
Abstract
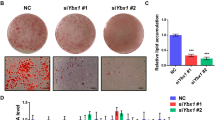
Indole-3-carbinol (I3C), a natural product of Brassica vegetables such as broccoli and cabbage, inhibits proliferation and induces apoptosis in various cancer cells. I3C has recently received attention as a possible anti-obesity agent. However, how I3C interacts with specific targets in the pathways involved in obesity and metabolic disorders is unknown. Silent mating type information regulation 2 homolog 1 (SIRT1), a NAD+-dependent deacetylase sirtuin, has recently emerged as a novel therapeutic target for metabolic diseases. Herein, we report that I3C is a potent, specific SIRT1 activator efficacious in cultured 3T3-L1 cell lines. A pull-down assay showed that I3C binds to SIRT1. To assess the significance of this binding, we determined whether I3C could activate SIRT1 deacetylase activity in a cell-free system. We found that I3C binds to SIRT1 and activates SIRT1 deacetylase activity in 3T3-L1 cells. In addition, I3C did not inhibit adipocyte differentiation in 3T3-L1 cells in which SIRT1 was knockdowned. Further, reverse transcriptase polymerase chain reaction analysis showed that I3C treatment reduced mRNA levels of adipogenic genes that encode for C/EBPα, PPARγ2, FAS, and aP2 in 3T3-L1 cells but not in SIRT1 knockdown cells. Overall, these results suggested that I3C ameliorates adipogenesis by activating SIRT1 in 3T3-L1 cells.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout


Similar content being viewed by others

References
Chinni SR, Li Y, Upadhyay S, Koppolu PK, Sarkar FH . Indole-3-carbinol (I3C) induced cell growth inhibition, G1 cell cycle arrest and apoptosis in prostate cancer cells. Oncogene 2001; 20: 2927–2936.
Aggarwal BB, Ichikawa H . Molecular targets and anticancer potential of indole-3-carbinol and its derivatives. Cell Cycle 2005; 4: 1201–1215.
Chang HP, Wang ML, Chan MH, Chiu YS, Chen YH . Antiobesity activities of indole-3-carbinol in high-fat-diet-induced obese mice. Nutrition 2011; 27: 463–470.
Chang HP, Wang ML, Hsu CY, Liu ME, Chan MH, Chen YH . Suppression of inflammation-associated factors by indole-3-carbinol in mice fed high-fat diets and in isolated, co-cultured macrophages and adipocytes. Int J Obes (Lond) 2011; 35: 1530–1538.
Choi Y, Kim Y, Park S, Lee KW, Park T . Indole-3-carbinol prevents diet-induced obesity through modulation of multiple genes related to adipogenesis, thermogenesis or inflammation in the visceral adipose tissue of mice. J Nutr Biochem 2012. e-pub ahead of print 7 May 2012 doi: 10.1016/j.jnutbio.2011.12.005.
Reagan-Shaw S, Nihal M, Ahmad N . Dose translation from animal to human studies revisited. FASEB J 2008; 22: 659–661.
Reed GA, Peterson KS, Smith HJ, Gray JC, Sullivan DK, Mayo MS et al. A phase I study of indole-3-carbinol in women: tolerability and effects. Cancer Epidemiol Biomarkers Prev 2005; 14: 1953–1960.
Shimamoto K, Dewa Y, Ishii Y, Kemmochi S, Taniai E, Hayashi H et al. Indole-3-carbinol enhances oxidative stress responses resulting in the induction of preneoplastic liver cell lesions in partially hepatectomized rats initiated with diethylnitrosamine. Toxicology 2011; 283: 109–117.
Haigis MC, Sinclair DA . Mammalian sirtuins: biological insights and disease relevance. Annu Rev Pathol 2010; 5: 253–295.
Fischer-Posovszky P, Kukulus V, Tews D, Unterkircher T, Debatin KM, Fulda S et al. Resveratrol regulates human adipocyte number and function in a Sirt1-dependent manner. Am J Clin Nutr 2010; 92: 5–15.
Baur JA, Pearson KJ, Price NL, Jamieson HA, Lerin C, Kalra A et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006; 444: 337–342.
Murata M, Ide T, Hara K . Reciprocal responses to dietary diacylglycerol of hepatic enzymes of fatty acid synthesis and oxidation in the rat. Br J Nutr 1997; 77: 107–121.
Gerhart-Hines Z, Rodgers JT, Bare O, Lerin C, Kim SH, Mostoslavsky R et al. Metabolic control of muscle mitochondrial function and fatty acid oxidation through SIRT1/PGC-1alpha. EMBO J 2007; 26: 1913–1923.
Yeung F, Hoberg JE, Ramsey CS, Keller MD, Jones DR, Frye RA et al. Modulation of NF-kappaB-dependent transcription and cell survival by the SIRT1 deacetylase. EMBO J 2004; 23: 2369–2380.
Zhang J . The direct involvement of SirT1 in insulin-induced insulin receptor substrate-2 tyrosine phosphorylation. J Biol Chem 2007; 282: 34356–34364.
Picard F, Kurtev M, Chung N, Topark-Ngarm A, Senawong T, Machado De Oliveira R et al. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-gamma. Nature 2004; 429: 771–776.
Acknowledgements
This work was supported by a grant of the Korea Health 21 R&D Project, Ministry of Health & Welfare (#090282), Republic of Korea and by the SRC program of the Korea Science and Engineering Foundation (KOSEF) grant funded by the Korea government (#2012-0000643).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Competing interests
The authors declare no conflict of interest.
Additional information
Supplementary Information accompanies the paper on International Journal of Obesity website
Supplementary information
Rights and permissions
About this article
Cite this article
Choi, Y., Um, SJ. & Park, T. Indole-3-carbinol directly targets SIRT1 to inhibit adipocyte differentiation. Int J Obes 37, 881–884 (2013). https://doi.org/10.1038/ijo.2012.158
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ijo.2012.158
