
Abstract
The HMG-box transcription factor Sox2 plays a role throughout neurogenesis1 and also acts at other stages of development2, as illustrated by the multiple organs affected in the anophthalmia syndrome caused by SOX2 mutations3,4,5. Here we combined proteomic and genomic approaches to characterize gene regulation by Sox2 in neural stem cells. Chd7, a chromatin remodeling ATPase associated with CHARGE syndrome6,7, was identified as a Sox2 transcriptional cofactor. Sox2 and Chd7 physically interact, have overlapping genome-wide binding sites and regulate a set of common target genes including Jag1, Gli3 and Mycn, genes mutated in Alagille, Pallister-Hall and Feingold syndromes, which show malformations also associated with SOX2 anophthalmia syndrome or CHARGE syndrome8,9,10. Regulation of disease-associated genes by a Sox2-Chd7 complex provides a plausible explanation for several malformations associated with SOX2 anophthalmia syndrome or CHARGE syndrome. Indeed, we found that Chd7-haploinsufficient embryos showed severely reduced expression of Jag1 in the developing inner ear.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout




Similar content being viewed by others
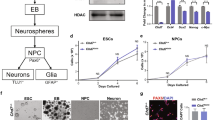
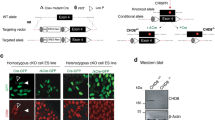
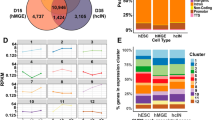
Accession codes
References
Pevny, L.H. & Nicolis, S.K. Sox2 roles in neural stem cells. Int. J. Biochem. Cell Biol. 42, 421–424 (2010).
Guth, S.I. & Wegner, M. Having it both ways: Sox protein function between conservation and innovation. Cell. Mol. Life Sci. 65, 3000–3018 (2008).
Williamson, K.A. et al. Mutations in SOX2 cause anophthalmia-esophageal-genital (AEG) syndrome. Hum. Mol. Genet. 15, 1413–1422 (2006).
Fantes, J. et al. Mutations in SOX2 cause anophthalmia. Nat. Genet. 33, 461–463 (2003).
Kelberman, D. et al. Mutations within Sox2/SOX2 are associated with abnormalities in the hypothalamo-pituitary-gonadal axis in mice and humans. J. Clin. Invest. 116, 2442–2455 (2006).
Vissers, L.E. et al. Mutations in a new member of the chromodomain gene family cause CHARGE syndrome. Nat. Genet. 36, 955–957 (2004).
Zentner, G.E., Layman, W.S., Martin, D.M. & Scacheri, P.C. Molecular and phenotypic aspects of CHD7 mutation in CHARGE syndrome. Am. J. Med. Genet. A. 152A, 674–686 (2010).
Okuno, T., Takahashi, H., Shibahara, Y., Hashida, Y. & Sando, I. Temporal bone histopathologic findings in Alagille's syndrome. Arch. Otolaryngol. Head Neck Surg. 116, 217–220 (1990).
van Bokhoven, H. et al. MYCN haploinsufficiency is associated with reduced brain size and intestinal atresias in Feingold syndrome. Nat. Genet. 37, 465–467 (2005).
Kang, S., Graham, J.M. Jr., Olney, A.H. & Biesecker, L.G. GLI3 frameshift mutations cause autosomal dominant Pallister-Hall syndrome. Nat. Genet. 15, 266–268 (1997).
Gontan, C. et al. Exportin 4 mediates a novel nuclear import pathway for Sox family transcription factors. J. Cell Biol. 185, 27–34 (2009).
van den Berg, D.L. et al. An Oct4-centered protein interaction network in embryonic stem cells. Cell Stem Cell 6, 369–381 (2010).
Hurd, E.A. et al. Loss of Chd7 function in gene-trapped reporter mice is embryonic lethal and associated with severe defects in multiple developing tissues. Mamm. Genome 18, 94–104 (2007).
Alavizadeh, A. et al. The Wheels mutation in the mouse causes vascular, hindbrain, and inner ear defects. Dev. Biol. 234, 244–260 (2001).
Visel, A., Thaller, C. & Eichele, G. GenePaint.org: an atlas of gene expression patterns in the mouse embryo. Nucleic Acids Res. 32, D552–D556 (2004).
Hu, Q. et al. The EGF receptor-sox2-EGF receptor feedback loop positively regulates the self-renewal of neural precursor cells. Stem Cells 28, 279–286 (2010).
Bani-Yaghoub, M. et al. Role of Sox2 in the development of the mouse neocortex. Dev. Biol. 295, 52–66 (2006).
Favaro, R. et al. Hippocampal development and neural stem cell maintenance require Sox2-dependent regulation of Shh. Nat. Neurosci. 12, 1248–1256 (2009).
Taranova, O.V. et al. SOX2 is a dose-dependent regulator of retinal neural progenitor competence. Genes Dev. 20, 1187–1202 (2006).
Howard, T.D. et al. Mutations in TWIST, a basic helix-loop-helix transcription factor, in Saethre-Chotzen syndrome. Nat. Genet. 15, 36–41 (1997).
Bajpai, R. et al. CHD7 cooperates with PBAF to control multipotent neural crest formation. Nature 463, 958–962 (2010).
Schnetz, M.P. et al. Genomic distribution of CHD7 on chromatin tracks H3K4 methylation patterns. Genome Res. 19, 590–601 (2009).
Schnetz, M.P. et al. CHD7 targets active gene enhancer elements to modulate ES cell-specific gene expression. PLoS Genet. 6, e1001023 (2010).
Roessler, E. et al. Loss-of-function mutations in the human GLI2 gene are associated with pituitary anomalies and holoprosencephaly-like features. Proc. Natl. Acad. Sci. USA 100, 13424–13429 (2003).
Bosman, E.A. et al. Multiple mutations in mouse Chd7 provide models for CHARGE syndrome. Hum. Mol. Genet. 14, 3463–3476 (2005).
Avilion, A.A. et al. Multipotent cell lineages in early mouse development depend on SOX2 function. Genes Dev. 17, 126–140 (2003).
Kiernan, A.E. et al. The Notch ligand Jagged1 is required for inner ear sensory development. Proc. Natl. Acad. Sci. USA 98, 3873–3878 (2001).
Li, L. et al. Alagille syndrome is caused by mutations in human Jagged1, which encodes a ligand for Notch1. Nat. Genet. 16, 243–251 (1997).
Brooker, R., Hozumi, K. & Lewis, J. Notch ligands with contrasting functions: Jagged1 and Delta1 in the mouse inner ear. Development 133, 1277–1286 (2006).
Kiernan, A.E. et al. Sox2 is required for sensory organ development in the mammalian inner ear. Nature 434, 1031–1035 (2005).
Shaw-Smith, C. Oesophageal atresia, tracheo-oesophageal fistula, and the VACTERL association: review of genetics and epidemiology. J. Med. Genet. 43, 545–554 (2006).
Conti, L. et al. Niche-independent symmetrical self-renewal of a mammalian tissue stem cell. PLoS Biol. 3, e283 (2005).
Sun, Y. et al. Long-term tripotent differentiation capacity of human neural stem (NS) cells in adherent culture. Mol. Cell. Neurosci. 38, 245–258 (2008).
Ivanova, N. et al. Dissecting self-renewal in stem cells with RNA interference. Nature 442, 533–538 (2006).
Soler, E. et al. The genome-wide dynamics of the binding of Ldb1 complexes during erythroid differentiation. Genes Dev. 24, 277–289 (2010).
Hou, J. et al. Gene expression-based classification of non-small cell lung carcinomas and survival prediction. PLoS ONE 5, e10312 (2010).
Kiernan, A.E. et al. ENU mutagenesis reveals a highly mutable locus on mouse chromosome 4 that affects ear morphogenesis. Mamm. Genome 13, 142–148 (2002).
Sakai, K. & Miyazaki, J. A transgenic mouse line that retains Cre recombinase activity in mature oocytes irrespective of the cre transgene transmission. Biochem. Biophys. Res. Commun. 237, 318–324 (1997).
Nowak, D.E., Tian, B. & Brasier, A.R. Two-step cross-linking method for identification of NF-κB gene network by chromatin immunoprecipitation. Biotechniques 39, 715–725 (2005).
Jothi, R., Cuddapah, S., Barski, A., Cui, K. & Zhao, K. Genome-wide identification of in vivo protein-DNA binding sites from ChIP-Seq data. Nucleic Acids Res. 36, 5221–5231 (2008).
Bailey, T.L. & Elkan, C. The value of prior knowledge in discovering motifs with MEME. Proc. Int. Conf. Intell. Syst. Mol. Biol. 3, 21–29 (1995).
Acknowledgements
We thank G. Abelo for advice on the otocyst stainings, A. Smith for 46C ES cells, S. Pollard for advice on deriving neural stem cells, Z. Ozgür for micro-array hybridizations, M. van den Hout-van Vroonhoven for Illumina GAP analyses and P. Wade for Mi2-β antibody. R.A.P., E.E. and U.A. were supported by a Vidi grant, ALW-open program grant and a Chemical Sciences ECHO grant, respectively, all from the Netherlands Organisation for Scientific Research (NWO). J.C.B. was supported by EuTRACC, B.L. was supported by grants from the Norwegian Research Council (YFF) and the Bergen Research Foundation. C.G. and R.J.R. were supported in part by the Sophia Foundation for Medical Research. S.B. was supported by a British Heart Foundation Chair Award (CH/09/003) and Project Grant award (PG/08/045/25069).
Author information
Authors and Affiliations
Contributions
E.E. and U.A. performed nearly all experiments and analyzed the data. J.C.B. and B.L. normalized the ChIP-Seq data and performed all bioinformatic analyses. J.H. and S.P. normalized and formatted the microarray gene expression data. C.G., R.A.P. and R.J.R. created the F-Sox2 embryonic stem cells. D.S. and S.B. assisted in the mouse work, M.M. performed the GST pull down experiment, C.K. and W.v.IJ. performed the microarray analyses and Illumina sequencing of the ChIP material. D.H.W.D. and J.D. performed the mass spectrometry analyses. E.-J.R. provided bioinformatic assistance in the early stages of this work. L.H.P. provided Sox2 COND mice. F.G.G. set up the ChIP sequencing facility and the bioinformatics infrastructure. R.J.R. created Sox2+/− mice from Sox2 COND mice. R.A.P. designed the study, analyzed the data and wrote the manuscript with support from coauthors.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–7 and Supplementary Tables 1–3 and 11–13. (PDF 10090 kb)
Supplementary Table 4
List of genes regulated by Sox2 and Chd7, identified by microarrays. (XLS 64 kb)
Supplementary Table 5
Genome-wide Sox2 peaks, identified by ChIP-sequencing. (XLS 848 kb)
Supplementary Table 6
Sox2 peaks within 10 kb of genes. (XLS 1359 kb)
Supplementary Table 7
List of genes that are activated and bound by Sox2. (XLS 119 kb)
Supplementary Table 8
Genome-wide Chd7 peaks, identified by ChIP-sequencing. (XLS 2599 kb)
Supplementary Table 9
Chd7 peaks within 10 kb of genes. (XLS 3279 kb)
Supplementary Table 10
List of genes that are activated and bound by Sox2 and Chd7. (XLS 39 kb)
Rights and permissions
About this article
Cite this article
Engelen, E., Akinci, U., Bryne, J. et al. Sox2 cooperates with Chd7 to regulate genes that are mutated in human syndromes. Nat Genet 43, 607–611 (2011). https://doi.org/10.1038/ng.825
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ng.825
This article is cited by
-
Identification of the DNA methylation signature of Mowat-Wilson syndrome
European Journal of Human Genetics (2024)
-
Combined pituitary hormone deficiency harboring CHD7 gene missense mutation without CHARGE syndrome: a case report
BMC Endocrine Disorders (2023)
-
Targeting the SOX2/CDP protein complex with a peptide suppresses the malignant progression of esophageal squamous cell carcinoma
Cell Death Discovery (2023)
-
MZF1 mediates oncogene-induced senescence by promoting the transcription of p16INK4A
Oncogene (2022)
-
CHD7 regulates otic lineage specification and hair cell differentiation in human inner ear organoids
Nature Communications (2022)
