Blood 87, 1928–1938 (1996).
Dankbar, B. et al. Vascular endothelial growth factor and interleukin-6 in paracrine tumor-stromal cell interactions in multiple myeloma. Blood 95, 2630–2636 (2000).
Gupta, D. et al. Adherence of multiple myeloma cells to bone marrow stromal cells upregulates vascular endothelial growth factor secretion: therapeutic applications. Leukemia 15, 1950–1561 (2001).
Urashima, M., Chauhan, D., Uchiyama, H., Freeman, G. J. & Anderson, K. C. CD40 ligand triggered interleukin-6 secretion in multiple myeloma. Blood 85, 1903–1912 (1995).
Tai, Y. T. et al. Mechanisms by which SGN-40, a humanized anti-CD40 antibody, induces cytotoxicity in human multiple myeloma cells: clinical implications. Cancer Res. 64, 2846–2852 (2004).
Tai, Y. T. et al. Human anti-CD40 antagonist antibody triggers significant antitumor activity against human multiple myeloma. Cancer Res. 65, 5898–5906 (2005).
Hideshima, T. et al. The proteasome inhibitor PS-341 inhibits growth, induces apoptosis, and overcomes drug resistance in human multiple myeloma cells. Cancer Res. 61, 3071–3076 (2001).
Bergsagel, P. L. & Kuehl, W. M. Critical roles for immunoglobulin translocations and cyclin D dysregulation in multiple myeloma. Immunol. Rev. 194, 96–104 (2003).
Vacca, A. et al. Bone marrow neovascularization, plasma cell angiogenic potential, and matrix metalloproteinase-2 secretion parallel progression of human multiple myeloma. Blood 93, 3064–3073 (1999).
Roccaro, A. M. et al. Bortezomib mediates antiangiogenesis in multiple myeloma via direct and indirect effects on endothelial cells. Cancer Res. 66, 184–191 (2006).
Kline, M. et al. Cytokine and chemokine profiles in multiple myeloma; significance of stromal interaction and correlation of IL-8 production with disease progression. Leuk. Res. 31, 591–598 (2007).
Rajkumar, S. V. et al. Bone marrow angiogenesis in 400 patients with monoclonal gammopathy of undetermined significance, multiple myeloma, and primary amyloidosis. Clin. Cancer Res. 8, 2210–2216 (2002).
Andersen, N. F. et al. Syndecan-1 and angiogenic cytokines in multiple myeloma: correlation with bone marrow angiogenesis and survival. Br. J. Haematol. 128, 210–217 (2005).
Vacca, A. et al. Endothelial cells in the bone marrow of patients with multiple myeloma. Blood 102, 3340–3348 (2003).
Vacca, A. et al. Thalidomide downregulates angiogenic genes in bone marrow endothelial cells of patients with active multiple myeloma. J. Clin. Oncol. 23, 5334–5346 (2005).
Ribatti, D. & Vacca, A. Novel therapeutic approaches targeting vascular endothelial growth factor and its receptors in haematological malignancies. Curr. Cancer Drug Targets 5, 573–578 (2005).
Mundy, G. R., Raisz, L., G,, Cooper, R. A., Schecter, G. P. & Salmon, S. E. Evidence for the secretion of an osteoclast stimulating factor in myeloma. N. Engl. J. Med. 291, 1041–1046 (1974).
Yaccoby, S. et al. Myeloma interacts with the bone marrow microenvironment to induce osteoclastogenesis and is dependent on osteoclast activity. Br. J. Haematol. 116, 278–290 (2002).
Gori, F. et al. The expression of osteoprotegerin and RANK ligand and the support of osteoclast formation by stromal-osteoblast lineage cells is developmentally regulated. Endocrinology 141, 4768–4776 (2000).
Giuliani, N., Colla, S. & Rizzoli, V. New insight in the mechanism of osteoclast activation and formation in multiple myeloma: focus on the receptor activator of NFκB ligand (RANKL). Exp. Hematol. 32, 685–691 (2004).
Atkins, G. J. et al. RANKL expression is related to the differentiation state of human osteoblasts. J. Bone Miner. Res. 18, 1088–1098 (2003).
Sezer, O., Heider, U., Zavrski, I., Kuhne, C. A. & Hofbauer, L. C. RANK ligand and osteoprotegerin in myeloma bone disease. Blood 101, 2094–2098 (2003).
Hofbauer, L. C. & Schoppet, M. Clinical implications of the osteoprotegerin/RANKL/RANK system for bone and vascular diseases. JAMA 292, 490–495 (2004).
Croucher, P. I. et al. Osteoprotegerin inhibits the development of osteolytic bone disease in multiple myeloma. Blood 98, 3534–3540 (2001).
Michigami, T. et al. Cell–cell contact between marrow stromal cells and myeloma cells via VCAM-1 and α(4)β(1)-integrin enhances production of osteoclast-stimulating activity. Blood 96, 1953–1960 (2000).
Pearse, R. N. et al. Multiple myeloma disrupts the TRANCE/ osteoprotegerin cytokine axis to trigger bone destruction and promote tumor progression. Proc. Natl Acad. Sci. USA 98, 11581–11586 (2001).
Giuliani, N., Bataille, R., Mancini, C., Lazzaretti, M. & Barille, S. Myeloma cells induce imbalance in the osteoprotegerin/osteoprotegerin ligand system in the human bone marrow environment. Blood 98, 3527–3533 (2001).
Choi, S. J. et al. Antisense inhibition of macrophage inflammatory protein 1-a blocks bone destruction in a model of myeloma bone disease. J. Clin. Invest. 108, 1833–1841 (2001).
Oba, Y. et al. MIP1α utilizes both CCR1 and CCR5 to induce osteoclast formation and increase adhesion of myeloma cells to marrow stromal cells. Exp. Hematol. 33, 272–278 (2005).
Roodman, G. D. et al. Interleukin 6. A potential autocrine/paracrine factor in Paget's disease of bone. J. Clin. Invest. 89, 46–52 (1992).
Nguyen, A. N. et al. Normalizing the bone marrow microenvironment with p38 inhibitor reduces multiple myeloma cell proliferation and adhesion and suppresses osteoclast formation. Exp. Cell Res. 312, 1909–1923 (2006).
Yin, L. Chondroitin synthase 1 is a key molecule in myeloma cell-osteoclast interactions. J. Biol. Chem. 280, 15666–15672 (2005).
Ducy, P., Zhang, R., Geoffroy, V., Ridall, A. L. & Karsenty, G. Osf2/Cbfa1: a transcriptional activator of osteoblast differentiation. Cell 89, 747–754 (1997).
Karsenty, G. et al. Cbfa1 as a regulator of osteoblast differentiation and function. Bone 25, 107–108 (1999).
Giuliani, N., Rizzoli, V. & Roodman, G. D. Multiple myeloma bone disease: pathophysiology of osteoblast inhibition. Blood 108, 3992–3996 (2006).
Giuliani, N. et al. Myeloma cells block RUNX2/CBFA1 activity in human bone marrow osteoblast progenitors and inhibit osteoblast formation and differentiation. Blood 106, 2472–2483 (2005).
Tian, E. et al. The role of the Wnt-signaling antagonist DKK1 in the development of osteolytic lesions in multiple myeloma. N. Engl. J. Med. 349, 2483–2494 (2003). This is the first study showing that DKK1 inhibits osteoblastogenesis in patients with multiple myeloma.
Yaccoby, S. et al. Antibody-based inhibition of DKK1 suppresses tumor-induced bone resorption and multiple myeloma growth in vivo. Blood 109, 2106–2111 (2007).
Glass, D. A. 2nd et al. Canonical Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Dev. Cell 8, 751–764 (2005).
Holmen, S. L. et al. Essential role of β-catenin in postnatal bone acquisition. J. Biol. Chem. 280, 21162–21168 (2005).
Spencer, G. J., Utting, J. C., Etheridge, S. L., Arnett, T. R. & Genever, P. G. Wnt signalling in osteoblasts regulates expression of the receptor activator of NFκB ligand and inhibits osteoclastogenesis in vitro. J. Cell Sci. 119, 1283–1296 (2006).
Oshima, T. et al. Myeloma cells suppress bone formation by secreting a soluble Wnt inhibitor, sFRP-2. Blood 106, 3160–3165 (2005).
Ehrlich, L. A. et al. IL3 is a potential inhibitor of osteoblast differentiation in multiple myeloma. Blood 106, 1407–1414 (2005).
Franchimont, N., Rydziel, S. & Canalis, E. Transforming growth factor-β increases interleukin-6 transcripts in osteoblasts. Bone 26, 249–253 (2000).
Standal, T. et al. HGF inhibits BMP-induced osteoblastogenesis: possible implications for the bone disease of multiple myeloma. Blood 109, 3024–3030 (2006).
Murray, E. J., Bentley, G. V., Grisanti, M. S. & Murray, S. S. The ubiquitin-proteasome system and cellular proliferation and regulation in osteoblastic cells. Exp. Cell Res. 242, 460–469 (1998).
Garrett, I. R. et al. Selective inhibitors of the osteoblast proteasome stimulate bone formation in vivo and in vitro. J. Clin. Invest. 111, 1771–1782 (2003).
Heider, U. et al. Bortezomib increases osteoblast activity in myeloma patients irrespective of response to treatment. Eur. J. Haematol. 77, 233–238 (2006).
Mitsiades, C. S., Mitsiades, N., Munshi, N. C. & Anderson, K. C. Focus on multiple myeloma. Cancer Cell 6, 439–444 (2004).
Gunn, W. G. et al. A crosstalk between myeloma cells and marrow stromal cells stimulates production of DKK1 and interleukin-6: a potential role in the development of lytic bone disease and tumor progression in multiple myeloma. Stem Cells 24, 986–991 (2006).
Raje, N. et al. Seliciclib (CYC202 or R-roscovitine), a small-molecule cyclin-dependent kinase inhibitor, mediates activity via down-regulation of Mcl-1 in multiple myeloma. Blood 106, 1042–1047 (2005).
Mitsiades, C. S. et al. Antimyeloma activity of heat shock protein-90 inhibition. Blood 107, 1092–1100 (2006).
Mitsiades, C. S. et al. Activation of NF-kB and upregulation of intracellular anti-apoptotic proteins via the IGF-1/Akt signaling in human multiple myeloma cells: therapeutic implications. Oncogene 21, 5673–5683 (2002).
Shaughnessy, J. D. Jr. & Barlogie, B. Using genomics to identify high-risk myeloma after autologous stem cell transplantation. Biol. Blood Marrow Transplant. 12, 77–80 (2006).
Walker, B. A. et al. Integration of global SNP-based mapping and expression arrays reveals key regions, mechanisms, and genes important in the pathogenesis of multiple myeloma. Blood 108, 1733–1743 (2006).
Stewart, A. K. et al. A practical guide to defining high-risk myeloma for clinical trials, patient counseling and choice of therapy. Leukemia 21, 529–534 (2007).
Chng, W. J. et al. Molecular dissection of hyperdiploid multiple myeloma by gene expression profiling. Cancer Res. 67, 2982–2989 (2007).
Mitsiades, N. et al. Molecular sequelae of histone deacetylase inhibition in human malignant B cells. Blood 101, 4055–4062 (2003).
Catley, L. et al. NVP-LAQ824 is a potent novel histone deacetylase inhibitor with significant activity against multiple myeloma. Blood 102, 2615–2622 (2003).
Mitsiades, C. S. et al. Transcriptional signature of histone deacetylase inhibition in multiple myeloma: biological and clinical implications. Proc. Natl Acad. Sci. USA 101, 540–545 (2004).
Catley, L. et al. Aggresome induction by proteasome inhibitor bortezomib and α-tubulin hyperacetylation by tubulin deacetylase (TDAC) inhibitor LBH589 are synergistic in myeloma cells. Blood 108, 3441–3449 (2006).
Zhu, W. G. & Otterson, G. A. The interaction of histone deacetylase inhibitors and DNA methyltransferase inhibitors in the treatment of human cancer cells. Curr. Med. Chem. Anti-Canc. Agents 3, 187–199 (2003).
Hu, J. & Colburn, N. H. Histone deacetylase inhibition down-regulates cyclin D1 transcription by inhibiting nuclear factor-κB/p65 DNA binding. Mol. Cancer Res. 3, 100–109 (2005).
Caraglia, M., Tagliaferri, P., Budillon, A. & Abbruzzese, A. Post-translational modifications of eukaryotic initiation factor-5A (eIF-5A) as a new target for anti-cancer therapy. Adv. Exp. Med. Biol. 472, 187–198 (1999).
Palakurthi, S. S. et al. Inhibition of translation initiation mediates the anticancer effect of the n-3 polyunsaturated fatty acid eicosapentaenoic acid. Cancer Res. 60, 2919–2925 (2000).
Dragnev, K. H., Freemantle, S. J., Spinella, M. J. & Dmitrovsky, E. Cyclin proteolysis as a retinoid cancer prevention mechanism. Ann. NY Acad. Sci. 952, 13–22 (2001).
Tsutsumi, S. et al. Regulation of cell proliferation by autocrine motility factor/phosphoglucose isomerase signaling. J. Biol. Chem. 278, 32165–32172 (2003).
Malumbres, M. & Barbacid, M. To cycle or not to cycle: a critical decision in cancer. Nature Rev. Cancer 1, 222–231 (2001).
Sherr, C. J. & McCormick, F. The RB and p53 pathways in cancer. Cancer Cell 2, 103–112 (2002).
Chen, J. et al. FGFR3 as a therapeutic target of the small molecule inhibitor PKC412 in hematopoietic malignancies. Oncogene 24, 8259–8267 (2005).
Chang, H. et al. Immunohistochemistry accurately predicts FGFR3 aberrant expression and t(4;14) in multiple myeloma. Blood 106, 353–355 (2005).
Trudel, S. et al. CHIR-258, a novel, multitargeted tyrosine kinase inhibitor for the potential treatment of t(4;14) multiple myeloma. Blood 105, 2941–2948 (2005).
Urashima, M. et al. The development of a model for the homing of multiple myeloma cells to human bone marrow. Blood 90, 754–765 (1997).
Mitsiades, C. S. et al. Fluorescence imaging of multiple myeloma cells in a clinically relevant SCID/NOD in vivo model: biologic and clinical implications. Cancer Res. 63, 6689–6696 (2003).
Tassone, P. et al. Cytotoxic activity of the maytansinoid immunoconjugate B-B4-DM1 against CD138+ multiple myeloma cells. Blood 104, 3688–3696 (2004).
Tassone, P. et al. A clinically relevant SCID-hu in vivo model of human multiple myeloma. Blood 106, 713–716 (2005).
Carrasco, D. R. et al. The differentiation and stress response factor XBP-1 drives multiple myeloma pathogenesis. Cancer Cell 11, 349–360 (2007). This work provides a description of the first genetically engineered mouse model recapitulating the main clinical and pathogenetic features of human multiple myeloma.
Iwakoshi, N. N. et al. Plasma cell differentiation and the unfolded protein response intersect at the transcription factor XBP-1. Nature Immunol. 4, 321–329 (2003).
Klein, B. et al. Survival and proliferation factors of normal and malignant plasma cells. Int. J. Hematol. 78, 106–113 (2003).
Mitsiades, N. et al. Apoptotic signaling induced by immunomodulatory thalidomide analogs in human multiple myeloma cells: therapeutic implications. Blood 99, 4525–4530 (2002).
Hideshima, H. et al. Small molecule inhibition of proteasome and aggresome function induces synergistic anti-tumor activity in multiple myeloma. Proc. Natl Acad. Sci. USA 102, 8567–8572 (2005).
Chauhan, D. et al. A novel Bcl-2/Bcl-X(L)/Bcl-w inhibitor ABT-737 as therapy in multiple myeloma. Oncogene 26, 2374–2380 (2007).
Chauhan, D. et al. Targeting mitochondrial factor Smac/DIABLO as therapy for multiple myeloma (MM). Blood 109, 1220–1227 (2007).
Hideshima, T. et al. Molecular mechanisms mediating antimyeloma activity of proteasome inhibitor PS-341. Blood 101, 1530–1534 (2003).
Mitsiades, N. et al. The proteasome inhibitor PS-341 potentiates sensitivity of multiple myeloma cells to conventional chemotherapeutic agents: therapeutic applications. Blood 101, 2377–2380 (2003).
Berenson, J. R. et al. Phase I/II trial assessing bortezomib and melphalan combination therapy for the treatment of patients with relapsed or refractory multiple myeloma. J. Clin. Oncol. 24, 937–944 (2006).
Orlowski, R. Z. et al. Phase 1 trial of the proteasome inhibitor bortezomib and pegylated liposomal doxorubicin in patients with advanced hematologic malignancies. Blood 105, 3058–3065 (2005).
Mitsiades, N. et al. Molecular sequelae of proteasome inhibition in human multiple myeloma cells. Proc. Natl Acad. Sci. USA 99, 14374–14379 (2002).
Podar, K. et al. Caveolin-1 is required for vascular endothelial growth factor-triggered multiple myeloma cell migration and is targeted by bortezomib. Cancer Res. 64, 7500–7506 (2004).
Podar, K. et al. Targeting PKC in multiple myeloma: in vitro and in vivo effects of the novel, orally available small-molecule inhibitor enzastaurin (LY317615. HCl). Blood 109, 1669–1677 (2007).
Lorch, J. H., Thomas, T. O. & Schmoll, H. J. Bortezomib inhibits cell-cell adhesion and cell migration and enhances epidermal growth factor receptor inhibitor-induced cell death in squamous cell cancer. Cancer Res. 67, 727–734 (2007).
Hideshima, T. et al. Thalidomide and its analogues overcome drug resistance of human multiple myeloma cells to conventional therapy. Blood 96, 2943–2950 (2000).
Raje, N. et al. Combination of the mTOR inhibitor Rapamycin and RevlimidTM(CC-5013) has synergistic activity in multiple myeloma. Blood 104, 4188–4193 (2004).
Tai, Y. T. et al. Immunomodulatory drug lenalidomide (CC-5013, IMiD3) augments anti-CD40 SGN-40-induced cytotoxicity in human multiple myeloma: clinical implications. Cancer Res. 65, 11712–11720 (2005).
Rajkumar, S. V. et al. Combination therapy with lenalidomide plus dexamethasone (Rev/Dex) for newly diagnosed myeloma. Blood 106, 4050–4053 (2005).
Dispenzieri, A. et al. The activity of lenalidomide with or without dexamethasone in patients with primary systemic amyloidosis. Blood 109, 465–470 (2007).
Mulligan, G. et al. Gene expression profiling and correlation with outcome in clinical trials of the proteasome inhibitor bortezomib. Blood 109, 3177–3188 (2007).
Altun, M. et al. Effects of PS-341 on the activity and composition of proteasomes in multiple myeloma cells. Cancer Res. 65, 7896–7901 (2005).
Berkers, C. R. et al. Activity probe for in vivo profiling of the specificity of proteasome inhibitor bortezomib. Nature Methods 2, 357–362 (2005).
Munshi, N. C. et al. Identification of genes modulated in multiple myeloma using genetically identical twin samples. Blood 103, 1799–1806 (2004).
Bergsagel, P. L. et al. Promiscuous mutations frequently activate the non-canonical NFκB pathway in multiple myeloma (MM). Blood 108, 36a (2006).
Acknowledgements
The authors would like to thank D. Chauhan and N.C. Munshi for their helpful comments. We apologize to our many colleagues whose work is not cited owing to space constraints.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Related links
Related links
DATABASES
National Cancer Institute
FURTHER INFORMATION
US National Cancer Institute Clinical Trials website
Glossary
- Plasma cell
-
A plasma cell differentiates from a B lymphocyte and secretes large amounts of immunoglobulins. Plasma cells are rarely found in the peripheral blood, and represent 0.2% to 2.8% of bone marrow mononuclear cells.
- Monoclonal (M) protein
-
An antibody (immunoglobulin), or part of an antibody, found in excess in the blood and/or urine in patients with multiple myeloma and other plasma cell tumours.
- Plasmacytosis
-
A condition in which there is an increased population of plasma cells.
- Hyperviscosity
-
Increased viscosity of blood caused by an excess of M proteins in multiple myeloma.
- Bone marrow stromal cells
-
(BMSCs). Cellular components in the bone marrow that support multiple myeloma cell growth, survival and drug resistance.
- Thalidomide, lenalidomide and bortezomib
-
Immunomodulatory drugs and proteasome inhibitors that target not only multiple myeloma cells, but also bone marrow accessory cells, and overcome conventional drug resistance.
- Cell adhesion-mediated drug resistance
-
(CAMDR) Resistance to conventional chemotherapy owing to the adhesion of multiple myeloma cells to accessory cells in the bone marrow.
- Bone marrow accessory cells
-
Accessory cells in the bone marrow include stromal cells, osteoclasts, osteoblasts and endothelial cells.
- Matrix metalloproteinase
-
(MMP) MMPs are endopeptidases that degrade extracellular matrix proteins (such as collagen) and chemokines (such as SDF1). They are activated by other MMPs and plasmin, and inactivated by tissue inhibitor of metalloproteinases (TIMPs). Multiple myeloma cells express MMP9 and BMSCs express MMP1 and 2. The interaction of multiple myeloma cells and BMSCs upregulates MMP1, promoting bone resorption and tumour invasion.
- Plasma cell leukaemia
-
(PCL). PCL is a variant of multiple myeloma characterized by greater than or equal to 2 × 109 circulating plasma cells in one litre of peripheral blood. Pateints can present with primary PCL, or it can evolve from previously recognized multiple myeloma (secondary PCL).
- Microvessel density
-
(MVD). A quantitative measure of the density of small vessels in tissues, which is assessed by immunohistochemical staining with anti-endothelial antibodies (such as anti-CD31). In multiple myeloma, increasing MVD is associated with progressive disease.
- Osteopaenia
-
One of the most common findings in skeletal radiographs in patients with multiple myeloma is the increased radiolucency of bone, termed osteopaenia. Osteopaenia is caused by low mineral density.
- SCID mouse models
-
In vivo xenograft models of human multiple myeloma in immunodeficient mice. In the NOD;LtSz-scid;scid (SCID/NOD) model, diffuse fluorescence-protein labelled multiple myeloma cells are injected intravenously and their homing and anatomical localization are monitored by whole body-bioluminescence. In the SCID-hu model, a human bone chip is implanted subcutaneously into SCID mice, and human multiple myeloma cells directly injected into the bone graft. This allows for the evaluation of human multiple myeloma cells in the context of human bone marrow accessory cells and extracellular matrix proteins.
- Aggresome autophagy
-
Perinuclear inclusions that form in multiple myeloma cells in the setting of stress associated with misfolded or mutated proteins. Several protein components, including ubiquitin, HDAC6, dynein and vimentin have a crucial role in aggresome formation. These protein complexes are ultimately degraded by lysosomes.
- Antibody-dependent cell-mediated cytotoxicity
-
(ADCC). An immune response in which antibodies bind to epitopes on target cells, thereby marking them for attack by effector cells (such as natural killer cells, T cells and macrophages). It is necessary for effector cells to have Fc receptors.
Rights and permissions
About this article
Cite this article
Hideshima, T., Mitsiades, C., Tonon, G. et al. Understanding multiple myeloma pathogenesis in the bone marrow to identify new therapeutic targets. Nat Rev Cancer 7, 585–598 (2007). https://doi.org/10.1038/nrc2189
Issue Date:
DOI: https://doi.org/10.1038/nrc2189
This article is cited by
-
Characterization and application of a lactate and branched chain amino acid metabolism related gene signature in a prognosis risk model for multiple myeloma
Cancer Cell International (2023)
-
The roles of bone remodeling in normal hematopoiesis and age-related hematological malignancies
Bone Research (2023)
-
FTO promotes Bortezomib resistance via m6A-dependent destabilization of SOD2 expression in multiple myeloma
Cancer Gene Therapy (2023)
-
CXCL13 chemokine is a novel player in multiple myeloma osteolytic microenvironment, M2 macrophage polarization, and tumor progression
Journal of Hematology & Oncology (2022)
-
BTK inhibitors in the treatment of hematological malignancies and inflammatory diseases: mechanisms and clinical studies
Journal of Hematology & Oncology (2022)
