
Abstract
Bacteria and their viral pathogens face constant pressure for augmented immune and infective capabilities, respectively. Under this reciprocally imposed selective regime, we expect to see a runaway evolutionary arms race, ultimately leading to the extinction of one species. Despite this prediction, in many systems host and pathogen coexist with minimal coevolution even when well-mixed. Previous work explained this puzzling phenomenon by invoking fitness tradeoffs, which can diminish an arms race dynamic. Here we propose that the regular loss of immunity by the bacterial host can also produce host-phage coexistence. We pair a general model of immunity with an experimental and theoretical case study of the CRISPR-Cas immune system to contrast the behavior of tradeoff and loss mechanisms in well-mixed systems. We find that, while both mechanisms can produce stable coexistence, only immune loss does so robustly within realistic parameter ranges.
Similar content being viewed by others

Introduction
While the abundance of bacteria observed globally is impressive (Whitman et al., 1998; Hug et al., 2016; Schloss et al., 2016), any apparent microbial dominance is rivaled by the ubiquity, diversity, and abundance of predatory bacteriophages (or ‘phages’), which target these microbes (Wilhelm and Suttle, 1999; Wommack and Colwell, 2000; Suttle, 2005; Weitz and Wilhelm, 2012; Wigington et al., 2016). As one might expect, phages are powerful modulators of microbial population and evolutionary dynamics, and of the global nutrient cycles these microbes control (Sieburth et al., 1988; Bergh et al., 1989; Bratbak et al., 1990, 1994; Proctor and Fuhrman, 1990; Whitman et al., 1998; Wilhelm and Suttle, 1999; Weinbauer and Rassoulzadegan, 2004; Suttle, 2005; Weitz and Wilhelm, 2012). Despite this ecological importance, we still lack a comprehensive understanding of the dynamical behavior of phage populations. More specifically, it is an open question what processes sustain phages in the long term across habitats.
Bacteria can evade phages using both passive forms of resistance (for example, receptor loss, modification, and masking) and active immune systems that degrade phages (for example, restriction-modification systems, CRISPR-Cas) (Labrie et al., 2010). These defenses can incite an escalating arms race dynamic in which host and pathogen each drive the evolution of the other (Rodin and Ratner, 1983a, 1983b). However, basic theory predicts that such an unrestricted arms race will generally be unstable and sensitive to initial conditions (Schrag and Mittler, 1996). Additionally, if phages have limited access to novel escape mutations, an arms race cannot continue indefinitely (Lenski, 1984; Lenski and Levin, 1985; Hall et al., 2011). This leads to an expectation that phage populations will go extinct in the face of host defenses (Lenski and Levin, 1985).
While typically this expectation holds (for example, van Houte et al., 2016), phages sometimes coexist with their hosts, both in natural (for example, Waterbury and Valois, 1993; Gómez and Buckling, 2011) and laboratory settings (for example, Horne, 1970; Chao et al., 1977; Levin and Udovic, 1977; Lenski and Levin, 1985; Schrag and Mittler, 1996; Bohannan and Lenski, 1999; Wei et al., 2011; Paez-Espino et al., 2015). These examples motivate a search for mechanisms to explain the deescalation and eventual cessation of a coevolutionary arms race dynamic, even in the absence of any spatial structure to the environment. Previous authors have identified (1) fluctuating selection and (2) costs of defense as potential drivers of coexistence in well-mixed systems. Here we propose (3) the loss of immunity, wherein the host defense mechanism ceases to function, as an additional mechanism. We focus on intracellular immunity (for example, CRISPR-Cas) in which immune host act as a sink for phages rather than extracellular resistance (for example, receptor modifications), since the former poses more of an obstacle for phages and thus more of a puzzle for explaining long-term coexistence.
Under a fluctuating selection dynamic, frequencies of immune and infective alleles in the respective host and phage populations cycle over time (Van Valen, 1973, 1974; Agrawal and Lively, 2002; Gandon et al., 2008). That is, old, rare genotypes periodically reemerge because the dominant host or pathogen genotype faces negative frequency dependent selection. Fluctuating selection is likely in situations where host immune and phage infectivity phenotypes match up in a one-to-one “lock and key” type manner (Agrawal and Lively, 2002), and there is evidence that arms races do give way to fluctuating selection in some host-phage systems (Hall et al., 2011). Fluctuating selection cannot always proceed, though. When novel phenotypes correspond to increased generalism we do not expect past phenotypes to recur (Agrawal and Lively, 2002; Gandon et al., 2008) since they will no longer be adaptive. Such expanding generalism during coevolution has been seen in other host-phage systems (Buckling and Rainey, 2002). Thus the relevance of fluctuating selection depends on the nature of the host-phage immune-infective phenotype interaction.
Another possible driver of coexistence are costs incurred by tradeoffs between growth and immunity (for host) or host range and immune evasion (for phage) (Chao et al., 1977; Levin et al., 1977; Jover et al., 2013; Meyer et al., 2016). A tradeoff between immunity and growth rate in the host can lead to the maintenance of a susceptible host population on which phages can persist (Chao et al., 1977; Levin and Udovic, 1977; Lenski and Levin, 1985; Jones and Ellner, 2007; Yoshida et al., 2007; Jover et al., 2013). Tradeoffs often imply a high cost of immunity that does not always exist (for example, Schrag and Mittler, 1996), particularly in the case of intracellular host immunity, as we show later.
Finally, in large host populations typical of bacteria, even low rates of immune loss could produce a substantial susceptible host subpopulation, which, in turn, could support phage reproduction and coexistence. Such loss of function in the host defenses could be due to either mutation or stochastic phenotypic changes. Delbrück (1946) initially described this hypothesis of loss of defense via back-mutation in order to challenge the evidence for lysogeny. Lenski (1988) reiterated this hypothesis in terms of phenotypic plasticity and noted that conditioning the production of a susceptible host population on a resistant one could lead to very robust, host-dominated coexistence. More recently, Meyer et al. (2012) presented an empirical example of a system in which stochastic phenotypic loss of resistance leads to persistence of a coevolving phage population.
We hypothesize that coexistence equilibria will be more robust under an immune loss mechanism than under a tradeoff mechanism (Lenski, 1988). We build a general mathematical model to demonstrate this point and then use a combination of experimental evidence and simulation-based modeling to apply this result to the coevolution of Streptococcus thermophilus and its lytic phage 2972 in the context of CRISPR immunity.
General Immune Loss Model
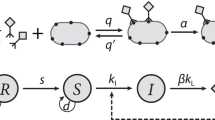
We begin with a general model that considers two populations of host (‘defended’ with a functional immune system; ‘undefended’ without) and one population of pathogen. Starting from classical models of bacteria-phage dynamics (Levin et al., 1977; Weitz, 2016), we add key terms to capture the effects autoimmunity (that is, a tradeoff), immune loss, and the implicit effects of coevolution. This relatively simple model allows us to analyze steady states and parameter interactions analytically. Later, we examine the CRISPR-Cas immune system in detail and build a model with explicit coevolutionary dynamics.
We examine the chemostat system with resources:

defended host:

undefended host:

and phage:

where parameter definitions and values can be found in Table 1 and rationale/references for parameter values in Supplementary Text S2. However, we describe here the parameters of direct relevance to coexistence.
First, we allow for defended host to come with the tradeoff of autoimmunity (α), which applies naturally to the CRISPR-Cas system examined later. While autoimmunity could either decrease the host growth rate (Vercoe et al., 2013) or be lethal, we focus on the latter as lethality will increase the stabilizing effect of this tradeoff (Dy et al., 2013; Paez-Espino et al., 2013; Vercoe et al., 2013). However, we also find similar general results when applying a penalty to the resource affinity or maximum growth rate of the defended host (Supplementary Text S1, Supplementary Figures 1-8). Second, we add flow from the defended to undefended host populations representing loss of immunity at rate μ.
Finally, we model the effect of coevolution by allowing a fraction of even the defended host population to remain susceptible ( ). In a symmetric fashion, even nominally undefended host may have secondary defenses against phage (
). In a symmetric fashion, even nominally undefended host may have secondary defenses against phage ( ).
).
We analyze our model analytically as well as numerically to verify which equilibria are reachable from plausible (for example, experimental) starting values (Supplementary Text S3).
Assuming no phage coevolution ( ), this model has a single analytic equilibrium in which all populations coexist (Supplementary Table 1). In Figure 1, we explore model behavior under varying rates of autoimmunity (α) and immune loss (μ). Clearly when autoimmunity and loss rates surpass unity, defended host go extinct in the face of excessive immune loss and autoimmune targeting. At the opposite parameter extreme, we see coexistence disappear from the numeric solutions (Figure 1b) as phage populations collapse. This leads to a band of parameter space where coexistence is possible, stable, and robust. In this band, autoimmunity and/or immune loss occur at high enough rates to ensure maintenance of coexistence, but not so high as to place an excessive cost on immunity. Crucially, this band is much more constrained in the α-dimension, with autoimmunity restricted to an implausibly high and narrow region of parameter space. This suggests a greater robustness of coexistence under an immune loss mechanism even at low loss rates (Figure 1, Supplementary Figures 2-8). To assess more directly the degree of robustness of each driver of coexistence we can perturb our system and see its response. We move our system away from equilibrium
), this model has a single analytic equilibrium in which all populations coexist (Supplementary Table 1). In Figure 1, we explore model behavior under varying rates of autoimmunity (α) and immune loss (μ). Clearly when autoimmunity and loss rates surpass unity, defended host go extinct in the face of excessive immune loss and autoimmune targeting. At the opposite parameter extreme, we see coexistence disappear from the numeric solutions (Figure 1b) as phage populations collapse. This leads to a band of parameter space where coexistence is possible, stable, and robust. In this band, autoimmunity and/or immune loss occur at high enough rates to ensure maintenance of coexistence, but not so high as to place an excessive cost on immunity. Crucially, this band is much more constrained in the α-dimension, with autoimmunity restricted to an implausibly high and narrow region of parameter space. This suggests a greater robustness of coexistence under an immune loss mechanism even at low loss rates (Figure 1, Supplementary Figures 2-8). To assess more directly the degree of robustness of each driver of coexistence we can perturb our system and see its response. We move our system away from equilibrium  so that
so that  where Y∼Uniform[0,1], and then solve numerically using
where Y∼Uniform[0,1], and then solve numerically using  as our initial condition. Under increasing levels of perturbation the system is less likely to reach stable coexistence, specifically in the α-dimension, indicating that autoimmunity produces a far less robust coexistence regime (Figures 1c and e, Supplementary Figures 2-8).
as our initial condition. Under increasing levels of perturbation the system is less likely to reach stable coexistence, specifically in the α-dimension, indicating that autoimmunity produces a far less robust coexistence regime (Figures 1c and e, Supplementary Figures 2-8).

Model behavior under variations in the rates of autoimmunity (α) and CRISPR-Cas system loss (μ) Equilibria (Supplementary Table 1) derived from Equations 1–4 are shown in (a) where orange indicates a stable equilibrium with all populations coexisting and defended host dominating phage populations, green indicates that all populations coexist but phages dominate, and blue indicates that defended bacteria have gone extinct but phages and undefended bacteria coexist. In (b) we find numerical solutions to the model at 80 days using realistic initial conditions more specific to the experimental setup (R(0)=350, D(0)=106, U(0)=100, P(0)=106). In this case orange indicates coexistence at 80 days with defended host at higher density than phages, green indicates a phage-dominated coexistence at 80 days, and blue indicates that coexistence did not occur. Numerical error is apparent as noise near the orange-blue boundary. We neglect coevolution and innate immunity in this analysis ( ). (c-e) Phase diagrams with perturbed starting conditions. Numerical simulations with starting conditions (X(0)=[R(0),D(0),U(0),P(0)]) were perturbed by a proportion of the equilibrium condition
). (c-e) Phase diagrams with perturbed starting conditions. Numerical simulations with starting conditions (X(0)=[R(0),D(0),U(0),P(0)]) were perturbed by a proportion of the equilibrium condition  where Y∼U[0, 1] and
where Y∼U[0, 1] and  signifies an equilibrium value to explore how robust the equilibria are to starting conditions. A single simulation was run for each parameter combination.
signifies an equilibrium value to explore how robust the equilibria are to starting conditions. A single simulation was run for each parameter combination.
If we add large amounts of innate immunity to undefended host ( ), we find phage-dominated coexistence for a wider range of α (Supplementary Figure 10). This result is in line with the counterintuitive suggestion that higher immunity may increase phage density by allowing the host population to increase in size (Iranzo et al., 2013). However, secondary defense has minimal effects for more plausible levels of protection (
), we find phage-dominated coexistence for a wider range of α (Supplementary Figure 10). This result is in line with the counterintuitive suggestion that higher immunity may increase phage density by allowing the host population to increase in size (Iranzo et al., 2013). However, secondary defense has minimal effects for more plausible levels of protection ( closer to 1).
closer to 1).
In the case of phage coevolution ( ), the equilibria still have closed forms, but are not easily representable as simple equations and so are not written here. When
), the equilibria still have closed forms, but are not easily representable as simple equations and so are not written here. When  , defended host contribute positively to phage growth, eventually shifting the coexistence equilibrium from host to phage dominance (Supplementary Figure 9).
, defended host contribute positively to phage growth, eventually shifting the coexistence equilibrium from host to phage dominance (Supplementary Figure 9).
A Case Study: CRISPR-Phage Coevolution
The CRISPR (Clustered Regularly Inter-spaced Short Palindromic Repeats) prokaryotic adaptive immune system incorporates specific immune memory in the form of short sequences of DNA acquired from foreign genetic elements (‘spacers’) and then uses this memory to target the corresponding sequences (‘protospacers’) during subsequent infections (Bolotin et al., 2005; Mojica et al., 2005; Barrangou et al., 2007; Garneau et al., 2010). CRISPR can lead to rapidly escalating arms races between bacteria and phages (Deveau et al., 2008; Horvath et al., 2008; Paez-Espino et al., 2015), in which evolutionary and population dynamics occur on the same timescale (Gerrish and Lenski, 1998; Desai et al., 2007; Childs et al., 2012; Paez-Espino et al., 2015).
CRISPR-Cas can quickly drive phages extinct in an experimental setting (van Houte et al., 2016), but in some cases long-term CRISPR-phage coexistence has been observed (Paez-Espino et al., 2015). Previous theoretical and limited experimental work has explained short-term coexistence through tradeoffs and spacer loss (Bradde et al., 2017), and long-term coexistence by invoking continued coevolution via fluctuating selection (Childs et al., 2012) or tradeoffs with host switching to a constitutive defense strategy such as surface receptor modification (Westra et al., 2015; Chabas et al., 2016).
However, these previous hypotheses are insufficient to explain simple coevolution experiments with Streptococcus thermophilus (type II-A CRISPR-Cas system) and its lytic phage 2972 resulting in long-term coexistence (Paez-Espino et al., 2013, 2015). In these experiments, bacteria are resource-limited and appear immune to phages, implying they have ‘won’ the arms race and that phages are persisting on a small susceptible subpopulation of hosts. Deep sequencing of the same experimental system shows dominance by a few spacers that drift in frequency over time, inconsistent with a fluctuating selection dynamic (Paez-Espino et al., 2013). Specifically, these results contradict the coexistence regime seen in the (Childs et al. 2012, 2014) model, wherein host are phage-limited and the system undergoes a fluctuating selection dynamic. Thus either (1) costs associated with CRISPR immunity or (2) the loss of CRISPR immunity is playing a role in maintaining susceptible host subpopulations on which phages can persist.
In this system, the primary cost of a functional CRISPR-Cas system is autoimmunity via the acquisition of self-targeting spacers. It is unclear how or if bacteria distinguish self from non-self during the acquisition step of CRISPR immunity (Stern et al., 2010; Yosef et al., 2012; Kumar et al., 2015; Levy et al., 2015; Wei et al., 2015). In S. thermophilus, experimental evidence suggests that there is no mechanism of self vs. non-self recognition and that self-targeting spacers are acquired frequently (Wei et al., 2015), which implies that autoimmunity may be a significant cost.
Outright loss of CRISPR immunity at a high rate could also lead to coexistence. The bacterium Staphylococcus epidermidis loses phenotypic functionality in its CRISPR-Cas system, either due to wholesale deletion of the relevant loci or mutation of essential sequences (i.e. the leader sequence or cas genes), at a rate of 10–4-10−3 inactivation/loss events per individual per generation (Jiang et al., 2013). Functional CRISPR loss has been observed in other systems as well (Palmer and Gilmore, 2010; Garrett et al., 2011).
Below we replicate the serial-transfer coevolution experiments performed by (Paez-Espino et al. 2013, 2015) and develop a simulation-based coevolutionary model to explain the phenomenon of coexistence.
Experiments
We performed long-term daily serial transfer experiments with S. thermophilus and its lytic phage 2972 in milk, a model system for studying CRISPR evolution (see Supplementary Text S4 for detailed methods). We measured bacteria and phage densities on a daily basis. Further, on selected days we PCR-amplified and sequenced the CRISPR1 and CRISPR3 loci, the two adaptive CRISPR loci in this bacterial strain.
From the perspective of density, phages transiently dominated the system early on, but the bacteria quickly took over and by day five appeared to be resource-limited rather than phage-limited (Figures 2a and b). This switch to host-dominance corresponded to a drop in phage populations to a titer two to three orders of magnitude below that of the bacteria. Once arriving at this host-dominated state, the system either maintained quasi-stable coexistence on an extended timescale (over a month and a half), or phages continued to decline and went extinct relatively quickly (Figures 2a and b). We performed six additional replicate experiments which confirmed this dichotomy between either extended coexistence (4 lines quasi-stable for >2 weeks) or quick phage extinction (2 lines <1 week) (Supplementary Figure 11).

Serial transfer experiments carried out with S. thermophilus and lytic phage 2972 Bacteria are resource-limited rather than phage-limited by day five and phages can either (a) persist at relatively low density in the system on long timescales (greater than 1 month) or (b) collapse relatively quickly. These results agree with those of (Paez-Espino et al., 2015) where coexistence was observed in S. thermophilus and phage 2972 serially transferred culture for as long as a year. Experiments were initiated with identical starting populations and carried out following the same procedure. In (c-e) we show that our simulations replicate the qualitative patterns seen in the data, with an early phage peak, followed by host-dominated coexistence that can either be (c) stable, (d) sustained but unstable, or (e) short-lived. Each plot is a single representative simulation and simulations were ended when phages went extinct. Note that experimental data has a resolution of one time point per day, preventing conclusions about the underlying population dynamics (for example, cycling), whereas simulations are continuous in time.
Sequencing of the CRISPR1 and CRISPR3 loci revealed the rapid gain of a single spacer (albeit different spacers in different sequenced clones) in CRISPR1 followed by minor variation in spacer counts with time (Supplementary Figure 12), with CRISPR1 being more active than CRISPR3. We tracked the identity of the first novel spacer in the CRISPR1 array over time. We found a cohort of four spacers that persisted over time and were repeatedly seen despite a small number of samples taken at each time point (less than 10 per time point; Table 2). Other spacers were sampled as well, but this small cohort consistently reappeared while other spacers were only found at one or two timepoints, indicating this cohort was dominating the system (Supplementary Table 2). Such a pattern is inconsistent with a fluctuating selection hypothesis. Further, we did not observe frequent spacer loss in the CRISPR1 or CRISPR3 arrays.
CRISPR-phage Coevolutionary Model
We next built a hybrid deterministic/stochastic lineage-based model similar to an earlier model by (Childs et al. 2012, 2014) that explicitly models the coevolutionary dynamics of the CRISPR-phage system wherein bacteria acquire spacers to gain immunity and phages escape spacers via mutations. Our simulations also replicate the resource dynamics of a serial dilution experiment, wherein the system undergoes large daily perturbations.
We model phage mutations only in the protospacer adjacent motif (PAM) region, which is the dominant location of CRISPR escape mutations (Paez-Espino et al., 2015) to prevent the possibility of spacer re-acquisition. This approach differs from previous models which considered mutations in the protospacer region itself (for example, Childs et al., 2012; Weinberger et al., 2012; Iranzo et al., 2013) and thus allowed for the possibility of spacer re-acquisition. We justify modeling only PAM mutations with three arguments. First, the probability of spacer re-acquisition will be quite low if there are many protospacers. Second, re-acquired spacers will already have undergone selection for escape mutation by phage, and, assuming that there are therefore diverse escape mutations in the phage population, these spacers will thus provide limited benefit to the host. Third, as we move away from the PAM along the protospacer sequence, more substitutions are tolerated by the CRISPR matching machinery (Semenova et al., 2011), meaning that mutations farther away from the PAM will be less effective at escaping immunity (Martel and Moineau, 2014).
We model population dynamics using differential equations for resources:

CRISPR-enabled bacteria with spacer set Xi:

a pool of undefended bacteria with a missing or defective CRISPR-Cas system:

and phages with protospacer set Yi:

and stochastic events occur according to a Poisson process with rate λ:

which is a sum of the total per-strain spacer-acquisition rates:

total per-strain PAM mutation rates:

and total per-strain PAM back mutation rates:

In this way each unique CRISPR genotype (Xi), defined as a set of linked spacers sharing the same array, is modeled individually, as is each phage genotype (Yi). As new spacers are added and new PAMs undergo mutation, new pairs of genotypes and equations are added to the system. Host that have undergone immune loss are modeled separately (U), as if they have no CRISPR-Cas system.
The function M(Xi,Yj) is a binary matching function between (proto)spacer content of bacterial and phage genomes that determines the presence or absence of immunity. We refer to the ‘order’ of a host or phage strain, which is the number of evolutionary events that strain has undergone, |Xi| or ns-|Yi| respectively. The PAM back mutation rate μq describes the rate at which we expect a mutated PAM to revert to its original sequence (assuming the mutation is a substitution). While back mutation is not required to generate stable host-dominated coexistence, it greatly expands the relevant region of parameter space because it allows phages to avoid the cost we will impose on PAM mutations, discussed below, when those immune escape mutations are no longer beneficial. Recombination among viral strains could have a similar effect by providing another route to an un-mutated or less mutated genome. Paez-Espino et al. (2015) suggest that recombination can produce stable host-dominated coexistence, although we reject such diversity-driven hypotheses (for example, Childs et al., 2012) based on our sequencing data.
We assume that the number of PAM mutations in a single phage genome is constrained by a tradeoff with phage fitness, as this is necessary to prevent the total clearance of protospacers from a single strain at high mutation rates. Increases in host breadth at the species level generally come at a cost for viruses due to pleiotropic effects (Ferris et al., 2007). More broadly, mutations tend to be deleterious on average (for example, Chao, 1990). It is reasonable to speculate that phages have evolved under pressure to lose any active PAMs on their genomes, and thus that the persisting PAMs may have been preserved because their loss is associated with a fitness cost.
The function

incorporates a linear cost of mutation into the phage burst size. See Table 3 for further definitions of variables, functions, and parameters in Equations 5,6,7,8,9,10,11,12,13. Simulation procedures and rationale for parameter values, including phage genome size, are detailed in Supplementary Text S3.
Stable Host-Dominated coexistence
Simulations with immune loss reliably produce extended coexistence within a realistic region of the parameter space (Figure 3) thus replicating our experimental results (Figure 2), and confirming our qualitative results from the simpler deterministic model (Figure 1). We observed no simulations in which autoimmunity alone produced stable coexistence. This agrees with our earlier numerical results from the general model where unrealistically high rates of autoimmunity were required to produce coexistence.

Distribution of phage extinction times in bacterial-dominated cultures with different possible combinations of coexistence mechanisms The peak at >75 corresponds to what we call stable coexistence (simulations ran for a maximum of 80 days). There is no significant difference between the top two panels in the number of simulations reaching the 80 day mark (x2=2.8904, df=1, P-value=0.08911). Back mutation was set at μq=5 × 10−9.
Similar to our experimental results, for a single set of parameters this model can stochastically fall into either stable coexistence or a phage-free state (Figure 3). The relative frequencies with which we see each outcome, as well as the distribution of times that phages are able to persist, depend on the specific set of parameters chosen. In particular, increasing the PAM back mutation rate will increase the probability of the coexistence outcome (Figure 4), although even in the absence of back mutation the system will occasionally achieve stable coexistence. This dependence on back mutation is caused by the combined effects of the cumulative cost we impose on PAM mutations and the inability of phages to keep up with host in a continuing arms race. In the early stages of the arms race it is optimal for phages to continue undergoing PAM mutations as the most abundant available hosts are high-order CRISPR variants, whereas once hosts are able to pull sufficiently ahead of phages in the arms race it becomes optimal for phages to feed on the lower-density but consistently available CRISPR-lacking host population (Supplementary Figure 13).

Distribution of phage extinction times in bacterial-dominated cultures with different rates of PAM back mutation in phages (μq) The peak at >70 corresponds to what we call stable coexistence (simulations ran for a maximum of 80 days). These results are shown for a locus-loss mechanism only (μL=5 × 10−4, α=0). The histogram for μq=5 × 10-8 is omitted as it is nearly identical to that for μq=5 × 10−9, indicating that the height of the coexistence peak saturates at high back mutation.
The adsorption rate, on a coarse scale, has an important effect on how the model behaves (Supplementary Figure 14). At high values of δ where we would expect phages to cause host extinction in the absence of CRISPR immunity (δ=10−7) we see that long-term coexistence occurs rarely, and is negatively associated with the phage back mutation rate. In this case phages will rapidly consume the susceptible host population and crash to extinction unless they have undergone PAM mutations that lower their growth rate. This causes a reversal in the previous trend seen with back mutation where the ability of phages to escape the costs of PAM mutation was essential to their persistence. A decrease in the adsorption rate to a very low value (δ=10−9) leads to most simulations persisting in host-dominated coexistence until the 80 day cutoff. Because both evolutionary and demographic dynamics occur much more slowly in this case, long term persistence does not necessarily imply actual stability, as suggested by our and previous (Paez-Espino et al., 2015) experimental results in which coexistence eventually ends. In general, lower adsorption rates lead to longer periods of host-dominated coexistence and reduce the chance of phage extinction.
The failure of autoimmunity to produce coexistence warrants further investigation. Upon closer examination, it is clear that in the early stages of the arms race where CRISPR-enabled bacteria have not yet obtained spacers or been selected for in the host population, phages are able to proliferate to extremely high levels and greatly suppress the CRISPR-lacking host. Because autoimmunity as a mechanism of coexistence relies on the continued presence of immune-lacking host, it may not be able to function in the face of this early phage burst if susceptible host are driven extinct. There is a possibility that very low locus loss rates that reintroduce CRISPR-lacking bacteria but do not appreciably contribute to their density combined with high rates of autoimmunity could maintain high enough density susceptible host populations to sustain phage. To investigate this possibility we imposed a floor of U>1 and ran further simulations. Even with very high rates of autoimmunity based on an upper limit of likely spacer acquisition rates (α=50μb, μb=10−5) the susceptible host population does not grow quickly enough to sufficiently high levels to sustain phage (Supplementary Figure 15). Thus it is not early dynamics that rule out autoimmunity but the insufficiency of the mechanism itself for maintaining large enough susceptible host populations.
Transient coexistence with low density phage
While we do not observe stable coexistence in any case where there is not loss of the CRISPR-Cas immune system, we did observe prolonged phage persistence in some cases where μL=α=0 (Figure 3) and in cases with autoimmunity only (μL=0). Phages were able to persist at very low density (:10-100 particles/mL) for as long as two months in a host-dominated setting without the presence of a CRISPR-lacking host subpopulation (Figure 3, Supplementary Figure 16). It appears that in these cases phages are at sufficiently low density as to have a minimal effect on their host population and thus that host strain is selected against very slowly. Because the phages have undergone many PAM mutations at this point they are unable to proliferate rapidly enough between dilution events to have an easily measurable impact on the host population. Essentially, phages delay their collapse by consuming their host extremely slowly (Supplementary Figure 16). However, with an active locus loss mechanism (that is, μL>0), we did not see this sustained but unstable coexistence occur, likely because the undefended hosts would have driven the phage population to higher levels and increased selection on the susceptible CRISPR variants.
Discussion
We paired a general model of immunity with a case study of the CRISPR immune system to characterize and contrast the potential drivers of long-term host-phage coexistence in well-mixed systems. We found that a tradeoff mechanism does not lead to a robust coexistence equilibirum in the case of intracellular host immunity. We also ruled out coevolutionary drivers of coexistence in the S. thermophilus-phage 2972 system based on a combination of our own sequencing data and previous work on the same system (Paez-Espino et al., 2013). Since some mechanism(s) must be producing susceptible hosts on which phages can replicate, we are left with an immune loss hypothesis as the best remaining explanation for our empirical results. Our simulations showed that the addition of early coevolutionary dynamics alongside immune loss replicates key features of our experimental results, including stochastic switching between the possible outcomes of long term coexistence and rapid phage clearance. Therefore we predict that that the CRISPR-Cas immune system is lost at a nontrivial rate in S. thermophilus in addition to S. epidermidis (Jiang et al., 2013), and possibly other species.
With regards to CRISPR, while our experiments do not speak to the relative importance of locus loss versus costly autoimmunity, our theoretical results reject autoimmunity as a realistic mechanism of phage persistence. Our experimental setup was in serial dilution, which subjects the culture to large daily perturbations, ruling out any mechanism that does not produce a robust coexistence regime.
We emphasize that CRISPR immunity, and immunity in general, is still likely costly (Vale et al., 2015).Nevertheless, in cases of intracellular host immunity those costs are insufficient to drive continued phage persistence in the environment. Intracellular immunity destroys phages rather than simply preventing phage replication. Thus the threshold density of susceptible host for phage persistence is higher than in systems where hosts have an extracellular defense strategy (i.e. receptor/envelope modification), meaning the cost of immunity must be higher. When hosts escape phage predation via receptor modifications, a growth-resistance tradeoff may lead to coexistence.
Our sequencing results in the S. thermophilus system reject coevolutionary mechanisms for coexistence. We can directly reject an arms race dynamic since it predicts the rapid, continued accumulation of spacers, which does not occur in our data. A fluctuating selection dynamic makes the more subtle prediction that the frequencies of spacers in the population should cycle over time, with different spacers dominating at different times. Even with relatively small sample sizes (<10 CRISPR loci sequenced per timepoint), we see a small cohort of spacers increase in frequency early in the experiment and continue to be detected at later timepoints (Table 2). These results are consistent with those of Paez-Espino et al. (2013) who performed deep sequencing with the same phage-host system and observed dominant spacers that drifted in frequency over time. This continued detection and dominance of particular spacers rules out strong fitness differences between these spacers, which, in turn, contradicts the expectation of fluctuating selection that fitnesses change over time. Our stochastic simulations agree, with coevolutionary dynamics in the absence of loss or cost most often yielding rapid phage extinction and only occasionally showing coexistence for over a month - but never exhibiting sustained coexistence (Figure 3).
A similar model by Childs et al. (2014) found that a fluctuating selection dynamic could lead to long term coexistence in a CRISPR-phage system when arrays were ‘saturated”, in the sense that they were filled to some preset maximum capacity with spacers, which we do not observe in our experimental data. The fact that we see little expansion of the array suggests that hosts are completely immune to phages, as rapid phage genome degradation inside the CRISPR-immune cells can prevent further uptake of spacers (Semenova et al., 2016).
While we conclude that immune loss plays a key role in our system, it is not immediately clear why bacterial immune systems would lose functionality at such a high rate. Our sequencing of the S. thermophilus CRISPR loci did not reveal pervasive spacer loss events, indicating that immune loss is at the system rather than spacer level. Perhaps in the case of CRISPR there is some inherent instability of the locus, leading to high rates of horizontal transfer (Godde and Bickerton, 2006; Chakraborty et al., 2010; Palmer and Gilmore, 2010; Garrett et al., 2011; Shah and Garrett, 2011). Jiang et al. (2013) propose that CRISPR loss is a bet-hedging strategy that allows horizontal gene transfer to occur in stressful environments (for example, under selection for antibiotic resistance). This proposal is consistent with evidence that CRISPR does not inhibit horizontal gene transfer on evolutionary timescales (Gophna et al., 2015). A high rate of CRISPR loss and inactivation could produce pressure for bacteria to frequently acquire new CRISPR-Cas systems through horizontal gene transfer, perhaps explaining why strains with multiple CRISPR-Cas systems are frequently observed, including S. thermophilus (Horvath et al., 2009; Cai et al., 2013). This is consistent with a broader view in which prokaryotic defense systems appear to be labile, having higher rates of gain and loss than other genetic content (Puigbò et al., 2017).
While some clear anecdotes of immune loss exist (Meyer et al., 2012; Jiang et al., 2013), other examples of this phenomenon may have been missed because it is difficult to detect. Phages will quickly destroy any evidence of loss, and loss rates can be low while still affecting population dynamics. Jiang et al. (2013) go to great lengths to demonstrate loss in their system. Particularly with complex systems like CRISPR-Cas, a mutation in any number of components can lead to inactivation, making loss hard to detect from genetic screens. Phenotypic screens like those of Jiang et al. (2013) require the engineering of CRISPR spacer content and/or plasmid sequence as well as an otherwise competent host.
Other paths to sustained coexistence between CRISPR-enabled hosts and phages may also exist. There is a great diversity of CRISPR-Cas system types and modes of action (Makarova et al., 2015) and the particular mechanism of each system may lead to distinct host-phage dynamics. That being said, our model of CRISPR evolutionary dynamics is rather general, and we recovered similar qualitative results over a wide range of parameter values apart from the S. thermophilus-specific parameter space.
Finally, our results show that the regular loss of immunity can sustain a viable phage population, leading to the maintenance of selective pressure and thus keeping immunity prevalent in the population overall. Even though long-term coexistence with phages may not affect overall host population density, we suggest that, counterintuitively, the periodic loss of individual immunity may drive the maintenance of a high population immune prevalence.
References
Agrawal A, Lively CM . (2002). Infection genetics: gene-for-gene versus matching-alleles models and all points in between. Evolutionary Ecology Research 4: 91–107.
Barrangou R, Fremaux C, Deveau H, Richards M, Boyaval P, Moineau S et al. (2007). CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science 315: 1709–1712.
Bergh Ø, Børsheim KY, Bratbak G, Heldal M . (1989). High abundance of viruses found in aquatic environments. Nature 340: 467–468.
Bohannan BJM, Lenski RE . (1999). Effect of prey heterogeneity on the response of a model food chain to resource enrichment. The American Naturalist 153: 73–82.
Bolotin A, Quinquis B, Sorokin A, Ehrlich SD . (2005). Clustered regularly interspaced short palindrome repeats (CRISPRs) have spacers of extrachromosomal origin. Microbiology 151: 2551–2561.
Bradde S, Vucelja M, Tesileanu T, Balasubramanian V . (2017). Dynamics of adaptive immunity against phage in bacterial populations. PLoS Computational Biology 13: e1005486.
Bratbak G, Heldal M, Norland S, Thingstad TF . (1990). Viruses as Partners in Spring Bloom Microbial Trophodynamics. Applied and Environmental Microbiology 56: 1400–1405.
Bratbak G, Thingstad F, Heldal M . (1994). Viruses and the Microbial Loop. Microbial Ecology 28: 209–221.
Buckling A, Rainey PB . (2002). Antagonistic coevolution between a bacterium and a bacteriophage. Proceedings of the Royal Society of London B: Biological Sciences 269: 931–936.
Cai F, Axen SD, Kerfeld CA . (2013). Evidence for the widespread distribution of CRISPR-Cas system in the Phylum Cyanobacteria. RNA Biology 10: 687–693.
Chabas H, van Houte S, Høyland-Kroghsbo NM, Buckling A, Westra ER . (2016). Immigration of susceptible hosts triggers the evolution of alternative parasite defence strategies. Proceedings Biological sciences 283: 2016721.
Chakraborty S, Snijders AP, Chakravorty R, Ahmed M, Tarek AM, Hossain MA . (2010). Comparative network clustering of direct repeats (DRs) and cas genes confirms the possibility of the horizontal transfer of CRISPR locus among bacteria. Molecular Phylogenetics and Evolution 56: 878–887.
Chao L, Levin BR, Stewart FM . (1977). A Complex Community in a Simple Habitat: An Experimental Study with Bacteria and Phage. Ecology 58: 369–378.
Chao L . (1990). Fitness of RNA virus decreased by Muller's ratchet. Nature 348: 454–455.
Childs LM, England WE, Young MJ, Weitz JS, Whitaker RJ . (2014). CRISPR-Induced Distributed Immunity in Microbial Populations. PLoS One 9: e101710.
Childs LM, Held NL, Young MJ, Whitaker RJ, Weitz JS . (2012). Multiscale Model of Crispr-Induced Coevolutionary Dynamics: Diversification at the Interface of Lamarck and Darwin. Evolution 66: 2015–2029.
Delbrück M . (1946). Bacterial Viruses or Bacteriophages. Biological Reviews 21: 30–40.
Desai MM, Fisher DS, Murray AW . (2007). The Speed of Evolution and Maintenance of Variation in Asexual Populations. Current Biology 17: 385–394.
Deveau H, Barrangou R, Garneau JE, Labonté J, Fremaux C, Boyaval P et al. (2008). Phage Response to CRISPR-Encoded Resistance in Streptococcus thermophilus. Journal of Bacteriology 190: 1390–1400.
Dy RL, Pitman AR, Fineran PC . (2013). Chromosomal targeting by CRISPR-Cas systems can contribute to genome plasticity in bacteria. Mobile Genetic Elements 3: e26831.
Ferris MT, Joyce P, Burch CL . (2007). High Frequency of Mutations That Expand the Host Range of an RNA Virus. Genetics 176: 1013–1022.
Gandon S, Buckling A, Decaestecker E, Day T . (2008). Host–parasite coevolution and patterns of adaptation across time and space. Journal of Evolutionary Biology 21: 1861–1866.
Garneau JE, Dupuis ME, Villion M, Romero DA, Barrangou R, Boyaval P et al. (2010). The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid dna. Nature 468: 67–71.
Garrett RA, Shah SA, Vestergaard G, Deng L, Gudbergsdottir S, Kenchappa CS et al. (2011). CRISPR-based immune systems of the Sulfolobales: complexity and diversity. Biochemical Society Transactions 39: 51–57.
Gerrish PJ, Lenski RE . (1998). The fate of competing beneficial mutations in an asexual population. Genetica 102-103: 127.
Godde JS, Bickerton A . (2006). The repetitive DNA elements called CRISPRs and their associated genes: evidence of horizontal transfer among prokaryotes. Journal of Molecular Evolution 62: 718–729.
Gómez P, Buckling A . (2011). Bacteria-Phage Antagonistic Coevolution in Soil. Science 332: 106–109.
Gophna U, Kristensen DM, Wolf YI, Popa O, Drevet C, Koonin EV . (2015). No evidence of inhibition of horizontal gene transfer by CRISPR–Cas on evolutionary timescales. The ISME Journal 9: 2021–2027.
Hall AR, Scanlan PD, Morgan AD, Buckling A . (2011). Host–parasite coevolutionary arms races give way to fluctuating selection. Ecology Letters 14: 635–642.
Horne MT . (1970). Coevolution of Escherichia coli and Bacteriophages in Chemostat Culture. Science 168: 992–993.
Horvath P, Coûté-Monvoisin AC, Romero DA, Boyaval P, Fremaux C, Barrangou R . (2009). Comparative analysis of CRISPR loci in lactic acid bacteria genomes. International Journal of Food Microbiology 131: 62–70.
Horvath P, Romero DA, Coûté-Monvoisin AC, Richards M, Deveau H, Moineau S et al. (2008). Diversity, Activity, and Evolution of CRISPR Loci in Streptococcus thermophilus. Journal of Bacteriology 190: 1401–1412.
Hug LA, Baker BJ, Anantharaman K, Brown CT, Probst AJ, Castelle CJ et al. (2016). A new view of the tree of life. Nature Microbiology 1: 16048.
Iranzo J, Lobkovsky AE, Wolf YI, Koonin EV . (2013). Evolutionary Dynamics of the Prokaryotic Adaptive Immunity System CRISPR-Cas in an Explicit Ecological Context. Journal of Bacteriology 195: 3834–3844.
Jiang W, Maniv I, Arain F, Wang Y, Levin BR, Marraffini LA . (2013). Dealing with the Evolutionary Downside of CRISPR Immunity: Bacteria and Beneficial Plasmids. PLoS Genet 9: e1003844.
Jones LE, Ellner SP . (2007). Effects of rapid prey evolution on predator–prey cycles. Journal of Mathematical Biology 55: 541–573.
Jover LF, Cortez MH, Weitz JS . (2013). Mechanisms of multi-strain coexistence in host–phage systems with nested infection networks. Journal of Theoretical Biology 332: 65–77.
Kumar MS, Plotkin JB, Hannenhalli S . (2015). Regulated CRISPR Modules Exploit a Dual Defense Strategy of Restriction and Abortive Infection in a Model of Prokaryote-Phage Coevolution. PLoS Computational Biology 11: e1004603.
Labrie SJ, Samson JE, Moineau S . (2010). Bacteriophage resistance mechanisms. Nature reviews Microbiology 8: 317.
Lenski RE, Levin BR . (1985). Constraints on the Coevolution of Bacteria and Virulent Phage: A Model, Some Experiments, and Predictions for Natural Communities. The American Naturalist 125: 585–602.
Lenski RE . (1984). Coevolution of bacteria and phage: Are there endless cycles of bacterial defenses and phage counterdefenses? Journal of Theoretical Biology 108: 319–325.
Lenski RE . (1988) Dynamics of Interactions between Bacteria and Virulent Bacteriophage. In: Marshall KC, editor Advances in Microbial Ecology, number 10 in Advances in Microbial Ecology pages Springer: US, 1–44.
Levin BR, Stewart FM, Chao L . (1977). Resource-Limited Growth, Competition, and Predation: A Model and Experimental Studies with Bacteria and Bacteriophage. The American Naturalist 111: 3–24.
Levin SA, Udovic JD . (1977). A Mathematical Model of Coevolving Populations. The American Naturalist 111: 657–675.
Levy A, Goren MG, Yosef I, Auster O, Manor M, Amitai G et al. (2015). CRISPR adaptation biases explain preference for acquisition of foreign DNA. Nature 520: 505–510.
Makarova KS, Wolf YI, Alkhnbashi OS, Costa F, Shah SA, Saunders SJ et al. (2015). An updated evolutionary classification of CRISPR-Cas systems. Nature Reviews Microbiology 13: 722–736.
Martel B, Moineau S . (2014). Crispr-cas: an efficient tool for genome engineering of virulent bacteriophages. Nucleic acids research 42: 9504–9513.
Meyer JR, Dobias DT, Medina SJ, Servilio L, Gupta A, Lenski RE . (2016). Ecological speciation of bacteriophage lambda in allopatry and sympatry. Science 354: 1301–1304.
Meyer JR, Dobias DT, Weitz JS, Barrick JE, Quick RT, Lenski RE . (2012). Repeatability and Contingency in the Evolution of a Key Innovation in Phage Lambda. Science 335: 428–432.
Mojica FJM, Díez-Villaseñor C, García-Martínez J, Soria E . (2005). Intervening Sequences of Regularly Spaced Prokaryotic Repeats Derive from Foreign Genetic Elements. Journal of Molecular Evolution 60: 174–182.
Paez-Espino D, Morovic W, Sun CL, Thomas BC, Ueda KI, Stahl B et al. (2013). Strong bias in the bacterial CRISPR elements that confer immunity to phage. Nature Communications 4: 1430.
Paez-Espino D, Sharon I, Morovic W, Stahl B, Thomas BC, Barrangou R et al. (2015). CRISPR immunity drives rapid phage genome evolution in Streptococcus thermophilus. MBio 6: e00262–15.
Palmer KL, Gilmore MS . (2010). Multidrug-Resistant Enterococci Lack CRISPR-cas. mBio 1: e00227–10.
Proctor LM, Fuhrman JA . (1990). Viral mortality of marine bacteria and cyanobacteria. Nature 343: 60–62.
Puigbò P, Makarova KS, Kristensen DM, Wolf YI, Koonin EV . (2017). Reconstruction of the evolution of microbial defense systems. BMC Evolutionary Biology 17: 94.
Rodin SN, Ratner VA . (1983a). Some theoretical aspects of protein coevolution in the ecosystem "phage-bacteria" II. The deterministic model of microevolution. Journal of Theoretical Biology 100: 197–210.
Rodin SN, Ratner VA . (1983b). Some theoretical aspects of protein coevolution in the ecosystem "phage-bacteria" I. The problem. Journal of Theoretical Biology 100: 185–195.
Schloss PD, Girard RA, Martin T, Edwards J, Thrash JC . (2016). Status of the Archaeal and Bacterial Census: an Update. mBio 7: e00201–e00216.
Schrag SJ, Mittler JE . (1996). Host-Parasite Coexistence: The Role of Spatial Refuges in Stabilizing Bacteria-Phage Interactions. The American Naturalist 148: 348–377.
Semenova E, Jore MM, Datsenko KA, Semenova A, Westra ER, Wanner B et al. (2011). Interference by clustered regularly interspaced short palindromic repeat (CRISPR) RNA is governed by a seed sequence. Proceedings of the National Academy of Sciences 108: 10098–10103.
Semenova E, Savitskaya E, Musharova O, Strotskaya A, Vorontsova D, Datsenko KA et al. (2016). Highly efficient primed spacer acquisition from targets destroyed by the Escherichia coli type I-E CRISPR-Cas interfering complex. Proceedings of the National Academy of Sciences 113: 7626–7631.
Shah SA, Garrett RA . (2011). CRISPR/Cas and Cmr modules, mobility and evolution of adaptive immune systems. Research in Microbiology 162: 27–38.
Sieburth JM, Johnson PW, Hargraves PE . (1988). Ultrastructure and Ecology of Aureococcus Anophageferens Gen. Et Sp. Nov. (chrysophyceae): The Dominant Picoplankter During a Bloom in Narragansett Bay, Rhode Island, Summer 19851. Journal of Phycology 24: 416–425.
Stern A, Keren L, Wurtzel O, Amitai G, Sorek R . (2010). Self-targeting by CRISPR: gene regulation or autoimmunity? Trends in Genetics 26: 335–340.
Suttle CA . (2005). Viruses in the sea. Nature 437: 356–361.
Vale PF, Lafforgue G, Gatchitch F, Gardan R, Moineau S, Gandon S . (2015). Costs of CRISPR-Cas-mediated resistance in Streptococcus thermophilus. Proceedings Biological Sciences / The Royal Society 282: 20151270.
van Houte S, Ekroth AKE, Broniewski JM, Chabas H, Ashby B, Bondy-Denomy J et al. (2016). The diversity-generating benefits of a prokaryotic adaptive immune system. Nature 532: 385–388.
Van Valen L . (1973). A new evolutionary law. Evolutionary Theory 1: 1–30.
Van Valen L . (1974). Molecular evolution as predicted by natural selection. Journal of Molecular Evolution 3: 89–101.
Vercoe RB, Chang JT, Dy RL, Taylor C, Gristwood T, Clulow JS et al. (2013). Cytotoxic Chromosomal Targeting by CRISPR/Cas Systems Can Reshape Bacterial Genomes and Expel or Remodel Pathogenicity Islands. PLOS Genet 9: e1003454.
Waterbury JB, Valois FW . (1993). Resistance to Co-Occurring Phages Enables Marine Synechococcus Communities To Coexist with Cyanophages Abundant in Seawater. Applied and Environmental Microbiology 59: 3393–3399.
Weinbauer MG, Rassoulzadegan F . (2004). Are viruses driving microbial diversification and diversity? Environmental microbiology 6: 1–11.
Weinberger AD, Wolf YI, Lobkovsky AE, Gilmore MS, Koonin EV . (2012). Viral Diversity Threshold for Adaptive Immunity in Prokaryotes. mBio 3: e00456–12.
Weitz JS, Wilhelm SW . (2012). Ocean viruses and their effects on microbial communities and biogeochemical cycles. F1000 Biology Reports 4 doi:10.3410/B4-17.
Weitz JS . (2016) Quantitative Viral Ecology: Dynamics of Viruses and Their Microbial Hosts. Princeton University Press, Google-Books-ID: 0zNJCgAAQBAJ.
Wei Y, Kirby A, Levin BR, Rohani AEP, Bronstein EJL . (2011). The Population and Evolutionary Dynamics of Vibrio cholerae and Its Bacteriophage: Conditions for Maintaining Phage-Limited Communities. The American Naturalist 178: 715–725.
Wei Y, Terns RM, Terns MP . (2015). Cas9 function and host genome sampling in Type II-A CRISPR–Cas adaptation. Genes & Development 29: 356–361.
Westra ER, van Houte S, Oyesiku-Blakemore S, Makin B, Broniewski JM, Best A et al. (2015). Parasite Exposure Drives Selective Evolution of Constitutive versus Inducible Defense. Current Biology 25: 1043–1049.
Whitman WB, Coleman DC, Wiebe WJ . (1998). Prokaryotes: The unseen majority. Proceedings of the National Academy of Sciences 95: 6578–6583.
Wigington CH, Sonderegger D, Brussaard CPD, Buchan A, Finke JF, Fuhrman JA et al. (2016). Re-examination of the relationship between marine virus and microbial cell abundances. Nature microbiology 1: 15024.
Wilhelm SW, Suttle CA . (1999). Viruses and nutrient cycles in the sea: Viruses play critical roles in the structure and function of aquatic food webs. BioScience 49: 781.
Wommack KE, Colwell RR . (2000). Virioplankton: viruses in aquatic ecosystems. Microbiology and molecular biology reviews : MMBR 64: 69–114.
Yosef I, Goren MG, Qimron U . (2012). Proteins and DNA elements essential for the CRISPR adaptation process in Escherichia coli. Nucleic Acids Research page gks216.
Yoshida T, Ellner SP, Jones LE, Bohannan BJM, Lenski RE, Hairston NG Jr . (2007). Cryptic Population Dynamics: Rapid Evolution Masks Trophic Interactions. PLOS Biol 5: e235.
Acknowledgements
This work was supported by funding from the University of Maryland and the US Department of Education GAANN program. This material is based upon work supported in part by the US Army Research Laboratory and the US Army Research Office under contract/grant number #W911NF-14-1-0490. SM acknowledges funding from the Natural Sciences and Engineering Research Council of Canada (Discovery program). SM holds a T1 Canada Research Chair in Bacteriophages. PLFJ was supported in part by NIH R00 GM104158.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies this paper on The ISME Journal website
Supplementary information
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-sa/4.0/
About this article

Cite this article
Weissman, J., Holmes, R., Barrangou, R. et al. Immune loss as a driver of coexistence during host-phage coevolution. ISME J 12, 585–597 (2018). https://doi.org/10.1038/ismej.2017.194
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ismej.2017.194
This article is cited by
-
CRISPR-Cas provides limited phage immunity to a prevalent gut bacterium in gnotobiotic mice
The ISME Journal (2023)
-
Ecological memory preserves phage resistance mechanisms in bacteria
Nature Communications (2021)
-
The network structure and eco-evolutionary dynamics of CRISPR-induced immune diversification
Nature Ecology & Evolution (2020)
-
The arms race between bacteria and their phage foes
Nature (2020)
