
Abstract
The bed nucleus of the stria terminalis (BNST) is a center of integration for limbic information and valence monitoring. The BNST, sometimes referred to as the extended amygdala, is located in the basal forebrain and is a sexually dimorphic structure made up of between 12 and 18 sub-nuclei. These sub-nuclei are rich with distinct neuronal subpopulations of receptors, neurotransmitters, transporters and proteins. The BNST is important in a range of behaviors such as: the stress response, extended duration fear states and social behavior, all crucial determinants of dysfunction in human psychiatric diseases. Most research on stress and psychiatric diseases has focused on the amygdala, which regulates immediate responses to fear. However, the BNST, and not the amygdala, is the center of the psychogenic circuit from the hippocampus to the paraventricular nucleus. This circuit is important in the stimulation of the hypothalamic–pituitary–adrenal axis. Thus, the BNST has been largely overlooked with respect to its possible dysregulation in mood and anxiety disorders, social dysfunction and psychological trauma, all of which have clear gender disparities. In this review, we will look in-depth at the anatomy and projections of the BNST, and provide an overview of the current literature on the relevance of BNST dysregulation in psychiatric diseases.
Similar content being viewed by others
Introduction
The bed nucleus of the stria terminalis (BNST), also referred to as the BST,1 is fast becoming a relevant region of interest with regard to human stress-related psychiatric disease. This is due to its established role in sustained fear states,2 and social attachment behaviors,3 which comprise aggressive behaviors,4, 5 initiation of mating3 and offspring and parental bonding3, 6 but additionally due to its diverse anatomy, connectivity and receptor subpopulations. Human stress-related psychiatric diseases such as anxiety, post-traumatic stress disorder (PTSD) and social dysfunctions all display long-term changes in mood,7 arousal,8 sleep,7, 8 appetite9 and interest in social interactions.7, 8, 9, 10 The BNST processes information and readiness for response to a threat, by maintaining online information from a vast connectivity network. Mood state and arousal are processed via connections to the amygdala,2, 11 dorsal raphe,12 ventral tegmental area (VTA)13, 14 and medial prefrontal cortex (mPFC).15 The BNST’s projections to the hypothalamus allow monitoring of feeding, drinking and fluid maintenance via brainstem connections.16 Connections to the lateral septum17 and medial amygdala (MeA)11, 17 allow coordination of reproductive behaviors as part of the social behavioral network. Since anatomically, the BNST is sexually dimorphic,18 it may explain the gender disparity in the prevalence and treatment of stress-related psychiatric diseases19, 20 and as such should be investigated as a possible target for treatments. Indeed, drug targets already involve subpopulations of receptors abundant in the BNST such as serotonin.21, 22, 23 While research into the BNST’s role in sustained fear is well-established2 and critical in the treatment of human anxiety, the BNST has further promise in understanding other human-related psychiatric diseases. This is the first review to incorporate several roles of this region in both rodent and human data to promote a united view of the BNST. This is essential in the study of psychiatric diseases such as anxiety, which are part of a larger umbrella of long-term anxiety disorders such as generalized anxiety disorder (GAD), PTSD, social anxiety and anti-social behaviors. This review outlines the complex anatomy of the BNST together with its diversity and interconnectivity. It then discusses the role of the BNST in stress-related psychiatric disease. It concludes with a discussion on the importance of gender disparities in these psychiatric disorders and the possible role of the BNST. These topics aim to help the reader gauge the profound promise of the BNST as a potential target for the treatment of stress-related psychiatric diseases.
BNST’s vs amygdala’s role in fear and anxiety
In rodents, the BNST is a center for integration of information with negative valence or anxiety-like states due to the vastly studied role of corticotrophin-releasing factor (CRF) containing projections to the amygdala, which releases CRF from the central amygdala (CeA) that go on to activate the hypothalamic–pituitary–adrenal (HPA) axis.2, 24 Previous research has established the BNST’s role in sustained fear (which we will term here 'anxiety') as distinct from the amygdala’s role in phasic fear, (which we will refer to as 'fear').2 Studies have suggested that the amygdala is responsible for mediating specific cue-based fear responses, and thus controls the assessment of immediate or phasic fear. The basolateral amygdala receives sensory input25 about a threat and passes the information on to the CeA, which has the capacity to modify the HPA axis.26 Fear conditioning studies have indicated that the BNST’s action is related to the ability to differentiate between safe and non-safe contexts,27 which suggests BNST activity depends on contextual cue reminders that do not predict immediate threat, but rather cause a sustained state of apprehension.2 Blanchard & Blanchard’s28 earlier work on defensive and anxiety-like behaviors in rodents has suggested that fear and anxiety can be divided into an immediate threat, for example, the presence of a predator, and pre- or post-encounter threats, for example, the apprehension of encountering a predator again. The BNST is hypothesized to mediate these longer-term responses to anxiety, where a challenge or recovery from a challenge to homeostasis occurs. In contrast, the CeA is thought to mediate shorter duration phasic fear responses, which occur in response to a current challenge to homeostasis.2
The amygdala’s response to threat is highly individual and often critical to the perception of the threat.29, 30 The role of the amygdala cannot be ignored as it can predispose an individual to an exaggerated stress response when a threat is encountered. Admon et al.29 showed that new army recruits (that is, yet to engage in combat) with more responsive amygdalae were more likely to have greater stress symptoms after a traumatic event. Whalen et al.30 demonstrated hyper-responsiveness to faces in the amygdala of PTSD patients.
In contrast, studies in both male and female patients with phobias, which cause anxiety states, show functional magnetic resonance imaging (fMRI) activation of the BNST, anterior cingulate cortex and insula in anticipation to visual stimuli of phobia-inducing objects (such as spiders), with no activation in the amygdala.31 Another study in which a real Tarantula spider was used during brain imaging in healthy male and female participants, found that the closer the Tarantula was placed to the foot of the participant, the greater the activity in the BNST, but also in the amygdala.32 This suggests diffuse states of threat apprehension are represented differently to the presence of an actual threat; while only the BNST is active during imagery of a future threat, both the BNST and amygdala are activated when faced with an actual threat.
Other models used in human fMRI studies include models of unpredictable shock vs predictable shock or loss of control, in which a threat is anticipated. In both female and male adults, greater BNST activation was found during periods when unpredictable shock was expected, in contrast to the amygdala, which was activated in predictable shock context.33 Using a different model of unpredictability, Somerville et al.34 created an experiment in which participants were asked to manipulate parameters on the monitor. When the line on the screen fell below threshold the participant was shocked. However, in reality the participants’ keyboard was not connected to the computer. Authors found that in both sexes BNST activation was greater when the threat of impeding shock was looming and loss of control was gauged.34
Games involving high monetary uncertainty increased activation in the BNST seen with fMRI BOLD in GAD patients compared with controls.35 In another study, in which the BNST was activated, there was a significant correlation with anxiety levels, while only weak amygdala activation was observed.36
The BNST therefore mediates the anticipatory phase of a threat as well as the general apprehension due to threat and loss of control to prevent the threat. However, when an actual physical threat is present both the BNST and amygdala will be activated.
BNST’s vs amygdala’s role in social behavior
Social decisions requiring the initiation of positive or negative interactions are generated via interactions between the highly interconnected nodes in the social network such as the vomeronasal cavity, the MeA, VTA, several nuclei of the hypothalamus such as the preoptic area, anterior hypothalamus and the ventromedial hypothalamus.37, 38 Convergence of sensory input and limbic networks outputs about mood, homeostasis and reward occur in the BNST, as previously mentioned.38 The BNST, and in particular, its medial and posterior regions, are directly linked to the social behavioral network that regulates behaviors including aggression and defensive behaviors, social communication, mating and parental care.3, 4, 6, 38, 39 In particular, social information in rodents is transferred to the BNST via inputs from the vomeronasal organ to the olfactory bulb and project bilaterally to either the lateral septum or MeA.38, 40 Partner preference and copulation behaviors are also heavily mediated by the BNST with bilateral connections to the MeA.37, 38, 41 After the sensory information to the BNST is processed; the BNST and other nodes in the network can initiate an appropriate social behavioral response.
The MeA mediates social memory and social recognition via its direct connections with the olfactory bulb. Oxytocin in the MeA was found to be crucial and sufficient for social recognition.3, 42 Manipulations of the MeA, such as lesion or stimulation can in males, impair copulation and reduce contact with familiar females,37, 43 and in females mediate an opposite-sex odor preference.44, 45 Lesions of the vomeronasal organ do not change c-Fos expression in the BNST,46 suggesting a dissociation between the two areas. The BNST is involved in precopulatory motivation behaviors for finding a mating partner in both males and females voles,37, 43 and opposite-sex odor preference in female golden hamsters.44, 45 In males, this is demonstrated by increases in BNST androgen receptor expression and testosterone rises in response to receptive females precopulation, as in other nodes of the social network. However, in contrast to other nodes, there is no drop in postcopulation androgens and testosterone levels in the BNST as occurs in MeA after ejaculation.47 In addition, males lose attraction to female odors following posterior medial BNST lesions, though the ability to discriminate between social and non-social odors remain.48 In females, evidence shows that oxytocin receptors in the BNST are responsible for vaginal marking to solicit males and lesions abolish preference for solicitation to males over females.48, 49
Finally, in songbirds, BNST neurons can show differential activation to positive and negative social attraction, affiliation or competition, respectively, based on accessibility to a mate.50 Vasopressin (AVP) immunoreactivity in the BNST is detected when a potential mate or existing partner is present.50 Higher levels of AVP immunoreactivity are seen in social species in which partners are present in flocking vs anti-social species.50 Unlike the well-investigated role of the BNST in fear and anxiety, the role of the social BNST is still emerging as distinct from the amygdala. From the above studies, we can conclude that while the two areas are highly interconnected, specific lesion studies have demonstrated that the role of the BNST involves positive valence, preference and motivation for sexual behaviors that are distinct from mere recognition of opposite-sex conspecifics in context.
The integrated role of the BNST: 'valence surveillance'
To properly understand the seemingly diverse properties of the BNST such as sustained fear and social behaviors, the two roles attributed to the BNST must be reconciled. The BNST has long been understood to assess contextual threat,27 fear-potentiation51 or unconditioned fear.52 However, these studies only used single male mice in a non-social fear conditioning system or startle apparatuses, which enables monitoring safety from a physical threat from the environment. However, negative social encounters can also be considered a physical threat and predator stress or apprehension toward a negative encounter can cause anxiety,28 a socially based type. Therefore, the BNST collectively may assess sensory information from the physical/social environment. Sensory information may include conspecific or interspecies individuals in that context. In addition internally, the BNST may assess the current mood and arousal, which would determine the physical readiness. Together, this accumulation of information will be used to make decisions about the proper response to changes in the physical environment or in the social setting. For example, the return to an inhabitable shelter or the scent of an aggressive conspecific both represent a unspecified threat that can initiate the stress response in the absence of an immediate threat. Alternatively, certain contexts that do not require fight or flight, might be rewarding due to the presence of food or possible mating and BNST-mediated systems will initiate interaction. Therefore, opposing BNST circuits maintained by distinct, opposing sub-nuclei and neurotransmitter systems, as suggested in the study by Daniel et al.,53 may regulate these shifts from homeostasis to stress and vice versa53 depending on the conditions and BNST activation patterns.
We therefore propose a unified role for the BNST in the surveillance and assignment of valence to the information collected. The integrated information can then be used to shift from homeostasis to fight or flight with the help of the BNST’s control of the activation or attenuation of the HPA axis.54 In addition, autonomic control of food, fluid intake and micturition13, 16 are adjusted for the proper response, if necessary. Alternatively, connections with the reward and pain systems may monitor positive valence and adjust information on mood via projections with serotonergic centers.12 The term 'valence surveillance' unifies both the apprehension and social roles of the BNST. In fact, its role in every situation, whether social or not, is to assess the valence either negatively or positively and then, together with the mood, energy and motivation status input coming from relevant brain areas, act appropriately. While extensive anatomical research has been conducted on the BNST, the functionality of each subregion and its connectivity has yet to be dissected systematically with respect to the coordination of these behaviors.
BNST: anatomy in rodents
To better understand how the BNST can integrate information from multiple brain areas to perform 'valence surveillance,' the diversity of the sub-nuclei, projections and subpopulations of this medial forebrain area must be examined. This review will weave through the majority of sub-nuclei and provide an outline for firstly their projections; outputs and inputs and then their subpopulations as they relate to the overall integrative role of the BNST.
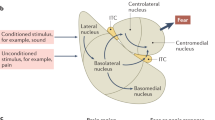
There are several nomenclatures used in the anatomy of the BNST, of which we explain the anatomy according the series of papers of Dong & Swanson11, 13, 16, 17, 24, 55, 56, 57, 58, 59 in which anterograde injections were performed on male rats. The BNST is located ventral to the septum, above and below the anterior commissure and anterior to the hypothalamus. It is enclosed on both sides by the globus pallidus of the basal ganglia. In rodents, the BNST is comprised of up to 18 subregions60 as anatomical techniques and nomenclature are updated. The BNST is primarily divided into anterior and posterior divisions. The anterior division (Figures 1I, 1II and 2II) includes the following well-accepted subregions, anterolateral, anteromedial, oval, fusiform, juxtacapsular, rhomboid, dorsomedial, ventral nucleus, magnocellular,60 in which each has distinct projections within the BNST and outside of the BNST.56, 57, 58, 59, 61 The posterior division (Figures 1III and 2III) is comprised of three well-characterized nuclei: the principal, the interfascicular and the transverse.13 Figure 3 shows a heatmap summary of the BNST’s receptors, neurotransmitters, transporters/proteins in 12 of its sub-nuclei, adapted from the study by Bota et al.60

Afferent projections of the rodent BNST. Diverse brain areas project to the different sub-nuclei of the BNST and often to several sub-nuclei (adapted from refs 10, 11, 12, 13, 23). (I) Purple arrows represent incoming projections to the anterolateral (al), anteromedial (am) and ventral (fu) areas of the BNST at the level of the anterior commissure (ac). (II) Orange arrows represent projections to the anterolateral (al) and anteromedial (am) BNST areas posterior to the anterior commissure (ac). (III) Green arrows represent projections to the posterior BNST comprising of three nuclei: the principle (pr), the interfascicular (if) and the transverse (tr). 3v, third ventricle; ac, anterior commissure; al, anterolateral BNST; am, anteromedial BNST; Amy, amygdala; BNST, bed nucleus stria terminalis; d, dorsal nucleus; dm, dorsomedial nucleus; DR, dorsal raphe; FC, frontal cortex; fu, fusiform nucleus; GP, globus pallidus; hypo, hypothalamus; if, interfascicular nucleus; ju, juxtacapsular nucleus; LC, locus coeruleus; LS, lateral septum; mg, magnocellular nucleus; MPO, medial preoptic area; NTS, nucleus solitary tract; OB, olfactory bulb; ov, oval nucleus; pr, principle nucleus; PVN, paraventricular nucleus; rh, rhomboid nucleus; st, striatum; tr, transverse nucleus; VS, ventral subiculum; VTA, ventral tegmental area.

Efferent projections of the rodent BNST. The BNST sends diverse and unique projections from different subregions (adapted from refs 10, 11, 12, 13, 23). (I) Purple arrows represent outgoing projections from the anterolateral (al), anteromedial (al) and ventral (fu) areas of the BNST at the level of the anterior commissure (ac). (II) Orange arrows represent projections from anterolateral and medial BNST areas posterior to the anterior commissure (ac). (III) Green arrows represent projections from the posterior BNST comprising of three nuclei: the principle (pr), the interfascicular (if) and the transverse (tr). 3v, third ventricle; ac, anterior commissure; al, anterolateral BNST; am, anteromedial BNST; Amy, amygdala; BNST, bed nucleus stria terminalis; d, dorsal nucleus; dm, dorsomedial nucleus; DR, dorsal raphe; FC, frontal cortex; fu, fusiform nucleus; GP, globus pallidus; hypo, hypothalamus; if, interfascicular nucleus; ju, juxtacapsular nucleus; LC, locus coeruleus; LS, lateral septum; mg, magnocellular nucleus; MPO, medial preoptic area; NTS, nucleus solitary tract; OB, olfactory bulb; ov, oval nucleus; pr, principle nucleus; PVN, paraventricular nucleus; rh, rhomboid nucleus; st, striatum; tr, transverse nucleus; VS, ventral subiculum; VTA, ventral tegmental area.

Heatmap representing subpopulations of receptors, neurotransmitters and transporters in all subregions of the BNST. Adapted from the study by Bota et al.60 with the following genes added: DARPP32,71 CGRP,71 VIP,71 PAC1,63 PACAP,63, 64 CRFR1,81 CRFR2,67, 81 PR,68, 69 A1-Adr,62, 96 A2-Adr,62, 65 B1-Adr,53, 62, 65 B2-Adr.62, 65 The original data were extracted from the study by Bota et al.60 which categorized the expression levels from 0 to 9, with 12 having no data. In our version, we binned all the genes previously published and newly added from 0 to 3, 4 to 6, 6 to 9 and no data into 4 ranks of strong, moderate, weak expression and no data to combine all the data into the qualitative expression heatmap. A1-Adr, adrenergic receptor alpha-1; A2-1 GABA A, GABRA2, GABA a2 receptor; A2-Adr, adrenergic receptor alpha-2; A3-1 GABA A, GABA3, GABA a3 receptor, B1-1 GABA A, GABA beta 1 receptor; AII, angiotensin I; AR, androgen receptor; AVP, vasopression; B1-Adr, adrenergic receptor beta 1; B2-Adr, adrenergic receptor beta 2; B3-1 GABA A, GABA beta 3 receptor; CaB, calbindin; CB-1R, cannabinoid receptor 1; CCK, cholecystokinin; CGRP, calcitonin gene-related peptide; ChAT, choline acetyltransferase; CR, calretinin; CRFR1, cortiotrophin releasing factor receptor type 1; CRFR2, corticotrophin-releasing factor, receptor type 2; D1, Drd1A, dopamine receptor type 1A; D2, Drd2A, dopamine receptor type 2A; DARPP32, dopamine- and cAMP-regulated phosphoprotein; DOR, delta opioid receptor; DYN, dynorphin; ENK, enkelphalin; ER, estrogen receptors; G1-1 GABA A, GABA, gamma 1 receptor; GAD65, glutamate decarboxylase 65; GAD67, glutamate decarboxylase 67; GAL, galanin; GluR7, GRIK3, kainate receptor; GlurR5, GRIK1 kainate receptor; Htr1A, 5Ht1A, serotonin receptor type 1; KA2, GRIK 5, kainate receptor; KOR, kappa-opioid receptor; mGluR2, metabotropic glutamate receptor 2; mGluR7A, metabotropic glutamate receptor 7A; mGluR7B, metabotropic glutamate receptor 7B; MOR, mu-opioid receptor; nChR, nicotinic acetylcholine receptor; NK3, tachykinin receptor 3; NKB, neurokinin B; NPY, neuropeptide Y; NPY1, neuropeptide Y receptor Y1; NPY2, neuropeptide Y receptor Y2; NPY4, neuropeptide Y receptor Y4; NPY5, neuropeptide Y receptor Y5; NT, neurotensin; Oxt-1A, oxytocin receptor 1A; Oxt-1B, oxytocin receptor 1B; PAC1, Adcyap1r1, pituitary adenylate cyclase-activating polypeptide type 1; PACAP, pituitary adenylate cyclase-activating polypeptide; PR, progesterone receptor; SOM, somatostatin; SP, substance P; VGLUT1, vesicular glutamate transporter 1; VGLUT2, vesicular glutamate transporter 2; VGLUT3, vesicular glutamate transporter 3; VIP, vasoactive intestinal peptide.
Anterior division
The oval nucleus integrates information about mood and negative valence via CRF, GABA, pituitary adenylate cyclase-activating polypeptide (PACAP), dopamine, encephalin, while promoting anxiety-like behavior.14 GABAergic projections from the oval nucleus reach the CeA, VTA and the lateral hypothalamus.24 CeA GABAergic neurons project to the lateral areas both dorsal and ventral to the anterior commissure, with a subset co-expressing CRF,62 which respond differentially to the CRF-expressing neurons of the fusiform nucleus.53 CRF mRNA levels in the dorsal BNST, now understood to refer to the oval nucleus, are increased after chronic stress and foot shock unlike the ventral BNST.53 PACAP and its receptor PAC1 are found in the oval nucleus and interact with CRF-positive neurons.63, 64 Dopamine receptors in the oval nucleus receive input from the VTA as well as from the dorsal raphe.65 Optogenetic stimulation of a population of dopaminergic receptor-labeled neurons in the oval nucleus were anxiogenic while anterolateral (dorsal area) stimulation was anxiolytic through receiving differential input from the basolateral amygdala.62 Enkephalinergic neurons from the globus pallidus project to the oval nucleus as well as the anterolateral, rhomboid and juxtacapsular areas.66 The presence of enkephalin in close proximity to oval CRF neurons may assist in the shift between negative to positive valence and between opposing roles of stress induction or return to homeostasis based on the valence assessment.
The anterolateral BNST, located just ventral of the oval nucleus, is an area where many of the distinct subpopulations responsible for shifting BNST roles meet. Convergence of steroid hormone receptors, oxytocin receptors, regulators such as dopamine- and cAMP-regulated phosphoprotein (DARP), serotonin receptors, anxiety-regulation by CRF receptor type 1 (CRFR1) GABAergic opioid regulation as well as HPA axis modulating connections occurs in this diverse sub-nuclei. The incoming information received may imply the integration possibilities about mating, bonding, neuroendocrine state, with mood and pain pathway processing.
The anterolateral region sends GABAergic projections to neuroendocrine centers of the paraventicular area of the hypothalamus13 and receives inputs from the ventral subiculum’s glutamatergic projections,62 a connection involved in negative feedback of the HPA axis.67 Connections also include reciprocal dorsal raphe serotonergic inputs12 and outputs.13 Steroid hormone receptors such as alpha and beta estrogen receptors,68, 69 androgen receptors as well as aromatase68, 69 and oxytocin receptors are located in the dorsal areas of the anterolateral BNST.70 Co-localized populations of DARP32, vasoactive intestinal peptide (VIP), tyrosine hydroxylase and calcitonin gene-related peptide are also abundant.71 Interestingly, mu-opioid and kappa-opioid receptors on GABAergic interneurons in both the oval and anterolateral areas including rhomboid and juxtacapsular areas synapse on CRFR1 receptors affecting synaptic transmission.72, 73 The inputs to the anterolateral region can therefore assess whether an anxiety-like experience awaits or a positive bonding is on the horizon. Subsequent outputs to the hypothalamus and raphe regions may influence the behavioral outcome.
While the fusiform, juxtacapsular and rhomboid nuclei have not been studied in-depth functionally, understanding their parallel projections to the same neurocircuitry but different nuclei within the anxiety, reward and pain network may provide useful insights into the BNST’s integration capabilities.
The fusiform nucleus projects to the CeA, paraventricular nucleus (PVN), nucleus accumbens, peri-aqueductal gray, which is involved in pain,74 and reticular nuclei,61 which is involved in attention. The juxtacapsular nucleus, in contrast projects both to the CeA, basolateral amygdala, substantia nigra and dorsal raphe.11, 61 In other words, their projections are to similar circuitry but to different nodes of each circuitry. What this may mean in terms of type of quality of information transferred is very intriguing. A high number of CRF-positive axons are found in the ventral lateral regions and CRF mRNA is highly expressed in both the oval and fusiform nuclei,54 as stated previously as well as mu-opioid, kappa-opioid receptors and enkephalin immunoreactive cells.66, 72, 73
The rhomboid nucleus projects to the CeA as well as important centers all over the brain ranging from mesolimbic reward centers such as the VTA to the substantia nigra, nucleus accumbens and globus pallidus.13 In addition, there are also projections to several areas of the hypothalamus including the PVN, and medial part of the preoptic hypothalamus, reticular nucleus and peri-aqueductal gray.13 This sub-nuclei contains GABA, glutamate, as well as dynorphins,73 part of the opioid system, enkephalin immunoreactive cells66 and dopamine receptors.60
The ventral BNST has the largest density of noradrenergic fibers in the brain,62 positioning it as a center of arousal, which is important in assessing appropriate responses to sensory information. The ventral BNST receives inputs from A1 (caudal ventrolateral medulla) and A2 (nucleus of the solitary tract) cell bodies in the brainstem, with a small population originating from the locus coeruleus.62, 65 Stimulation of the fusiform’s vast input from the nucleus of the solitary tract can depress cardiovascular responses.53 Tonic noradrenergic release has also been reported in outputs of the ventral BNST. Noradrenaline is released in response to synthetic fox odor (2,3,5-Trimethyl-3-thiazoline), immobilization stress, foot shock and at the anticipation of a reward that is in the end not received.53, 65, 75 Signaling is via presynaptic alpha-2 adrenergic receptors as well as synergistic facilitation via CRF and alpha-1 adrenergic receptors.62 Overall the ventral BNST is an important center for arousal, which may play an important role in assessing readiness to respond to either positive or negative valence.
The anteromedial areas of the BNST are part of a unique circuitry of stress-regulated autonomic control that receives inputs from medial preoptic area (MPOA),16 and other hypothalamic areas,16 as well as the MeA16 and posterior BNST,16, 76 which presumably can either activate or attenuate the HPA axis peripheral changes based on the olfactory information received.
The anteromedial group projections (see Figure 2I) are distinct from that of the anterolateral, which has more sensory and motor related projections to the vagus nerve controlling digestive homeostasis,16 while the anteromedial projects to Barrington’s nucleus, which is involved in drinking and fluid homeostasis.16 This region has the highest density of direct projections to the PVN of any brain region and also projects directly to the CeA.16 From the PVN, projections descend to the brainstem to regulate pelvic functions via the Barrington’s nucleus.16 Interaction with the PVN and digestive homeostasis is most likely critical for HPA axis response and fight or flight implementation, in which digestive activity is ebbed.77 This nuclei is also enlarged after antenatal glucocorticoid exposure.78
The dorsomedial BNST may be central for the integration of social information, and if necessary activation of the HPA axis to initiate fight or flight behaviors and adjustment of mood and reward circuitry. This region is located ventral to the anterior commissure and medial to the fusiform nucleus. This subregion has the most extensive direct projections to the hypothalamus.55 It additionally projects to AVP neurons in the supraoptic region, to oxytocin neuronal cell bodies in the magnocellular region of the PVN and to the median preoptic nucleus.55 It also projects to parvocellular PVN neurons that produce thyrotropin-releasing hormone, CRF, somatostatin and gonadotropin-releasing hormone.55 Projections to the PVN are mostly GABAergic and innervate the parvocellular and magnocellular neurons of the PVN. Progesterone receptors are found in the ventral medial BNST.68, 69
The dorsomedial nucleus (see Figure 2II) receives input from the basomedial amygdala, MeA and the CeA, as well as the ventral subiculum of the hippocampus, the insula and infralimbic prefrontal regions.55 Inputs to the dorsomedial nucleus also include dopaminergic projections from the VTA, serotonergic neurons from the raphe nuclei and noradrenergic neurons from the locus coeruleus and nucleus of the solitary tract.55 Inputs from the ventral subiculum, the output of the hippocampus, are primarily glutamatergic.62 Lesion of this subregion attenuates HPA axis activity.54
Both the dorsomedial and magnocellular neurons also project to preautonomic neurons in PVN, which project to the spinal cord and Barrington’s nucleus which innervates the bladder and the colon.55, 56 Therefore, these nuclei might play a role in integration of information necessary for neuroendocrine and autonomic control of voluntary drinking behavior, fluid balance, sodium regulation, especially micturition, defection and erection.56 Many of these behaviors are affected as part of the flight or fight response, but prolonged stress or chronic CRF may also dysregulate autonomic systems.79
Posterior BNST
The posterior division of the BNST is comprised of three nuclei: the principal, the interfascicular and the transverse (see Figure 1III and Figure 2III).17 The posterior BNST receives GABAergic input from the dorsal lateral septum, medial CeA and MeA posterior dorsal and MeA posterior ventral regions.37, 76 Glutamatergic input to the posterior BNST comes from the paraventricular thalamus, basomedial amygdala, cortical amygdala, CA1 of the hippocampus and ventral subiculum.80 This division is also rich in CRF type 2 receptors (CRFR2).67, 81 The posterior region of the BNST is sexually dimorphic with distinct alpha and beta estrogen receptors as well as androgen populations.68, 69 AVP receptors are located in the posteriolateral region70 and often interact with androgen receptors.
Connections of the principal nucleus may play a role in inhibition of other medial BNST areas involved in HPA axis activation based on assigned valence information obtained from the social behavioral network connections. The principal nucleus innervates the septum17 and many hypothalamic areas involved in reproduction and defensive behaviors in rodents.17, 82 These areas include the MPOA, pre-mammillary bodies, ventromedial hypothalamus and PVN.17 The principal nucleus has high expression of GAD65 (a GABA-synthesizing enzyme) mRNA.83 The posterior part also has many intra-BNST projections mainly to medial areas such as the magnocellular nuclei and dorsomedial BNST.17 It has also been shown to attenuate the HPA axis.54, 67
The transverse and intrafasicular nuclei send projections to mesolimbic brain regions such as the VTA and substantia nigra17 and receive projections from the anterior olfactory bulb and MeA, suggesting a role in social recogntion.44 In addition, fibers from the MeA pass through the BNST via the transverse and intrafascicular nucleus to innervate the septum.17 The transverse nucleus alone innervates the PVN, in particular the parvocellular stress-regulating neurons, which are GABAergic.17
To conclude, the posterior region’s connectivity suggests its involvement in reproduction and social defensive behaviors via modulation of the PVN. Additional interplay with the anteromedial BNST nuclei,17 via the principal nucleus, enables posterior division to maintain homeostasis in response to the stress of survival and HPA responsiveness. Based on the above evidence, interactions with the VTA may provide integration on motivation to engage in social behaviors after identification of the social benefits of the encounter by other social network circuitry. Activation of the HPA axis could be dependent on the outcome of the decision-making process of valence attachment to the encounter.
BNST: anatomy in humans
While in rodents the BNST’s anatomy and connectivity have been well-characterized, functional connectivity in humans has been addressed only recently. Using post-mortem tissue, the anatomy of human BNST has been mapped immunohistochemically.84 However, recent technological advancement such as improvement in magnetic field strength, allow imaging in higher spatial resolution of 7 T,85 which has helped visualize this small area. Emerging fMRI studies have described significant activation in the BNST accompanying anticipatory anxiety.31, 34
The human BNST can be divided into lateral, medial, central84 and ventral subdivisons (see Figure 4). The lateral, medial and central regions are grouped together anteriorly at the level of the anterior commissure (see Figure 4, anterior panel) becoming smaller and joining the ventral region more posteriorly (see Figure 4, posterior panel). Like in rodents, it is a sexually dimorphic structure, with human male brains having a BNST 2.5 times greater in volume than females.18 In a recent report three distinct, ipsilateral fiber pathways of the human BNST were described: the posterior bundle, the ventral bundle and the anterior bundle.86 The posterior bundle connects the BNST via the stria terminalis to the lateral amygdala. The ventral bundle connects the BNST through the ansa peduncularis to the MeA and hypothalamus. Both of these pathways match those found in animal models.87 However, the anterior pathway has not been reported previously. The anterior bundle runs through the head of the caudate body and nucleus accumbens to the mPFC and the orbitofrontal cortex.86 The authors also reported that BNST connections to the brainstem were subject to substantial inter-individual variability.

Anatomy and subdivisions (red text) of the human anterior BNST (left panel) and posterior BNST (right panel). The orange dots depict the network of brain areas with either anatomical connection or which have been implicated in its functional connectivity. Image adapted from Atlas of the Human Brain (http://www.thehumanbrain.info). ac, anterior commissure; Acc, nucleus accumbens; BLA, basal lateral nucleus of the amygdala; BM, basal medial nucleus of the amygdala; BNST, bed nucleus of the stria terminalis; BNSTc, bed nucleus of the stria terminalis central subdivision; BNSTL, bed nucleus of the stria terminalis lateral subdivision; BNSTm, bed nucleus of the stria terminalis medial subdivision; BNSTv, bed nucleus of the stria terminalis ventral subdivision; Cd, caudate; DB, diagonal band; Ent, entorhinal cortex; FPu, putaminal fundus region; GPe, globus pallidus external; Ic, internal capsule; LA, lateral nucleus of the anygdala; LH, lateral hypothalamus; LS, lateral septum; mPFC, medial prefrontal cortex; MPO, medial preoptic nucleus; oc, optic chiasm; PirF, piriform cortex; Pu, putamen; PVN, paraverntricular nucleus; Sch, suprachiasmatic nucleus; VP, ventral pallidum; vPFC, ventral prefrontal cortex.
FMRI scans of rhesus monkeys and young humans show functional resting state connectivity between the CeA and BNST signal88 and a significant likelihood of structural connectivity to the CeA as well as to the temporal pole, a paralimbic area.89 The human temporal pole is reported to be involved in the coupling of high level sensory processing and emotions. For example, lesions can cause intolerance of auditory stimuli coupled with increases in contextual fear, or loss of link between faces and names, which causes social deficits.90 In addition, the BNST was shown to be functionally connected to the thalamus, insula and mPFC in an emotional task requiring the subject to distinguish between safe and non-safe contexts.91 FMRI studies also show functional connectivity of the BNST with the ventromedial prefrontal cortex (vmPFC), with vmPFC-lesion patients showing decreased blood flow in the right BNST.92 The vmPFC has been previously associated with arousal and emotion evoking stimuli.93 An additional, functional connection was established with the paracingulate gyrus, also part of the prefrontal cortex and thought to be involved in anxiety and addiction.85 With enhanced imaging capabilities, functional connectivity studies and fMRI studies mapping higher cognitive functions will hopefully emerge to help us understand the neural circuitry of psychiatric BNST-related disorders.
Psychiatric disorders and their treatments: involvement of the BNST
Anxiety
An anxiety disorder is diagnosed when a patient presents with 'excessive worry' or negative valence about a variety of situations or contexts and difficulty in controlling these thoughts, which in turn may cause difficulty sleeping and concentrating or irritability.7 These symptoms affect the daily functioning of an individual over a sustained period of time.9 Anxiety disorders are treated using anti-depressants such as selective serotonin reuptake inhibitors (SSRIs), selective norepinephrine reuptake inhibitor (SNRIs) or benzodiazepines, which facilitate GABA transmission (less commonly used due to dependence problems).2, 7, 22
Currently available anti-depressants, which increase serotonin levels, either activate the BNST chronically or activate behaviors that are BNST-mediated. In rodents, as consistent with the BNST’s proposed role in 'valence surveillance,' projections to the anterolateral BNST have the greatest serotonergic innervation in the brain from the caudal dorsal raphe and can shift mood based on the specific serotonergic receptor types activated. Anxiogenic vs anxiolytic behaviors depend on the specific serotonergic receptor type activation23 and the BNST contains many serotonin receptors which are the major targets of anxiety disorder treatments: Htr1A, Htr2A, Htr2C and Htr7.12, 94 Htr1A autoreceptors mediate the inhibitory response of the BNST to serotonin. In contrast, Htr2A and Htr2C mediate anxiogenic behavior via depolarization in response to serotonin.12, 94 Co-localized interactions between this serotonergic population and CRF may also play a role in the antidepressant effects described above12 in particular in the oval nucleus.22 The BNST also contains alpha and beta adrenergic receptors and subtypes, 53, 62 possibly involved in arousal state and may be the reason for the efficacy in some patients of SNRIs.
GABAeric transmission, the target of benzodiazepines used to treat anxiety, includes abundant projections from the anteromedial BNST to the PVN.62, 95 Changes in GABA receptors in the dorsomedial and principal nuclei under chronic stress conditions have been demonstrated.96 It is also these two areas that regulate the activation or attenuation of the HPA axis, respectfully54, 67, 97 therefore possibly shifting anxiogenic states to anxiolytic states. In addition, the BNST–VTA GABAergic pathway reduces anxiety,98 however, activation of this connection while effective, may also cause dependence and explain the reduction in popularity of benzodiazepine treatment in favor of SSRIs.
While many rodent and human studies look at the effect of both acute and chronic SSRI treatments on fear and anxiety, in reality, treatment involves long-term administration.22 In male rats, acute systemic fluoxetine, a type of SSRI, treatment increases Arc, an intermediate early gene, expression in the dorsal lateral areas of BNST and in the CeA.23 Local infusions of acute fluoxetine initially exacerbate anxiety-like behavior and conditioned fear memories in the BNST but not in the CeA.23 Chronic fluoxetine, reduces stress-induced BNST c-Fos99 and inhibits sustained fear, but not phasic fear.100
Sustained fear in general is susceptible to clinical intervention with SSRIs, making the study of anxiety disorders, their pharmacology and the BNST more relevant for clinical success.2 Acute tryptophan depletion meant to model low serotonin levels, in human subjects increases eyeblink startle, a BNST-mediated paradigm, to context but not cued stimuli.101 The above data demonstrate not only the BNST’s role in anxiety, but its role in the responsiveness to SSRI treatments, which require shifts from anxiogenic to anyxiolytic after chronic administration.
Chronic SSRI treatment was effective in both rodents and humans in models of sustained fear such as light- and dark-enhanced startle as well.2 Startle studies of sustained fear in both male rodents and male humans show that acute administration of citalopram, an SSRI, increases fear and anxiety in response to shocks, while chronic administration reduces contextual anxiety but not cued fear.2 Human studies show reduction in startle with chronic benzodiazepines administration while rodent studies demonstrated an anxiolytic effect.2, 100
With an 18% prevalence rate of people who experience anxiety for a 12-month period,102 research on the BNST, known to mediate extended fear states in both humans and male rodents should be a priority for future pharmaceutical treatments. While anxiety has been the most intensively researched BNST-based psychiatric disorder, research is yet to encapsulate the full complexity of the shifts between anxiogenic vs anxiolytic states in BNST nuclei-specific neurotransmitter systems and connections.
Post-traumatic stress disorder
PTSD is a severely debilitating psychiatric condition characterized by persistent symptoms of intrusive re-experience, avoidance and hyperarousal months to years following exposure to a traumatic event.8, 103 PTSD patients show sustained fear behaviors attributed to the BNST, which may imply impaired 'valence surveillance.' Neurobiological changes in PTSD patients reflect shifts in integration of arousal, HPA axis activation and mood state that may impair realistic decisions about threat. Levels of cortisol, a heavily debated biomarker in PTSD,104 are also mediated by output from the BNST.54 Hyperarousal is mediated by noradrenergic projections to the BNST62 and negative valence by sertonergic inputs abundant in the BNST.12 The high comorbidity of drug abuse with PTSD103 also implies shifts in mood and appetite for increasing positive mood states.
PTSD patients, in particular, exhibit measurable sustained fear as demonstrated by startle and loss of control paradigms. Similar to patients with GAD, the most effective clinical treatments today are SSRIs and SNRIs. Grillion et al.105 show increased sustained fear in PTSD patients, but not in GAD patients, suggesting involvement of the BNST. Furthermore, two studies which measured single nucleotide polymorphisms (SNPs) in blood samples of PTSD patients, showed SNPs of genes also expressed by BNST neurons.106, 107 Interestingly, in both studies the effects were specific to female PTSD patients. Female PTSD patients show a significant association of SNPs for CRFR2, and PAC1 (ADCYAP1R1) with risk to develop PTSD and with severity of PTSD symptoms.106, 107 White, non-hispanic females cohabitating with traumatized combat veterans with PTSD were found to have significantly reduced lifetime risk of developing PTSD, associated with two CRFR2 SNPs.106 One SNP was also associated with severity of PTSD.106
In animal models, a possible mechanism for sustained fear is, in fact, the CRFR2, which mediates coping with challenges to homeostasis. The role of coping is actually an important component of the BNST’s proposed role of 'valence surveillance' in that coping requires physiological shifts based on new sensory information collected. While CRF/CRFR1 systems are involved in initiation of the stress response, CRFR2 involves turning off stressful responses when the stressor is no longer present.108 Knockdown of CRFR2, specifically in the BNST of male mice was also shown to cause changes in anxiety-like behaviors 24 or 48 h after restraint, but not right after (Lebow et al., unpublished data). Furthermore, CRFR2KD in the BNST was demonstrated to be involved in increased arousal and vigilance67 and robust changes in CRFR2 mRNA were exhibited in the BNST of PTSD-like mice that underwent unpredictable shock.67 The effects of CRFR2 are also proven to be sex-specific, with only CRFR2 female knockout mice showing increased anxiety-like and depressive-like behaviors.109 As such, BNST CRFR2 could represent a future target for human psychopharmacology in which initiation of downstream coping genes could possibly reduce PTSD symptoms of sustained fear and shift negative valence to positive valence. Furthermore, the BNST’s CRFR2s may have sex-specific effects in psychiatric disorders.
PACAP is another rodent and human target that may hold promise for the future treatment of PTSD, in a sex-specific manner, given it is heavily expressed in the BNST. PACAP is localized in the oval nucleus of the rodents BNST, where it is known to innervate CRF-expressing neurons12 and where it has been shown to cause increased anxiety states.63 BNST PACAP also stimulates activation of the HPA axis.110 Continuous blockade of PAC1 during chronic stress reverses this effect.110 A single human SNP in the PAC1 (ADCYAP1R1) gene, which is located in an estrogen responsive element, was recently found to be a predictor for PTSD and to be associated with severity of PTSD symptoms in a highly traumatized cohort of African-American women. Furthermore, methylation of this gene in patient blood was found to correlate with total PTSD symptom load only in women.107
Human psychiatric diseases such as anxiety and PTSD are also accompanied by changes in cortisol levels. Changes in cortisol are also a function of the proposed BNST’s role of 'valence surveillance,' in that information about the environment must be processed to activate the stress response or to turn it off. PTSD patients show reduced cortisol and this is mirrored in PTSD-like animal models which show reduced corticosterone levels.67, 111, 112, 113, 114, 115 Accordingly lesions of the posterior medial BNST in male rats cause increases in corticosterone levels 30 min after restraint stress, while lesions of the anterior ventral BNST attenuate corticosterone levels immediately after restraint stress.83, 97 The anterior BNST serves as a relay station between the mPFC and the PVN that mediate HPA responses to stress15 to activate the HPA axis and causes corticosterone release, while the posterior BNST inhibits the HPA axis and attenuates corticosterone release.54, 83
SSRIs are considered the most accepted treatment for PTSD symptoms in patients, and other treatments all target serotonin, dopamine, norepinephrine and GABAergic systems though not always most effectively,116 and self-administration of alcohol is also rampant.103 While the most effective treatment is yet to be discovered, current treatments are consistent with a mouse model of PTSD, in which, vast molecular changes take place in serotoergic, adrenergic and even GABAergic systems in the BNST. Downregulation of receptor mRNA is seen for adrenergic (Adra2a, Adrbk1), GABAergic (Gabra2, Gabra3) and serotonergic (Htr1A, Htr2c, Htr7) receptors.67 In addition, a metabolite of serotonin, 5-HIAA, showed decreased levels 2 weeks after the unpredictable shock paradigm and immediately following measurement of the startle response (personal communication, Christopher Lowry, University of Colorado), indicating alteration in the BNST serotonin system in a rodent PTSD model. This might possibly explain the mechanism of SSRI treatment in PTSD patients who have increased sustained fear.
PTSD itself is a disorder that is twice as prevalent in females than males. As reviewed above, the human literature shows sex-specific changes in PTSD patients in SNPs of genes expressed in neurons in BNST subpopulations. Overall, this makes the BNST an excellent target for future research in PTSD and treatment for PTSD.
Aggression and anti-social behaviors
Aggression is defined as behavior intended to inflict physical damage on another individual.4 In humans, aggression can be differentiated into two types; impulsive aggression in response to anger, often called reactive aggression, and goal-oriented aggression, which is referred to as controlled instrumental aggression.4 Reactive aggression in humans is often the result of PTSD or depression, which can be due to increased arousal combined with feelings of helplessness and loss of control. Controlled instrumental aggression in humans is found in anti-social personality types who show low arousal and often have emotionally blunted responses.4 Pharmaceutical drugs used to treat both types of aggression in humans often involve sedatives or antipsychotics, which target the serotonin system.117
Psychopathy or other anti-social behaviors characterized by low empathy and aggression are the result of low arousal and impaired fear processing.4 Many human studies on psychopathy focus on impaired fear processing in the amygdala and showed reduced amygdala activation in fMRI of psychopaths.5 However, cold calculated long-term decision-making behavior seems to be outside the realm of the amygdala and rather a frontal cortex process.
The prefrontal cortex is thought to mediate aggression and impulse control.4 SSRI treatment in borderline personality disorder patients increased activity in the prefrontal cortex, ameliorating the lack of inhibitory control.4 Patients with vmPFC lesions show blunted arousal to emotional stimuli and reduced BNST activity was observed.93 Research on anti-social behavior may benefit by the further study into the frontal cortex and ventral BNST connections, which could regulate controlled thinking in anti-social behaviors and influence low arousal states and aggressive behavior, respectively.
At least in rodents, the BNST’s proposed role as 'valence surveillance' may be important for indicating the presence of an aggressor and whether to fight or flight. This is based on the relay of information from the mPFC to the BNST and from BNST to the PVN, in which the mPFC inhibits the ventral BNST to activate the medial parvocellular PVN.15 The decision may also integrate BNST noradrenergic information to determine or increase readiness. The noradrenergic population of the BNST could therefore be an important target for treatment of aggression via its functional connectivity with the prefrontal cortex.
The ability to engage in aggressive behavior is also necessary in rodents to obtain resources such as food or mates. These behaviors are mediated by a vast network of sensory and limbic information. Olfactory cues transmitted via the social behavior network mediate aggressive behaviors and lesions in any of these nodes or in the frontal cortical regions can greatly reduce aggressive behavior between males.40 Other systems thought to biologically mediate aggression involve vasopressinergic,4 dopaminergic75, 98, 118 GABAergic,4, 39, 118 noradrenergic,4 and steroid hormones and receptors,4, 119 which are all abundant in the sub-nuclei of the BNST. Aggressive behavior is mediated by and correlated with vasopressinergic–androgen-dependent projections from the BNST to the lateral septum.119 There are positive correlations with estrogen alpha positive neurons in the BNST, as well as AVP immunoreactivity in the BNST and aggression.119 The most aggressive rodents also show the highest oxytocin receptor binding, in medial and posterior sections of the BNST.120 Aggression is also attributed to the ventromedial hypothalamus which is highly connected has projections from the principal nucleus.4, 17 As a result, the BNST and its subpopulation of AVP and steroid hormone receptors mediate the initiation of aggression in rodents after sensory evaluation, while connections with the frontal cortex mediate changes in homeostasis in response to the sensory information, further implicating the BNST’s integration of information for use in implementing responses to homeostatic challenges.
Social attachment disorders: social anxiety and motivation of bond formation
The BNST is implicated in the neurobiology of social attachment behavior in rodents and humans,3, 121 which includes behaviors such as mating between conspecifics and parental behaviors. Impairments in these BNST-mediated mating behaviors may also play a role in human social dysfunction such as social anxiety, autism or inadequate parental care, making a person less likely to coordinate behaviors to form or maintain the social bonds necessary for reproductive behaviors. Three stages are critical to complete the social attachment bonds; inhibition of fear, social recognition and reward motivation.3 Social anxiety disorder occurs in 6.8% of the population and is described by a fear of unfamiliar people who may scrutinize the sufferer in a social situation, which causes anxiety and social avoidance in the sufferer.122 One of the consequences of social anxiety disorder on an individual is the lack of ability to initiate a social bond out of fear and result in insecurity and increased stress.122 In addition, there is a lack of sufficient motivation and increases in inhibition of initiation of these behaviors.122 While the amygdala and frontal cortex may be crucial for social recognition and inhibition of fear,3 the BNST through its connections with the VTA3, 98 and frontal cortex3 and MPOA3 plays an important role in motivation for mating and anticipation of reward via dopamine and oxytocin circuitry3 or as hypothesized attaching positive valence to the process based on 'surveillance' information received.
Mating behavior in rodents involves a coordination of complex precopulatory behaviors such as solicitation behavior and odor discrimination to facilitate a successful outcome.3, 48 Complex connectivity between the brain circuits containing dopamine, oxytocin, noradrenaline, GABA and glutamate such as the VTA, BNST and MPOA may explain these behaviors.
Within the BNST itself, a dissociation exists between dopaminergic, GABAergic release in the oval nucleus vs noradrenergic and glutamatergic release in the ventral BNST based on presentation of a rewarding cue or anticipation of a rewarding cue, respectively.65, 123 While these systems have mostly been studied in the context of drug addiction and relapse,124 motivation for mating or anticipation of a mate, may work through this same circuitry. In fact, dopamine is known to be involved in pair bonding and pair bonding in social species is considered to be rewarding.3 Females monogamous voles have increased BNST oxytocin receptors, which are expressed in both dorsal and ventral areas of the BNST.60 There is also sex-specific expression of the dopamine D1 receptor in the lateral BNST in which males have stronger D1 receptor immunoreactivity.125 It is hypothesized that oxytocin receptor activation in the BNST and in the VTA via MPOA release, as well as dopamine release from the VTA, may mediate motivation to pair bond and monogamy after the initial rewarding experience is encoded by nucleus accumbens and VTA.3 In the BNST, increases in glutamate in response to the rewarding experience also change plasticity and hence motivation to obtain it.123
There are reciprocal GABAergic connections from the BNST to the VTA. In addition, VTA to BNST projections are glutamatergic and have glutamate transporters, suggesting that the BNST disinhibits VTA dopaminergic neurons, yielding a rewarding state.126 Optogenetic stimulation of BNST–VTA circuitry provides evidence of BNST GABAergic connections causing rewarding states and reduction in anxiety-like states, while BNST glutamatergic VTA connections are anxiogenic.98 Therefore, the BNST mediates the interplay between anxiety, anticipation and reward that comprises social attachment and the vicious cycle of substance abuse. In fact several articles have addressed the topic of the idea that social attachment is a form of addiction.3, 124 More research using BNST-mediated dopamine, oxytocin, CRF, glutamate and GABA in social behavior could elucidate the circuitry of motivation for a mate and maintaining monogamy.
Inadequate parental attachment and offspring care
Parental behavior is also considered part of the reward, motivation and addiction-like circuitry, involving the BNST. Dopamine plays a critical role in the rewarding aspect, but the BNST oxytocin and CRFR2 play a role in response to care.3, 127
In humans, the brain of new mothers and experienced mothers show increased activity in fMRI in both the ventral BNST and amygdala in response to one’s infant crying.121 The amygdala was additionally activated in other conditions of general emotionally arousing noise such as cries and laughter of other babies, thus demonstrating responses to phasic changes in emotional stimuli. However, in response to visual stimuli of one’s infant, the BNST showed increased activity in contrast to the amygdala.121 This may indicate that BNST activation occurs in response to both emotionally valent auditory and visual stimuli with respect to one’s infant and not general emotionally arousing information.
In rodents, receptor subpopulations of the posterior BNST, such as CRFR2 and oxytocin receptors, are also thought to be involved in maternal care. Hypoactivation in the BNST of CRFR2 is important for maternal care during lactation.127 Furthermore, oxytocin receptor levels in the BNST, CeA, MPOA and lateral septum are correlated with higher frequency of licking and grooming of pups.3 The human BNST is involved in assessing valence regarding offspring,121 presumably to act to protect the offspring and activate systems necessary to do so depending on the valence levels. In rodents, changes in posterior BNST circuitry and its projections determine lactation127 and pup care.3
Sex differences and steroid hormones
Any study of the neurobiology of human psychiatric diseases would be incomplete without mentioning the gross discrepancy in the prevalence and treatment outcome between the sexes. Women are two times more likely than men to have a major depressive episode and more likely to be diagnosed with an anxiety disorder.20, 102 There are also differential success rates in women vs men in the treatment of depression, with men being more responsive to imipramine and paroxetine.19 Women are also twice as likely to develop PTSD after exposure to a trauma128 and as previously discussed sex-specific SNPs were found for female PTSD patients.106, 107
In addition, the BNST itself, contains subpopulations of receptors for steroid hormones such as androgen, estrogen (both alpha and beta subtypes) as well as progesterone,6, 47, 68, 119 which may interact differently with states of sustained fear and anxiety in females vs males and influence 'survellience' and attachment of proper valence to situations.
In rodents, with regard to anxiety-related behaviors, steroid hormone manipulation affects behavior specifically in BNST-dependent paradigms. Progesterone reduces CRF-enhanced startle. Progesterone also activates BNST neurons via oxytocin.129 Intact males have the lowest light-potentiated startle and pregnant females the highest.69 Testosterone reduces light enhanced startle in male rats only.
Anatomically, many projections and subpopulation sex differences can be found in the BNST. The human female BNST has 76% greater structural connectivity in regions that had significant connectivity with the BNST.130 In rodents, the AVP pathway from the BNST to the lateral septum is larger in males131, 132 and gonadectomy of hypogonadism reduces BNST AVP immunoreactivity.133, 134 Testosterone exposure causes an increase in prenatal changes of CRF immunoreactive neurons in the BNST of ovariectomized rats. This area was more vulnerable than the MPOA during sexual differentiation.135 The bilateral connections between the posterior BNST and the MeA may also be involved in sexual differentiation. While MeA to posterior BNST connections appear by embryonic day 14, reciprocal projections from the BNST to the MeA appear by postnatal day 15 during the critical postnatal period suggesting the necessity of steroid hormones in completion of these circuits.136 Anatomical and behavioral differences between the sexes in the BNST or anxiety-related receptors and behavior might help explain the vast discrepancy between prevalence rates of human psychiatric diseases between men and women.
Summary
We propose that the BNST plays the essential role of 'valence surveillance' by processing salient information taken from the monitoring of the physical and social contexts. Decisions about threatening and rewarding situations are accessed while information on mood, homeostatic state, arousal, feeding and reproduction is kept online. Anatomically, the BNST is heterogeneous, containing many sub-nuclei and an abundant diversity of neurotransmitters and their receptors. Understanding the subpopulations and specific connectivity of this diverse area is critical for understanding its role in pathology and designing therapeutic interventions. While much research is focused on understanding human psychiatric diseases by using animal models to study the amygdala, the literature has somewhat overlooked the role of the BNST, which might explain their complexity. Figure 5 illustrates the proposed roles of the human BNST and dysfunction associated with impaired BNST function. Human psychiatric diseases are not only isolated to immediate altered fear processing but may include vast changes in long-term anxiety states, arousal, social and reproductive behaviors, in addition to the often overlooked different manifestations among the sexes. Therefore, increased understanding of the BNST is sure to provide profound translational insights for treatments of human psychiatric diseases.

Summary representing the behaviors that the BNST regulates and implicated pathology in event of dysfunction of connectivity or neurotransmitter populations. BNST, bed nucleus of the stria terminalis.
Future directions
The purpose of this review is to raise awareness of the plethora of research that is yet to be performed and the translational relevance of studying the BNST. BNST animal research has thus far only focused on anatomical projections, the startle circuitry and on social behaviors. Often in this research, the BNST is not the main focus of the study, but rather added as an afterthought and most are performed in non-rodent and primate species. Advanced models of psychiatric disorders in rodents unfortunately focus on more well-characterized brain areas and as such the 'state of the art' genetic toolbox is reserved for better-understood areas.
For example, anatomical tracing has benefitted by advances in technology. Combining CRE and flox lines137 and injections of herpes, an anterograde tracer,138 or rabies, a retrograde tracer,139 would give anatomical information regarding specific subpopulations projections within the known BNST circuitry. Functionally, CRE lines bred with flox lines can conditionally knockout a specific gene, from which its function can be accessed via behavioral changes in its absence. Changes in activation and subsequent changes in behavior can also be demonstrated using optogenetics in CRE lines in specific brain nuclei. The teasing out of BNST circuitry in this way has begun in several recent optogenetic BNST-focused papers.14, 41, 73, 98, 140 There are many permutations for research using these techniques in that the target projections from the BNST can be illuminated and tested for changes in behavior as well as the BNST sub-nuclei that is sending the projections. At the behavioral level, more BNST-specific paradigms are needed aside from the startle test, which tests for sustained fear. Due to the social nature of the BNST, biological markers such as HPA axis or receptor changes of any of the relevant subpopulations should be measured after behaviors in a group setting of same sex or after interactions of both sexes. Manipulations introducing stressors like shock or reward would be interesting as they affect the individual responses but in a group setting would enhance the understanding of the BNST’s proposed role of 'valence surveillance' in a more ethological setting.
Moreover, the latest advances in epigenetics, including methylation, acetylation, microRNAs, other small non-coding RNAs, RNA editing and other bioinformatics pathways from RNAseq often do not generate BNST-specific data. Arrays of the genome and micronome of the BNST in general may show unique patterns after acute or chronic stress or the BNST-specific paradigms to be generated, as discussed above. Individually methylated regions based on behavioral experiences may also provide information about the role of adaptivity in specific BNST sub-nuclei to environmental exposures. Again and again the challenge is specific dissection between anterior regions.
The entire field of these potential biomarkers from whole blood or plasma that correspond to changes in BNST receptor or neurotransmitter populations has yet to be established in rodents or humans. mRNA or microRNA levels in the BNST could be compared with plasma circulating levels to see if BNST levels are predictive enough for use as biomarkers in psychiatric diseases. Studies involving animal models of psychiatric disease in both females and males are also missing in most brain research, but there is greater rationale for doing so for the BNST. This has left a large gaping hole in the field of the psychiatric research in animal models.
While human research is also in its infancy, some strides have been made in deep brain stimulation specifically for the BNST.141, 142 Other than directly stimulating the BNST, studying its behavioral-induced fMRI signal as well as the general BNST functional connectivity during different states and in different pathologies is of great interest. Post-mortem tissue studies looking at gene expression changes may bring forward candidate genes for biomarker diagnostics of psychiatric disorders. Overall, there is still a vast amount of research needed to understand the full complexity of the BNST.
Change history
07 November 2019
A Correction to this paper has been published: https://doi.org/10.1038/s41380-019-0568-0
References
Dumont EC . What is the bed nucleus of the stria terminalis? Prog Neuropsychopharmacol Biol Psychiatry 2009; 33: 1289–1290.
Davis M, Walker DL, Miles L, Grillon C . Phasic vs sustained fear in rats and humans: role of the extended amygdala in fear vs anxiety. Neuropsychopharmacology 2010; 35: 105–135.
Coria-Avila GA, Manzo J, Garcia LI, Carrillo P, Miquel M, Pfaus JG . Neurobiology of social attachments. Neurosci Biobehav Rev 2014; 43: 173–182.
Nelson RJ, Trainor BC . Neural mechanisms of aggression. Nat Rev Neurosci 2007; 8: 536–546.
Marsh AA . What can we learn about emotion by studying psychopathy? Front Hum Neurosci 2013; 7: 181.
Frazier CRM, Trainor BC, Cravens CJ, Whitney TK, Marler CA . Paternal behavior influences development of aggression and vasopressin expression in male California mouse offspring. Horm Behav 2006; 50: 699–707.
Allgulander C . Generalized anxiety disorder: between now and DSM-V. Psychiatr Clin North Am 2009; 32: 611–628.
Chu JA . Posttraumatic stress disorder: beyond DSM-IV. Am J Psychiatry 2010; 167: 615–617.
Brown TA, Campbell LA, Lehman CL, Grisham JR, Mancill RB . Current and lifetime comorbidity of the DSM-IV anxiety and mood disorders in a large clinical sample. J Abnorm Psychol 2001; 110: 585–599.
Hare RD, Hart SD, Harpur TJ . Psychopathy and the DSM-IV criteria for antisocial personality disorder. J Abnorm Psychol 1991; 100: 391–398.
Dong HW, Petrovich GD, Swanson LW . Topography of projections from amygdala to bed nuclei of the stria terminalis. Brain Res Brain Res Rev 2001; 38: 192–246.
Hammack SE, Guo J-D, Hazra R, Dabrowska J, Myers KM, Rainnie DG . The response of neurons in the bed nucleus of the stria terminalis to serotonin: implications for anxiety. Prog Neuropsychopharmacol Biol Psychiatry 2009; 33: 1309–1320.
Dong H-W, Swanson LW . Organization of axonal projections from the anterolateral area of the bed nuclei of the stria terminalis. J Comp Neurol 2004; 468: 277–298.
Kim S-Y, Adhikari A, Lee SY, Marshel JH, Kim CK, Mallory CS et al. Diverging neural pathways assemble a behavioural state from separable features in anxiety. Nature 2013; 496: 219–223.
Radley JJ, Sawchenko PE . A common substrate for prefrontal and hippocampal inhibition of the neuroendocrine stress response. J Neurosci 2011; 31: 9683–9695.
Dong H-W, Swanson LW . Projections from bed nuclei of the stria terminalis, anteromedial area: cerebral hemisphere integration of neuroendocrine, autonomic, and behavioral aspects of energy balance. J Comp Neurol 2006; 494: 142–178.
Dong H-W, Swanson LW . Projections from bed nuclei of the stria terminalis, posterior division: implications for cerebral hemisphere regulation of defensive and reproductive behaviors. J Comp Neurol 2004; 471: 396–433.
Allen LS, Gorski RA . Sex difference in the bed nucleus of the stria terminalis of the human brain. J Comp Neurol 1990; 302: 697–706.
Sloan DM, Kornstein SG . Gender differences in depression and response to antidepressant treatment. Psychiatr Clin North Am 2003; 26: 581–594.
Steiner M, Allgulander C, Ravindran A, Kosar H, Burt T, Austin C . Gender differences in clinical presentation and response to sertraline treatment of generalized anxiety disorder. Hum Psychopharmacol 2005; 20: 3–13.
Hammack SE, Cooper MA, Lezak KR . Overlapping neurobiology of learned helplessness and conditioned defeat: implications for PTSD and mood disorders. Neuropharmacology 2012; 62: 565–575.
Burghardt NS, Bauer EP . Acute and chronic effects of selective serotonin reuptake inhibitor treatment on fear conditioning: implications for underlying fear circuits. Neuroscience 2013; 247: 253–272.
Ravinder S, Burghardt NS, Brodsky R, Bauer EP, Chattarji S . A role for the extended amygdala in the fear-enhancing effects of acute selective serotonin reuptake inhibitor treatment. Transl Psychiatry 2013; 3: e209.
Dong HW, Petrovich GD, Watts AG, Swanson LW . Basic organization of projections from the oval and fusiform nuclei of the bed nuclei of the stria terminalis in adult rat brain. J Comp Neurol 2001; 436: 430–455.
LeDoux J . The amygdala. Curr Biol 2007; 17: R868–R874.
Herman JP, Cullinan WE . Neurocircuitry of stress: central control of the hypothalamo–pituitary–adrenocortical axis. Trends Neurosci 1997; 20: 78–84.
Sullivan GM, Apergis J, Bush DEA, Johnson LR, Hou M, Ledoux JE . Lesions in the bed nucleus of the stria terminalis disrupt corticosterone and freezing responses elicited by a contextual but not by a specific cue-conditioned fear stimulus. Neuroscience 2004; 128: 7–14.
Blanchard RJ, Shepherd JK, Rodgers RJ, Magee L, Blanchard DC . Attenuation of antipredator defensive behavior in rats following chronic treatment with imipramine. Psychopharmacology (Berl) 1993; 110: 245–253.
Admon R, Lubin G, Stern O, Rosenberg K, Sela L, Ben-Ami H et al. Human vulnerability to stress depends on amygdala’s predisposition and hippocampal plasticity. Proc Natl Acad Sci USA 2009; 106: 14120–14125.
Whalen PJ, Shin LM, McInerney SC, Fischer H, Wright CI, Rauch SL . A functional MRI study of human amygdala responses to facial expressions of fear versus anger. Emotion 2001; 1: 70–83.
Straube T, Mentzel H-J, Miltner WHR . Waiting for spiders: brain activation during anticipatory anxiety in spider phobics. Neuroimage 2007; 37: 1427–1436.
Mobbs D, Yu R, Rowe JB, Eich H, FeldmanHall O, Dalgleish T . Neural activity associated with monitoring the oscillating threat value of a tarantula. Proc Natl Acad Sci USA 2010; 107: 20582–20586.
Alvarez RP, Chen G, Bodurka J, Kaplan R, Grillon C . Phasic and sustained fear in humans elicits distinct patterns of brain activity. Neuroimage 2011; 55: 389–400.
Somerville LH, Whalen PJ, Kelley WM . Human bed nucleus of the stria terminalis indexes hypervigilant threat monitoring. Biol Psychiatry 2010; 68: 416–424.
Yassa MA, Hazlett RL, Stark CEL, Hoehn-Saric R . Functional MRI of the amygdala and bed nucleus of the stria terminalis during conditions of uncertainty in generalized anxiety disorder. J Psychiatr Res 2012; 46: 1045–1052.
Somerville LH, Fani N, McClure-Tone EB . Behavioral and neural representation of emotional facial expressions across the lifespan. Dev Neuropsychol 2011; 36: 408–428.
Newman SW . The medial extended amygdala in male reproductive behavior. A node in the mammalian social behavior network. Ann NY Acad Sci 1999; 877: 242–257.
Goodson JL, Kabelik D . Dynamic limbic networks and social diversity in vertebrates: from neural context to neuromodulatory patterning. Front Neuroendocrinol 2009; 30: 429–441.
Caldwell HK, Lee H-J, Macbeth AH, Young WS . Vasopressin: behavioral roles of an “original” neuropeptide. Prog Neurobiol 2008; 84: 1–24.
Brennan PA, Keverne EB . Something in the air? New insights into mammalian pheromones. Curr Biol 2004; 14: R81–R89.
Maejima S, Ohishi N, Yamaguchi S, Tsukahara S . A neural connection between the central part of the medial preoptic nucleus and the bed nucleus of the stria terminalis to regulate sexual behavior in male rats. Neurosci Lett 2015; 606: 66–71.
Bielsky IF, Young LJ . Oxytocin, vasopressin, and social recognition in mammals. Peptides 2004; 25: 1565–1574.
Kirkpatrick B, Carter CS, Newman SW, Insel TR . Axon-sparing lesions of the medial nucleus of the amygdala decrease affiliative behaviors in the prairie vole (Microtus ochrogaster): behavioral and anatomical specificity. Behav Neurosci 1994; 108: 501–513.
Petrulis A, Johnston RE . Lesions centered on the medial amygdala impair scent-marking and sex-odor recognition but spare discrimination of individual odors in female golden hamsters. Behav Neurosci 1999; 113: 345–357.
Takahashi LK, Gladstone CD . Medial amygdaloid lesions and the regulation of sociosexual behavioral patterns across the estrous cycle in female golden hamsters. Behav Neurosci 1988; 102: 268–275.
Fernandez-Fewell GD, Meredith M . c-fos expression in vomeronasal pathways of mated or pheromone-stimulated male golden hamsters: contributions from vomeronasal sensory input and expression related to mating performance. J Neurosci 1994; 14: 3643–3654.
He F, Yu P, Wu R . Relationship between sexual satiety and motivation, brain androgen receptors and testosterone in male mandarin voles. Behav Brain Res 2013; 250: 257–263.
Martinez LA, Petrulis A . The bed nucleus of the stria terminalis is critical for sexual solicitation, but not for opposite-sex odor preference, in female Syrian hamsters. Horm Behav 2011; 60: 651–659.
Martinez LA, Levy MJ, Petrulis A . Endogenous oxytocin is necessary for preferential Fos expression to male odors in the bed nucleus of the stria terminalis in female Syrian hamsters. Horm Behav 2013; 64: 653–664.
Goodson JL, Wang Y . Valence-sensitive neurons exhibit divergent functional profiles in gregarious and asocial species. Proc Natl Acad Sci USA 2006; 103: 17013–17017.
Walker DL, Davis M . Light-enhanced startle: further pharmacological and behavioral characterization. Psychopharmacology (Berl) 2002; 159: 304–310.
Walker DL, Davis M . Double dissociation between the involvement of the bed nucleus of the stria terminalis and the central nucleus of the amygdala in startle increases produced by conditioned versus unconditioned fear. J Neurosci 1997; 17: 9375–9383.
Daniel SE, Rainnie DG . Stress modulation of opposing circuits in the bed nucleus of the stria terminalis. Neuropsychopharmacology 2015; 41: 103–125.
Choi DC, Furay AR, Evanson NK, Ostrander MM, Ulrich-Lai YM, Herman JP . Bed nucleus of the stria terminalis subregions differentially regulate hypothalamic-pituitary-adrenal axis activity: implications for the integration of limbic inputs. J Neurosci 2007; 27: 2025–2034.
Dong HW, Swanson LW . Projections from bed nuclei of the stria terminalis, dorsomedial nucleus: Implications for cerebral hemisphere integration of neuroendocrine, autonomic, and drinking responses. J Comp Neurol 2006; 494: 75–107.
Dong HW, Swanson LW . Projections from bed nuclei of the stria terminalis, magnocellular nucleus: Implications for cerebral hemisphere regulation of micturition, defecation, and penile erection. J Comp Neurol 2006; 494: 108–141.
Dong HW, Petrovich GD, Swanson LW . Organization of projections from the juxtacapsular nucleus of the BST: a PHAL study in the rat. Brain Res 2000; 859: 1–14.
Dong HW, Petrovich GD, Swanson LW . Topography of projections from amygdala to bed nuclei of the stria terminalis. Brain Res Rev 2001; 38: 192–246.
Dong HW, Swanson LW . Projections from the rhomboid nucleus of the bed nuclei of the stria terminalis: Implications for cerebral hemisphere regulation of ingestive behaviors. J Comp Neurol 2003; 463: 434–472.
Bota M, Sporns O, Swanson LW . Neuroinformatics analysis of molecular expression patterns and neuron populations in gray matter regions: the rat BST as a rich exemplar. Brain Res 2012; 1450: 174–193.
Dong HW, Swanson LW . Organization of axonal projections from the anterolateral area of the bed nuclei of the stria terminalis. J Comp Neurol 2004; 468: 277–298.
Forray MI, Gysling K . Role of noradrenergic projections to the bed nucleus of the stria terminalis in the regulation of the hypothalamic-pituitary-adrenal axis. Brain Res Brain Res Rev 2004; 47: 145–160.
Hammack SE, Roman CW, Lezak KR, Kocho-Shellenberg M, Grimmig B, Falls WA et al. Roles for pituitary adenylate cyclase-activating peptide (PACAP) expression and signaling in the bed nucleus of the stria terminalis (BNST) in mediating the behavioral consequences of chronic stress. J Mol Neurosci 2010; 42: 327–340.
Kozicz T, Vigh S, Arimura A . Axon terminals containing PACAP- and VIP-immunoreactivity form synapses with CRF-immunoreactive neurons in the dorsolateral division of the bed nucleus of the stria terminalis in the rat. Brain Res 1997; 767: 109–119.
Park J, Bucher ES, Fontillas K, Owesson-White C, Ariansen JL, Carelli RM et al. Opposing catecholamine changes in the bed nucleus of the stria terminalis during intracranial self-stimulation and its extinction. Biol Psychiatry 2013; 74: 69–76.
Arluison M, Vankova M, Cesselin F, Leviel V . Origin of some enkephalin-containing afferents to the ventro-medial region of the globus pallidus in the rat. Brain Res Bull 1990; 25: 25–34.
Lebow M, Neufeld-Cohen A, Kuperman Y, Tsoory M, Gil S, Chen A . Susceptibility to PTSD-like behavior is mediated by corticotropin-releasing factor receptor type 2 levels in the bed nucleus of the stria terminalis. J Neurosci 2012; 32: 6906–6916.
Laflamme N, Nappi RE, Drolet G, Labrie C, Rivest S . Expression and neuropeptidergic characterization of estrogen receptors (ERalpha and ERbeta) throughout the rat brain: anatomical evidence of distinct roles of each subtype. J Neurobiol 1998; 36: 357–378.
Toufexis DJ, Myers KM, Bowser ME, Davis M . Estrogen disrupts the inhibition of fear in female rats, possibly through the antagonistic effects of estrogen receptor alpha (ERalpha) and ERbeta. J Neurosci 2007; 27: 9729–9735.
Stoop R . Neuromodulation by oxytocin and vasopressin. Neuron 2012; 76: 142–159.
Gustafson EL, Greengard P . Localization of DARPP-32 immunoreactive neurons in the bed nucleus of the stria terminalis and central nucleus of the amygdala: co-distribution with axons containing tyrosine hydroxylase, vasoactive intestinal polypeptide, and calcitonin gene-related pept. Exp brain Res 1990; 79: 447–458.
Jaferi A, Pickel VM . Mu-opioid and corticotropin-releasing-factor receptors show largely postsynaptic co-expression, and separate presynaptic distributions, in the mouse central amygdala and bed nucleus of the stria terminalis. Neuroscience 2009; 159: 526–539.
Li C, Pleil KE, Stamatakis AM, Busan S, Vong L, Lowell BB et al. Presynaptic inhibition of gamma-aminobutyric acid release in the bed nucleus of the stria terminalis by kappa opioid receptor signaling. Biol Psychiatry 2012; 71: 725–732.
Bodnar RJ . Endogenous opiates and behavior: 2012. Peptides 2013; 50: 55–95.
Silberman Y, Winder DG . Emerging role for corticotropin releasing factor signaling in the bed nucleus of the stria terminalis at the intersection of stress and reward. Front psychiatry 2013; 4: 42.
Gomez DM, Newman SW . Differential projections of the anterior and posterior regions of the medial amygdaloid nucleus in the Syrian hamster. J Comp Neurol 1992; 317: 195–218.
Romero ML, Butler LK . Endocrinology of stress. Int J Comp Psychol 2007; 20: 89–95.
Oliveira M, Rodrigues A-J, Leão P, Cardona D, Pêgo JM, Sousa N . The bed nucleus of stria terminalis and the amygdala as targets of antenatal glucocorticoids: implications for fear and anxiety responses. Psychopharmacology (Berl) 2012; 220: 443–453.
Mcfadden K, Griffin TA, Levy V, Wolfe JH, Valentino RJ . Overexpression of corticotropin-releasing factor in Barrington’s nucleus neurons by adeno-associated viral transduction: Effects on bladder function and behavior. Eur J Neurosci 2012; 36: 3356–3364.
Myers B, Mark Dolgas C, Kasckow J, Cullinan WE, Herman JP . Central stress-integrative circuits: forebrain glutamatergic and GABAergic projections to the dorsomedial hypothalamus, medial preoptic area, and bed nucleus of the stria terminalis. Brain Struct Funct 2014; 219: 1287–1303.
Van Pett K, Viau V, Bittencourt JC, Chan RK, Li HY, Arias C et al. Distribution of mRNAs encoding CRF receptors in brain and pituitary of rat and mouse. J Comp Neurol 2000; 428: 191–212.
Wood RI, Newman SW . The medial amygdaloid nucleus and medial preoptic area mediate steroidal control of sexual behavior in the male Syrian hamster. Horm Behav 1995; 29: 338–353.
Choi DC, Evanson NK, Furay AR, Ulrich-Lai YM, Ostrander MM, Herman JP . The anteroventral bed nucleus of the stria terminalis differentially regulates hypothalamic-pituitary-adrenocortical axis responses to acute and chronic stress. Endocrinology 2008; 149: 818–826.
Walter A, Mai JK, Lanta L, Görcs T . Differential distribution of immunohistochemical markers in the bed nucleus of the stria terminalis in the human brain. J Chem Neuroanat 4: 281–298.
Avery SN, Clauss JA, Blackford JU . The human BNST: functional role in anxiety and addiction. Neuropsychopharmacology 2015; 41: 126–141.
Krüger O, Shiozawa T, Kreifelts B, Scheffler K, Ethofer T . Three distinct fiber pathways of the bed nucleus of the stria terminalis to the amygdala and prefrontal cortex. Cortex 2015; 66: 60–68.
Weller KL, Smith DA . Afferent connections to the bed nucleus of the stria terminalis. Brain Res 1982; 232: 255–270.
Oler JA, Birn RM, Patriat R, Fox AS, Shelton SE, Burghy CA et al. Evidence for coordinated functional activity within the extended amygdala of non-human and human primates. Neuroimage 2012; 61: 1059–1066.
Avery SN, Clauss JA, Winder DG, Woodward N, Heckers S, Blackford JU . BNST neurocircuitry in humans. Neuroimage 2014; 91: 311–323.
Olson IR, Plotzker A, Ezzyat Y . The Enigmatic temporal pole: a review of findings on social and emotional processing. Brain 2007; 130: 1718–1731.
Kinnison J, Padmala S, Choi J-M, Pessoa L . Network analysis reveals increased integration during emotional and motivational processing. J Neurosci 2012; 32: 8361–8372.
Motzkin JC, Philippi CL, Oler JA, Kalin NH, Baskaya MK, Koenigs M . Ventromedial prefrontal cortex damage alters resting blood flow to the bed nucleus of stria terminalis. Cortex 2015; 64: 281–288.
Motzkin JC, Philippi CL, Wolf RC, Baskaya MK, Koenigs M . Ventromedial prefrontal cortex is critical for the regulation of amygdala activity in humans. Biol Psychiatry 2015; 77: 276–284.
Hazra R, Guo JD, Dabrowska J, Rainnie DG . Differential distribution of serotonin receptor subtypes in BNSTALG neurons: modulation by unpredictable shock stress. Neuroscience 2012; 225: 9–21.
Herman JP, Figueiredo H, Mueller NK, Ulrich-Lai Y, Ostrander MM, Choi DC et al. Central mechanisms of stress integration: hierarchical circuitry controlling hypothalamo-pituitary-adrenocortical responsiveness. Front Neuroendocrinol 2003; 24: 151–180.
Ventura-Silva AP, Pêgo JM, Sousa JC, Marques AR, Rodrigues AJ, Marques F et al. Stress shifts the response of the bed nucleus of the stria terminalis to an anxiogenic mode. Eur J Neurosci 2012; 36: 3396–3406.
Choi DC, Furay AR, Evanson NK, Ulrich-Lai YM, Nguyen MMN, Ostrander MM et al. The role of the posterior medial bed nucleus of the stria terminalis in modulating hypothalamic-pituitary-adrenocortical axis responsiveness to acute and chronic stress. Psychoneuroendocrinology 2008; 33: 659–669.
Jennings JH, Sparta DR, Stamatakis AM, Ung RL, Pleil KE, Kash TL et al. Distinct extended amygdala circuits for divergent motivational states. Nature 2013; 496: 224–228.
Bechtholt AJ, Valentino RJ, Lucki I . Overlapping and distinct brain regions associated with the anxiolytic effects of chlordiazepoxide and chronic fluoxetine. Neuropsychopharmacology 2008; 33: 2117–2130.
Miles L, Davis M, Walker D . Phasic and sustained fear are pharmacologically dissociable in rats. Neuropsychopharmacology 2011; 36: 1563–1574.
Robinson OJ, Overstreet C, Allen PS, Pine DS, Grillon C . Acute tryptophan depletion increases translational indices of anxiety but not fear: serotonergic modulation of the bed nucleus of the stria terminalis? Neuropsychopharmacology 2012; 37: 1963–1971.
Kessler RC, Chiu WT, Demler O, Merikangas KR, Walters EE . Prevalence, severity, and comorbidity of 12-month DSM-IV disorders in the National Comorbidity Survey Replication. Arch Gen Psychiatry 2005; 62: 617–627.
Kessler RC, Sonnega A, Bromet E, Hughes M, Nelson CB . Posttraumatic stress disorder in the National Comorbidity Survey. Arch Gen Psychiatry 1995; 52: 1048–1060.
Fink G . Stress controversies: post-traumatic stress disorder, hippocampal volume, gastroduodenal ulceration. J Neuroendocrinol 2011; 23: 107–117.
Grillon C, Pine DS, Lissek S, Rabin S, Bonne O, Vythilingam M . Increased anxiety during anticipation of unpredictable aversive stimuli in posttraumatic stress disorder but not in generalized anxiety disorder. Biol Psychiatry 2009; 66: 47–53.
Wolf EJ, Mitchell KS, Logue MW, Baldwin CT, Reardon AF, Humphries DE et al. Corticotropin releasing hormone receptor 2 (CRHR-2) gene is associated with decreased risk and severity of posttraumatic stress disorder in women. Depress Anxiety 2013; 30: 1161–1169.
Ressler KJ, Mercer KB, Bradley B, Jovanovic T, Mahan A, Kerley K et al. Post-traumatic stress disorder is associated with PACAP and the PAC1 receptor. Nature 2011; 470: 492–497.
Bale TL, Vale WW . CRF and CRF receptors: role in stress responsivity and other behaviors. Annu Rev Pharmacol Toxicol 2004; 44: 525–557.
Bale TL, Vale WW . Increased depression-like behaviors in corticotropin-releasing factor receptor-2-deficient mice: sexually dichotomous responses. J Neurosci 2003; 23: 5295–5301.
Roman CW, Lezak KR, Hartsock MJ, Falls WA, Braas KM, Howard AB et al. PAC1 receptor antagonism in the bed nucleus of the stria terminalis (BNST) attenuates the endocrine and behavioral consequences of chronic stress. Psychoneuroendocrinology 2014; 47: 151–165.
McFarlane AC, Barton CA, Yehuda R, Wittert G . Cortisol response to acute trauma and risk of posttraumatic stress disorder. Psychoneuroendocrinology 2011; 36: 720–727.
Golier JA, Caramanica K, Demaria R, Yehuda R . A pilot study of mifepristone in combat-related PTSD. Depress Res Treat 2012; 2012: 393251.
Lenze EJ, Mantella RC, Shi P, Goate AM, Nowotny P, Butters MA et al. Elevated cortisol in older adults with generalized anxiety disorder is reduced by treatment: a placebo-controlled evaluation of escitalopram. Am J Geriatr Psychiatry 2011; 19: 482–490.
Neufeld-Cohen A, Kelly PAT, Paul ED, Carter RN, Skinner E, Olverman HJ et al. Chronic activation of corticotropin-releasing factor type 2 receptors reveals a key role for 5-HT1A receptor responsiveness in mediating behavioral and serotonergic responses to stressful challenge. Biol Psychiatry 2012; 72: 437–447.
Michels N, Sioen I, Clays E, De Buyzere M, Ahrens W, Huybrechts I et al. Children’s heart rate variability as stress indicator: association with reported stress and cortisol. Biol Psychol 2013; 94: 433–440.
Shalev AY . Posttraumatic stress disorder and stress-related disorders. Psychiatr Clin North Am 2009; 32: 687–704.
Patrick CJ, Durbin CE, Moser JS . Reconceptualizing antisocial deviance in neurobehavioral terms. Dev Psychopathol 2012; 24: 1047–1071.
Kash TL, Pleil KE, Marcinkiewcz CA, Lowery-Gionta EG, Crowley N, Mazzone C et al. Neuropeptide regulation of signaling and behavior in the BNST. Mol Cells 2015; 38: 1–13.
Trainor BC, Greiwe KM, Nelson RJ . Individual differences in estrogen receptor alpha in select brain nuclei are associated with individual differences in aggression. Horm Behav 2006; 50: 338–345.
Calcagnoli F, de Boer SF, Beiderbeck DI, Althaus M, Koolhaas JM, Neumann ID . Local oxytocin expression and oxytocin receptor binding in the male rat brain is associated with aggressiveness. Behav Brain Res 2014; 261: 315–322.
Bridges R . Neurobiology of the Parental Brain. Elsevier, 2010; https://books.google.com/books?id=5kGGAE90nmYC&pgis=1 (accessed 9 August 2015).
Bögels SM, Alden L, Beidel DC, Clark LA, Pine DS, Stein MB et al. Social anxiety disorder: Questions and answers for the DSM-V. Depress Anxiety 2010; 27: 168–189.
Wills TA, Winder DG . Ethanol effects on N-methyl-D-aspartate receptors in the bed nucleus of the stria terminalis. Cold Spring Harb Perspect Med 2013; 3: a012161.
Burkett JP, Young LJ . The behavioral, anatomical and pharmacological parallels between social attachment, love and addiction. Psychopharmacology (Berl) 2012; 224: 1–26.
Hagiwara H, Funabashi T, Akema T, Kimura F . Sex-specific differences in pain response by dopamine in the bed nucleus of the stria terminalis in rats. Neuroreport 2013; 24: 181–185.
Kudo T, Uchigashima M, Miyazaki T, Konno K, Yamasaki M, Yanagawa Y et al. Three types of neurochemical projection from the bed nucleus of the stria terminalis to the ventral tegmental area in adult mice. J Neurosci 2012; 32: 18035–18046.
Klampfl SM, Brunton PJ, Bayerl DS, Bosch OJ . Hypoactivation of CRF receptors, predominantly type 2, in the medial-posterior BNST is vital for adequate maternal behavior in lactating rats. J Neurosci 2014; 34: 9665–9676.
Breslau N . The epidemiology of posttraumatic stress disorder: What is the extent of the problem? J Clin Psychiatry 2001; 62: 16–22.
Wakerley JB, Terenzi MG, Housham SJ, Jiang QB, Ingram CD . Electrophysiological effects of oxytocin within the bed nuclei of the stria terminalis: influence of reproductive stage and ovarian steroids. Prog Brain Res 1998; 119: 321–334.
Avery SN, Clauss JA, Winder DG, Woodward N, Heckers S, Blackford JU . BNST neurocircuitry in humans. Neuroimage 2014; 91: 311–323.
Miller MA, Vician L, Clifton DK, Dorsa DM . Sex differences in vasopressin neurons in the bed nucleus of the stria terminalis by in situ hybridization. Peptides 1989; 10: 615–619.
De Vries GJ, al-Shamma HA . Sex differences in hormonal responses of vasopressin pathways in the rat brain. J Neurobiol 1990; 21: 686–693.
Rosie R, Wilson H, Fink G . Testosterone induces an all-or-none, exponential increase in arginine vasopressin mRNA in the bed nucleus of stria terminalis of the hypogonadal mouse. Mol Cell Neurosci 1993; 4: 121–126.
Viau V, Soriano L, Dallman MF . Androgens alter corticotropin releasing hormone and arginine vasopressin mRNA within forebrain sites known to regulate activity in the hypothalamic-pituitary-adrenal axis. J Neuroendocrinol 2001; 13: 442–452.
Fukushima A, Furuta M, Kimura F, Akema T, Funabashi T . Testosterone exposure during the critical period decreases corticotropin-releasing hormone-immunoreactive neurons in the bed nucleus of the stria terminalis of female rats. Neurosci Lett 2013; 534: 64–68.
Cooke BM, Simerly RB . Ontogeny of bidirectional connections between the medial nucleus of the amygdala and the principal bed nucleus of the stria terminalis in the rat. J Comp Neurol 2005; 489: 42–58.
Wang X . Cre transgenic mouse lines. Methods Mol Biol 2009; 561: 265–273.
McGovern AE, Davis-Poynter N, Rakoczy J, Phipps S, Simmons DG, Mazzone SB . Anterograde neuronal circuit tracing using a genetically modified herpes simplex virus expressing EGFP. J Neurosci Methods 2012; 209: 158–167.
Wall NR, Wickersham IR, Cetin A, De La Parra M, Callaway EM . Monosynaptic circuit tracing in vivo through Cre-dependent targeting and complementation of modified rabies virus. Proc Natl Acad Sci USA 2010; 107: 21848–21853.
Gungor NZ, Yamamoto R, Pare D . Optogenetic study of the projections from the bed nucleus of the stria terminalis to the central amygdala. J Neurophysiol 2015; 114: 2903–2911.
van Dijk A, Klanker M, van Oorschot N, Post R, Hamelink R, Feenstra MGP et al. Deep brain stimulation affects conditioned and unconditioned anxiety in different brain areas. Transl Psychiatry 2013; 3: e289.
Islam L, Franzini A, Messina G, Scarone S, Gambini O . Deep brain stimulation of the nucleus accumbens and bed nucleus of stria terminalis for obsessive-compulsive disorder: a case series. World Neurosurg 2015; 83: 657–663.
Acknowledgements
This work was supported by European Research Council FP7 Grant 260463, Israel Science Foundation Research Grants 803/11 and 1565/15, Roberto and Renata Ruhman, Nella and Leon Benoziyo Center for Neurological Diseases, the Henry Chanoch Krenter Institute for Biomedical Imaging and Genomics, the Perlman Family Foundation (founded by Louis L. and Anita M. Perlman), the Adelis Foundation, the Irving I. Moskowitz Foundation, I-CORE Program of the Planning and Budgeting Committee, and Israel Science Foundation Grant 1916/12. Alon Chen is a director of the Max Planck Institute for Psychiatry and head of the Max Planck - Weizmann Laboratory for Experimental Neuropsychiatry and Behavioral Neurogenetics. We thank Dr. Jessica Keverne for English editing, formatting, and scientific input. We would like to give a special thanks to all those who read and re-read versions of the manuscript through the years and commented and brainstormed on each version: Dr Yair Shemesh, Dr Orit Furman and Dr Yair Ben-Efraim. A special thanks also to Sergey Anpilov for the generation of the heatmap of BNST subregions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/4.0/
About this article

Cite this article
Lebow, M., Chen, A. Overshadowed by the amygdala: the bed nucleus of the stria terminalis emerges as key to psychiatric disorders. Mol Psychiatry 21, 450–463 (2016). https://doi.org/10.1038/mp.2016.1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/mp.2016.1
This article is cited by
-
Distinct µ-opioid ensembles trigger positive and negative fentanyl reinforcement
Nature (2024)
-
Threat Responses in Schizophrenia: A Negative Valence Systems Framework
Current Psychiatry Reports (2024)
-
CaMKIIa+ neurons in the bed nucleus of the stria terminalis modulate pace of natural reward seeking depending on internal state
Psychopharmacology (2024)
-
Butterflies in the gut: the interplay between intestinal microbiota and stress
Journal of Biomedical Science (2023)
-
Parasubthalamic calretinin neurons modulate wakefulness associated with exploration in male mice
Nature Communications (2023)
