
Abstract
TCF4 (transcription factor 4; E2-2, ITF2) is a transcription factor that when haplo-insufficient causes Pitt–Hopkins Syndrome (PTHS), an autism-spectrum disorder that is associated with pervasive developmental delay and severe intellectual disability. The TCF4 gene is also a risk factor with highly significant linkage to schizophrenia, presumably via overexpression of the TCF4 gene product in the central nervous system. This review will present an overview of the clinical manifestations of PTHS and relate those clinical attributes to the underlying molecular genetics of TCF4. In order to provide a molecular biological context for the loss of function of TCF4 in PTHS, the review will also present a brief overview of the basic biochemistry of TCF4-mediated regulation of cellular and neuronal gene expression. In the final section of this review, I will discuss and speculate upon possible roles for the TCF4 transcription factor in neuronal function and comment upon how understanding these roles may give new insights into the molecular neurobiology of human cognition.
Similar content being viewed by others

Introduction and overview
Pitt–Hopkins Syndrome (PTHS) is a rare disorder characterized by intellectual disability (ID), ‘atypical’ autistic characteristics, and hyperventilation. PTHS is caused by heterozygous hypomorphic or null mutation or deletion of the transcription factor 4 (TCF4) gene on human chromosome 18; this underlying genetic basis of PTHS has only recently been discovered, leading to a paucity of mechanistic investigation thus far concerning the disorder. Indeed, even the clinical natural history of the syndrome is still being characterized fully, and PTHS patients have, in the past, been mis-diagnosed as either Rett Syndrome or Angelman Syndrome (AS) patients.1, 2 The identification of the dysfunctional TCF4 transcription factor gene as the genetic basis of the disorder was a critical step forward in beginning to understand the diagnosis, etiology and molecular biology of PTHS.3, 4, 5 The first two-thirds of this review will focus both on the clinical picture of PTHS, describing symptoms and manifestations of the syndrome, and on the fundamental molecular biology of the TCF4 transcription factor and its disruption in PTHS.
Due to the rarity of previous studies in the literature concerning the underlying molecular neurobiology of PTHS, there is a vast gap in our understanding of how loss of the TCF4 gene product leads to the cellular and neuronal dysfunction that underlies PTHS. For this reason, in the final section of this review, I will speculate in several instances about possible neurobiological mechanisms potentially at play in PTHS. I do not seek to try to establish these speculations as anything approaching fact but rather present them to provoke interest, thought and new hypotheses concerning this much-understudied disorder.
Some terminology related to PTHS
It is worthwhile to start with a few comments concerning commonly used abbreviations related to PTHS and its underlying causative gene, TCF4. Pitt–Hopkins Syndrome is routinely abbreviated as PTHS in order to allow distinction and disambiguation from the unrelated disorders Pallister–Hall Syndrome and Parkinsonism-Hyperpyrexia Syndrome, both of which are abbreviated as PHS. The use of PTHS as apposed to PHS as the abbreviation for Pitt–Hopkins Syndrome thereby clarifies the relevant clinical and basic science literature. Thus, it is highly desirable for workers reporting on Pitt–Hopkins Syndrome presently and in the future to adopt the PTHS abbreviation convention, while the PTHS literature is still relatively young and of an easily manageable size: at present a PubMed search of ‘Pitt–Hopkins Syndrome’ returns only 45 total references, while ‘Parkinsonism Hyperpyrexia Syndrome’ returns 16 publications, and ‘Pallister Hall Syndrome’ 125 publications.
Although the PTHS versus PHS convention is widely (but not universally) adhered to in the scientific literature, unfortunately the same cannot be said for the abbreviation TCF4. TCF4 (HUGO Standard Nomenclature=TCF4; Entrez Gene ID 6925; ensemble ENSG00000196628) is the standard and scientifically accepted abbreviation for Transcription Factor 4, the gene that when mutated causes PTHS and which also is a schizophrenia susceptibility gene. TCF4 is also known as Immunoglobulin Transcription Factor 2 (ITF2) and E2-2 in association with its status as an E-protein (Ephrussi-box binding) transcription factor. When searching the literature and perusing publications related to TCF4, great care should be taken not to confuse TCF4 with TCF7L2 (Transcription Factor 7-Like 2), another transcription factor mapping to a different genetic locus, on chromosome 10q25-25.3.6 TCF7L2 is also know as T-Cell Factor 4 and thus is also commonly abbreviated TCF4: the TCF7L2 gene product is not involved in PTHS but rather has a key role in the Wnt signal transduction pathway and may be involved in cancer and other diseases. To further complicate matters, TCF7L2 has also been genetically associated with schizophrenia, as has TCF4. Entering TCF4 into any standard search engine at this point in time will return a large mixture of references, the majority of which will be regarding TCF7L2 (T-Cell Factor 4) and not TCF4/E2-2/ITF2. For this reason, authors writing about TCF4 sometimes will specify TCF4 as TCF4 (E2-2, ITF2) to help clarify exactly which TCF4 is under discussion, as I did in the first line of the abstract for this review.
The clinical picture of PTHS
PTHS (OMIM 610954) was first described by Pitt and Hopkins,7 when they reported two unrelated patients with mental retardation, wide mouth and intermittent hyper-respiration. Subsequent independent reports confirmed the initial clinical spectrum as a distinct entity and led to the formalization of a disorder thereafter referred to in the literature by the name ‘Pitt–Hopkins Syndrome’.3, 4, 5, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 PTHS is extremely rare and, as of this writing, approximately 200–300 diagnosed cases are known to exist worldwide.30, 31, 32 Thus, PTHS falls in the category of an ‘ultra-orphan’ disease for purposes of FDA (Food and Drug Administration) approval for novel therapies under the Orphan Drug Act of 1983, and in some cases special funding programs for orphan disease patient treatment is available through government, biotech and pharmaceutical industry programs.
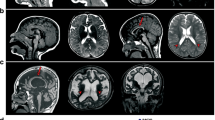
In modern clinical diagnosis, PTHS falls into the broad category of Pervasive Developmental Disorders and is a syndromic disorder characterized by severe ID, extensive developmental delay, absent speech, distinct facial features (for example, a gestalt of wide ‘cupid’s bow’ mouth, fleshy lips, broad nasal bridge and anteverted nares) and periods of hyperventilation followed by apnea.1, 33, 34 Thus in typical childhood clinical presentation in addition to the facial gestalt, the PTHS phenotype comprises various combinations of the following characteristics: pronounced developmental delay, an absence of speech development, normal growth parameters at birth, postnatal microcephaly, breathing abnormalities, motor uncoordination, ocular anomalies, including nystagmus, constipation, seizures and ‘atypical’ autistic behaviors (see Table 1).19 Additional common features include hypotonia and developmental delay in walking and slow development of fine motor control. Subtle brain anatomical abnormalities can include hypoplasia of the corpus callosum, enlarged ventricles and thin hindbrain.35, 36 Other magnetic resonance imaging features described with PTHS can include bulging caudate nuclei and a small hippocampus.27 As the individual matures, two of the most prominent aspects of PTHS are severe ID and pronounced language impairment—most PTHS patients develop little or no language utilization over their lifespan.
Epilepsy occurs frequently in PTHS and can be severe (see Table 2).19, 37 Based upon a review of the literature coupled with a genetic screening approach, Rosenfeld et al. 15 concluded that individuals with missense TCF4 mutations are more likely to develop seizures versus other types of TCF4 mutation. These authors also concluded that the onset of seizures occurs in the first decade of life in most reported patients with PTHS.
The most common extra-neurological manifestation of Pitt–Hopkins Syndrome is gastrointestinal disturbance, especially constipation, which may be medically significant and often requires medication. PTHS patients also can have ocular abnormalities in the form of strabismus, myopia and astigmatism.19, 37
A characteristic breathing abnormality in this syndrome, as was described even in the earliest descriptions of PTHS,7 can appear in mid-childhood and consists of abrupt paroxysms of hyperventilation (tachypnea) followed by breath holding and even overt cyanosis. Hyperventilation can be followed by apnea, but apnea can also occur independent from hyperventilation.22, 24, 29, 38 However, one very recent report identified a bona fide case of Pitt–Hopkins Syndrome, confirmed by genetic diagnostics, of a 12-year-old boy presenting with psychomotor retardation, recurrent respiratory tract infections and typical dysmorphic features but with absence of hyperventilation or other breathing abnormalities. This case suggests that breathing anomalies will not be universally associated with PTHS. Consistent with this conclusion, Rosenfeld et al. 15 screened 13 186 individuals with microarray-based comparative genomic hybridization and coupled that with a review of the literature. Their statistical analysis of the phenotypic features for all individuals with confirmed mutations or deletions of TCF48, 15 identified seven individuals with TCF4 deletions yet only three had breathing anomalies. All seven individuals had psychomotor retardation consistent with Pitt–Hopkins Syndrome.
As described in more detail by Van Balkom et al.,39 PTHS is an autism-spectrum disorder (ASD) and the autistic characteristics of PTHS are a crucial aspect of the syndrome (see Table 3). Specifically, PTHS falls in the category of pervasive developmental disorders—not otherwise specified. As will be discussed below, PTHS is phenotypically quite similar to AS, and both fall into the autism spectrum based on lack of communication, pervasive developmental delay, ID and perseverative behaviors.14 However, both AS and PTHS are syndromic disorders and these patients often do not manifest the high degree of social withdrawal associated with autism per se, and PTHS and AS patients are thus referred to as manifesting ‘atypical’ autistic characteristics or syndromic autism.
Molecular diagnostics
Although the above phenotypic characterization of PTHS allows clinical diagnosis, definitive diagnosis of PTHS is achieved through testing of patient DNA. Current genetic diagnostic testing utilizes molecular techniques, such as whole-exome sequencing, and chromosomal analysis for gene duplication/deletion/breakage. Sequence analysis covering all 18 TCF4 coding exons (exons 2–19), splice junctions and immediate intronic flanking regions detects approximately 70% of PTHS mutations, whereas approximately 30% of TCF4 mutations are whole- or partial-gene deletions.2 Gene deletions can be detected by either Chromosome Microarray Analysis or cytogenetic testing or a combination of both. A list of diagnostic testing laboratories providing genetic screening for PTHS can be found at:
http://www.ncbi.nlm.nih.gov/sites/GeneTests/lab/clinical_disease_id/318881. A very helpful description of diagnostic testing procedures and parameters specifically related to PTHS can be found at:http://www.ncbi.nlm.nih.gov/books/NBK100240/.1
Clinical treatment
There is available only a sparse literature concerning clinical therapeutics for PTHS.1, 12, 22 Currently, there is no specific therapy for Pitt–Hopkins Syndrome, and indeed the inter-patient variability in clinical presentation necessitates customized treatment regimens based on symptomatology. In terms of pharmacological approaches, symptomatic treatment for epilepsy is of course necessary in patients manifesting this aspect of the disorder: valproate and other anticonvulsants have been used to control the epilepsy associated with PTHS. One report suggests that high-dose valproate may also be ameliorative for PTHS-associated breathing abnormalities.40 The carbonic anhydrase inhibitor Acetazolamide, which regulates blood pH and CO2 levels and as a secondary effect impacts respiratory rate, has been reported to elicit improvement in PTHS patients regarding episodic hyperventilation with apnea while awake.38 Finally, medical treatment of constipation is frequently necessary.
Especially given the lack of any specific pharmacotherapy for PTHS, the standard of care for PTHS patients calls for the utilization of behavioral and training approaches. Physical and occupational therapy are indicated for the motor un-coordination and ataxia frequently present in PTHS. As the patient matures, the lack of language development presents significant challenges, and speech therapy and enhanced electronic communication devices and attendant software such as Verbal Victor (download site: https://sites.google.com/site/verbalvictor/) should be prescribed to help in communication with clinicians, caregivers and family members. As PTHS is an ASD (see above), where possible Applied Behavioral Analysis should be made available as part of a comprehensive treatment approach.
Finally, the Pitt–Hopkins Support Group and the Pitt–Hopkins International Network can serve as a valuable source of practical information for families and caregivers as regards many aspects of PTHS therapeutics (Pitt–Hopkins Syndrome Support Group: http://groups.google.com/group/pitt-hopkins/about?pli=1; Pitt–Hopkins Syndrome International Network: http://pitthopkins.org/)
Differential diagnosis—discovery of the TCF4 transcription factor as the PTHS gene
Clinical differential diagnosis of PTHS includes distinguishing PTHS versus the following syndromes: AS, Rett Syndrome, Mowat–Wilson Syndrome, Joubert Syndrome, ATR-X-related ID syndrome, Neurexin 1 (NRXN1)-associated autosomal recessive ID disorder, and contactin-associated protein-like 2 (CNTNAP2)-associated autosomal recessive ID disorder. A more complete guide to differential diagnosis among these various possibilities is given in references 1, 34 and 35. Fortunately, all these disorders have genetic screening tests available in order to allow distinguishing among them, which I will discuss in more detail in the following section.
A landmark step forward for our understanding of PTHS came in 2007 with the simultaneous independent discovery by three different research groups in Europe that disruption of the TCF4 gene (TCF4, located at 18q21.1, (OMIM 602272)) is the basis for PTHS.40, 41, 42 These groups discovered that point mutations, intragenic deletions or broader deletions in Chromosome 18 that disrupt the TCF4 gene are diagnostic for PTHS, and thus by definition PTHS is a disorder of disrupted TCF4 function. These pioneering human genetic studies and a wide variety of subsequent studies have clearly demonstrated that heterozygous deficiency (haplo-insufficiency, or loss of function of one of the two cellular copies of TCF4) is sufficient to cause PTHS in humans. Mosaicism of TCF4 mutation, variability in deletion/insertion mutations and the cellular locales wherein the TCF4 deficiency resides is also an important when considering the variable phenotypes observed in PTHS patients.21, 41
The breakthrough discovery of TCF4 as the PTHS gene provided a rationale for clinical genetic screening for mutation of TCF4 as an unambiguous means of diagnosis of the syndrome. This is not a trivial consideration, as PTHS shares considerable phenotypic similarity with the (presumably) more broadly occurring Angelman Syndrome (AS, (OMIM 105830)), Rett Syndrome (RTT, OMIM 312750) and Mowat–Wilson Syndrome (MWS, OMIM 235730).14, 33, 35 Clinically, one should consider Angelman, Rett and Mowat–Wilson Syndromes in the differential diagnosis of Pitt–Hopkins Syndrome because of the overlapping phenotype of severe ID with absent speech, epilepsy, microcephaly and constipation.
Indeed, a recent study has demonstrated that in a quasi-random sampling of patients diagnosed with AS, approximately 2% of individuals thought to have AS instead had TCF4 deficiency, that is, PTHS.14 Regarding Rett Syndrome, Armani et al. 42 identified a frameshift mutation in TCF4 in a patient who had previously been diagnosed clinically with ‘variant’ RTT. Takano et al. 33 have also suggested that PTHS be included in the differential diagnosis of X-linked α-thalassemia Intellectual Disability (ATR-X (OMIM 301040)) due to phenotypic similarities to this syndrome, as well as identifying at least one bona fide case of mis-diagnosis of a presumed ATR-X patient under their care. Lehalle et al. 36 have observed prominent fetal pads, on the fingers and toes, as a feature in several individuals with PTHS and have suggested that this phenotype can be used to help distinguish PTHS from other similar conditions during differential diagnosis, and thus this characteristic may be useful in suggesting specific genetic testing for PTHS.
Given the rarity of PTHS itself, co-morbidity with another genetic disorder is indeed highly unlikely. However, Ghosh et al. 37 have observed the co-occurrence of Charcot-Marie-Tooth Disease Type 1 (CMT1A, OMIM 118220) with PTHS in one individual, with PTHS diagnosis confirmed by genetic screening.
Although the majority of cases of PTHS arise as a result of sporadic, spontaneous mutations in the TCF4 gene, Steinbusch et al. 43 have reported somatic mosaicism of the TCF4 gene in a mother with two children manifesting PTHS. In both children, a heterozygous frameshift mutation (c.1901_1909delinsA, p.Ala634AspfsX67) was found in exon 19 of TCF4: the same mutation was found at low levels in DNA from the mother. As Steinbusch et al. 43 suggest, the possibility of familial recurrence with somatic mosaicism in a healthy mother has important consequences for genetic counseling of PTHS families. This consideration may also explain an early report of possible PTHS in two siblings,28 although an alternative explanation could be the more recently characterized possibility of manifestation of a PTHS-like ID disorder manifesting autosomal recessive inheritance through NRXN1 and CNTNAP2 mutations, as will be described in the next paragraph.
Differential diagnoses versus novel ‘Pitt–Hopkins-like’ syndromes
Recent studies of a few patients16, 44, 45, 46 have demonstrated the existence of two autosomal recessive disorders that are characterized by phenotypes very similar to PTHS: NRXN1-associated intellectual disability disorder, and CNTNAP2-associated intellectual disability disorder. This has led to these two disorders being categorized as Pitt–Hopkins-like syndromes.30 Indeed it has been hypothesized that TCF4, NRXN1 and CNTNAP2 (a Neurexin homolog) may all lie within a common signaling pathway that when disrupted leads to PTHS and related ID disorders.16, 44 Similar to PTHS, both NRXN1-associated ID disorder and CNTNAP2 -associated ID disorder manifest pervasive developmental delay, lack of speech, stereotypic movements and episodic hyperventilation or breath-holding. In one instance, a patient in this category exhibited epilepsy. However, these individuals lack the characteristic facial features seen in PTHS.
CNTNAP2 and NRXN1 are two distantly related members of the neurexin superfamily, and disruption of these genes has been implicated in a wide spectrum of neuropsychiatric disorders, such as developmental language disorders, ASDs, epilepsy and schizophrenia.16 In pioneering studies, Zweier et al. 16 identified deletions and mutations in CNTNAP2 and NRXN1 in four patients with severe ID and variable features, such as autistic behavior, epilepsy and breathing anomalies, phenotypically overlapping with Pitt–Hopkins Syndrome. Although PTHS is an autosomal dominant disorder caused by haplo-insufficiency in the TCF4 gene, NRXN1- and CNTNAP2-associated ID are autosomal recessive disorders.16, 44 Interestingly, the known function of NRXN1 to serve as a synaptic cell adhesion molecule linking the presynaptic terminal with the postsynaptic compartment suggests that synaptic defects contribute to NRXN1-associated ID and by implication CNTNAP2-associated ID and PTHS as well.16, 44 Especially intriguing is the linkage of CNTNAP2 as an autism-susceptibility gene47, 48, 49 and its known contribution as a gene controlling human auditory language cognition.50, 51
The biochemistry of TCF4
Mutations in PTHS—molecular genetics of the disorder
In this section, I will review the basic biochemistry and regulation of TCF4 and cover some basics of its structure–function relationships and how they are affected in PTHS. This topic has recently been covered in an excellent review by Navarette et al.,6 so I will not reiterate all the particulars here. The review by Navarette et al. is notable for its meticulous but comprehensive review of the TCF4 literature, including having taken great care to disambiguate TCF4 from T-Cell Factor 4/TCF7L2.
As already mentioned, PTHS is a neurodevelopmental disorder, the underlying genetic basis of which is mutation/deletion of the TCF4 gene and resultant disruption of normal TCF4 function. TCF4 is a transcription factor active in the developing and mature central nervous system (CNS) whose function is to regulate gene readout (transcription) from the genome, and TCF4 has a large number of potential gene targets that it might regulate. The TCF4 gene itself is quite large and complex, spanning 437 kb with approximately 40 exons and a corresponding number of intervening introns. The literature is replete with synonyms for TCF4, including: E2-2, ITF2, PTHS, SEF2, SEF2-1, SEF2-1A, SEF2-1B, bHLHb19, MGC149723 and MGC149724.
The mutational spectrum of TCF4 in PTHS includes large chromosomal deletions encompassing the whole TCF4 gene, partial gene deletions, frameshift (including premature stop codon), nonsense, splice site and missense mutations. So far, almost all the patients have a private mutation and only two recurrent mutations are known.22 Missense mutations are generally within the basic helix-loop-helix domain (bHLH domain), a mutational hotspot. Consistent with the hypothesis that the causative loss of function relevant to PTHS is a decline in gene transcriptional regulation, missense mutations in this DNA regulatory element-binding domain of TCF4 account for approximately 15% of PTHS cases. Point mutations, mostly causing premature stop codons, account for 40% of PTHS cases. It is unlikely that PTHS caused by these types of mutations can be accounted for by a mechanism other than loss of TCF4-mediated transcriptional regulation.
Basic molecular biology of TCF4
TCR4 is expressed in several tissues during human development, particularly in the CNS involving telencephalon, diencephalon and cerebellum.6 Extraneurally, TCF4 is expressed in the peribronchial and kidney mesenchyme. It is also expressed in the limb and genetic buds, the splanchopleural mesenchyme and the sympathetic, parasympathetic and enteric ganglia.6 Tissue specificity of TCF4 gene expression, along with its biochemically documented dimerization with other bHLH proteins, each of which partners has its own select tissue distribution, may explain the characteristic non-CNS based clinical manifestations of this disease, like typical dysmorphism, incoordination, constipation and muscle hypotonia.8, 9, 10, 11, 13, 17, 26
TCF4 is a bHLH transcription factor. The encoded protein recognizes an Ephrussi-box (‘E-box’)-binding site (‘CANNTG’)—a motif first identified in immunoglobulin enhancers. In the immune system, the gene is expressed predominantly in pre-B-cells, although it is found in other tissues as well.20, 23 Multiple alternatively spliced transcript variants that encode different proteins have been described.52, 53, 54, 55 An important recent study by Timmusk and colleagues52, 55 investigated the role of alternatively spliced transcripts of the human TCF4 gene in regulating the structure, expression and coding potential of TCF4 gene. The Timmusk group found that human TCF4 mRNA expression is particularly high in the brain, and found that usage of numerous 5′ exons of the human TCF4 gene potentially results in human TCF4 protein isoforms with 18 different N-termini. They also observed alternative splicing of several internal exons, which resulted in even greater transcript diversity. In terms of functional consequences, different splice variants resulted in differential subcellular distribution of TCF4 isoforms: some isoforms contained a bipartite nuclear localization signal and were exclusively nuclear, whereas others were more widely distributed throughout the cell.
E-box transcription factors such as TCF4 have no known DNA-binding activity as monomers but rather form homodimers or heterodimers with other proteins in order to enable DNA binding and activate gene expression.53, 54 Because each binding partner contributes a specific DNA recognition half-site, different heterodimers can provide distinct E box-binding specificities—this fact makes in silico analysis of potential TCF4 target genes problematic. It also is important to note that depending on which binding partner TCF4 binds to, it may be either a transcription activator or a transcription suppressor. In a recent study, Sepp et al. 55 found, using in vitro methods, that the subcellular distribution of TCF4 relies on its heterodimerization partners. Furthermore, Sepp et al. found that the ability of different TCF4 splice-variant isoforms to regulate E-box-controlled reporter gene transcription varied depending on whether one or both of the two TCF4 transcription activation domains were present in the protein.
As a transcription factor, TCF4’s function is to regulate gene expression. Normally transcription factors lie dormant in the nucleus, subject to activation by signaling pathways upstream of them. Thus, upon activation TCF4 then binds to its appropriate DNA response elements found in the upstream regions of genes, in order to regulate transcription of downstream target genes. The upstream mechanisms controlling TCF4 activation are largely unknown at this point. However, one documented mechanism for TCF4 regulation is that binding by calcium/calmodulin56 results in inhibition of DNA binding in vitro: an increased intracellular Ca2+ concentration potently inhibits the transcriptional activity of TCF4 on an E-box-containing reporter plasmid, as does calmodulin overexpression. Thus Ca2+ signaling may inhibit the transcriptional activity of E-proteins, including TCF4, by establishing an E-protein–calmodulin complex that prevents the E-protein from interaction with its target DNA.
As TCF4 was identified as a regulator of immune cell function and much of the published work relates to that role, not much is known concerning the functional roles of TCF4 in the nervous system (see Figure 1).1 TCF4 is known to be important for regulating glial cell differentiation, especially the maturation of oligodendrocyte progenitors. During neurodevelopment TCF4 is known to interact with several class II bHLH transcription factor genes such as Math1, HASH1, neuroD2, Id1 and Olig2. In the brain, TCF4 is expressed in both glial cells (oligodendrocytes) and neurons.57 In the human brain, TCF4 has the highest levels of expression in neocortex, hippocampus, striatum, thalamus and cerebellum.1, 57

Proposed TCF4-related signaling pathways in the nervous system. Major known mechanisms of TCF4 functional regulation include: alternative splicing, dimerization partners and calcium/calmodulin-dependent inhibition. Functional targets are virtually unknown, but possibilities include neurexin 1 (NRXN1) and contactin-associated protein-like 2 (CNTNAP2). Please note that this speculative figure is agnostic on whether the proposed signaling pathways reside in glia, neurons or both.
In terms of specific molecular targets, Forrest et al. 44 showed that TCF4 can modulate the expression of NRXN1 and CNTNAP2, using cell culture systems in vitro (Figure 1). Forrest et al. tested a panel of PTHS-associated mutations and found that mutations in the bHLH domain of TCF4 alter the subnuclear localization of TCF4 and regulated homo- and heterodimer formation. They also found that TCF4 can transactivate the NRXN1β and CNTNAP2 promoters using in vitro fluorescent reporter constructs and using this experimental system observed variable, context-specific deficits in the ability of the different PTHS-associated TCF4 mutants to activate gene transcription when coexpressed with different bHLH transcription factors. These interesting observations demonstrated that PTHS-associated missense mutations can have multiple effects on the function of TCF4 and, as described above, suggest that TCF4 may modulate the expression of NRXN1 and CNTNAP2, thereby defining a regulatory network in PTHS and PTHS-like ID syndromes. This important finding hopefully will propel extensive investigation of this possibility in the future.
Structure–function relationships for PTHS-associated mutations in TCF4
As regards the functional consequences of specific PTHS-associated mutations on TCF4 transcriptional activation, in a pioneering study Sepp et al. 52, 55 evaluated the functional impact of a wide variety of published PTHS-associated TCF4 mutations. In this comprehensive study, the Timmusk group took into account the diversity of TCF4 isoforms and assessed how the known PTHS-associated reading frame elongating and missense mutations affected TCF4 function. Their analysis revealed that not all deletions and truncating mutations in TCF4 result in complete loss-of-function and the impact of reading frame elongating and missense mutations ranges from subtle deficiencies to dominant-negative effects. Specifically, in overview they found that: (1) missense mutations in TCF4 bHLH domain and the reading frame elongating mutations diminished DNA-binding and transactivation ability in a manner dependent on dimer context (homodimer versus heterodimer with ASCL1 or NEUROD2); (2) the elongating mutation and the missense mutations at the dimer interface of the HLH domain destabilized the protein; and (3) missense mutations outside of the bHLH domain caused no major functional deficiencies in their in vitro cellular assay system. Based on these findings, Timmusk and colleagues concluded that different PTHS-associated mutations impair the functions of TCF4 by diverse mechanisms and to a varying extent and that these factors are likely to contribute to the phenotypic variability seen in PTHS patients.
Regulation of TCF4 by miRNA137
TCF4 has also recently been identified as a target for regulation by the microRNA (miRNA) system, specifically by miR-137.58, 59, 60 miRNAs are small non-coding RNAs that typically function as negative regulators of gene expression through their direct interaction with target mRNAs. MiRNAs have lately generated a high degree of interest in the schizophrenia field, and miR-137 regulation of TCF4 was discovered in this context. Thus, as the product of a large-scale genome-wide association study of schizophrenia, five schizophrenia-susceptibility loci (TCF4, CACNA1C, CSMD1, C10orf26, ZNF804A) were experimentally validated as miR-137 targets.59, 60 Based on these recent studies, miRNA-based regulation of cellular TCF4 levels by miR-137 is emerging as a potential candidate mechanism, although this mechanism has not been investigated in the context of PTHS. Assuming that miR-137 acts as a negative regulator of TCF4 in the nervous system, one might speculate that antagonizing miR-137 function could upregulate TCF4 levels in the brain.
A hypothesis—disruption of neuronal activity-dependent gene transcription as the neurobiological basis of PTHS
Thus far in this review, I have covered two areas critically important in understanding PTHS: the clinical spectrum of the disorder per se, and what is known about the molecular biology of the TCF4 gene and how it is disrupted in PTHS patients. However, it is clear that a vast gap exists in our knowledge in terms of bridging between these two conceptual areas. We have some understanding of the human clinical picture in PTHS and the beginnings of a model for TCF4 function in the cell, but the molecular neurobiology whereby loss of TCF4 leads to disruption of neuronal function, cognition and behavior has been essentially untouched to date. The final sections of this review will comprise hypotheses, speculations and editorial comments concerning the potential neurobiological basis of PTHS. The purpose of these final sections is to begin to develop a conceptual basis for how one, through experimentation, might advance our understanding of the neurobiological underpinnings of PTHS, the functional roles of TCF4 in the CNS. In addition, I will comment upon how these studies might expand our knowledge of the molecular neurobiology of human cognition, ID and ASDs and facilitate development of novel therapies for PTHS.
A developmental or ongoing need for TCF4?
PTHS is a neurodevelopmental disorder, the underlying genetic basis of which is mutation/deletion of the TCF4 gene. The mutated gene product is present throughout development but is also present in the fully developed adult CNS. It is unclear whether PTHS is caused exclusively by disruption of TCF4 function during development or whether loss of TCF4 in the mature CNS might also contribute to neurobehavioral and cognitive dysfunction in PTHS patients.61 Recent data from studies of a number of different ‘developmental’ disorders such as Rett Syndrome, AS and Fragile X Mental Retardation has suggested that loss of normal gene function in the post-developmental nervous system contributes to cognitive and neurobehavioral dysfunction in these disorders. Addressing this question in the case of PTHS is critically important because of the implications for developing potential new treatments for PTHS. If TCF4 functions to control cognition and synaptic function in the mature CNS, cognitive dysfunction in PTHS might, in significant part, be due to the disruption of TCF4’s actions in the fully developed CNS. A new understanding of the role of TCF4 in the adult CNS might allow the development of new therapeutic approaches to PTHS treatment based on restoration or augmentation of TCF4 function after CNS development is largely finished. Alternatively, if PTHS is found to be largely developmental in etiology, future drug development would need to focus on reversing developmental and structurally based deficits, potentially through re-opening developmental critical periods.
Neurobiological studies of PTHS
The identification of the dysfunctional TCF4 transcription factor gene as the genetic basis of the disorder is a critical step forward in beginning to understand the diagnosis, etiology and molecular biology of PTHS.3, 4, 5 A critical need at this point is the pursuit of laboratory studies to investigate the cognitive dysfunction associated with PTHS, focusing on mechanistic studies to understand the role of the TCF4 transcription factor in CNS function, utilizing laboratory animals and in vitro model systems. There is no doubt that the somatic, non-CNS-based aspects of PTHS are vitally important; however, I propose a need to focus on the cognitive and CNS-based aspects of PTHS in the post-developmental CNS for three reasons. First, the cognitive dysfunction associated with PTHS is profound and extremely debilitating, making this a priority area for drug development. Second, drug treatments are unlikely to be able to completely reverse developmental structural and anatomical defects but are likely to be effective in restoring CNS function if the underlying etiology of PTHS cognitive dysfunction is due to a lack of the ongoing presence of the PTHS gene product in the post-developmental CNS. Third, understanding the role of gene transcriptional regulation in cognitive function is a rapidly expanding area of biomedical research at present and is the area where research might best be able to make a rapid meaningful impact. Overall, characterizing the behavioral and physiological effects of loss of TCF4 function is necessary not only in order to understand PTHS etiology but also to allow for drug screening in the future.
One additional scientific objective in undertaking studies of the molecular neurobiology of PTHS is to test the hypothesis that TCF4 is necessary for normal cognitive function in the fully developed mature CNS. Thus, research is necessary to assess the role of the bHLH transcription factor TCF4 in adult learning and memory, hippocampal synaptic plasticity, and synaptic function. Overall, in terms of basic cognitive neurobiological studies, a central hypothesis that needs to be tested at present is the hypothesis that TCF4 regulates the brain’s ability to trigger long-term synaptic plasticity and memory formation by actively regulating transcriptional activity during learning.
Available genetically engineered mouse models of PTHS
In advancing our understanding of both the clinical and cognitive neurobiology of PTHS, transgenic and knockout mouse models are an extremely valuable resource. A genetically engineered ‘traditional’ knockout mouse with heterozygous deletion of TCF4 is currently commercially available from the Jackson Laboratories (Bar Harbor, ME, USA), and this mouse line models the lifelong TCF4 haplo-insufficiency of PTHS patients. Thus, this model can be used to study developmental-plus-mature loss of TCF4 function.
However, in the knockout mouse line, the function of the mutated gene product is not only absent throughout development but is also absent in the fully developed young and adult CNS. As discussed above, it is unclear whether PTHS is caused exclusively by disruption of TCF4 function during development or whether loss of TCF4 in the mature CNS might also contribute to neurobehavioral and cognitive dysfunction in PTHS patients. Addressing this question is critically important because of its implications for developing potential new treatments for PTHS. If TCF4 functions to control cognition in the mature CNS, cognitive dysfunction in PTHS might, in significant part, be due to disruption of TCF4’s actions in the fully developed CNS. A new understanding of the role of TCF4 in the fully developed CNS might allow the development of new therapeutic approaches to PTHS treatment based on restoration or augmentation of TCF4 function after CNS development is largely finished. Thus, if TCF4 function is necessary for learning and memory in the adult CNS, then drug treatments to augment TCF4 function are a viable therapeutic approach for PTHS patients of all ages.
In order to address the issue of an ongoing role for TCF4 in the mature CNS, genetically engineered mouse lines containing a floxed conditional allele of TCF4 are highly desirable. Fortunately, a floxed TCF4 allele mouse line has already been generated and is available. This line was generated by Allan Bradley’s group at the Sanger Institute in Cambridge and is available through the EMMA mouse repository. This conditional allele mouse line is referred to asTcf4tm1a(EUCOMM)Wtsi, which stands for: transcription factor 4; targeted mutation 1a, Wellcome Trust Sanger Institute (MGI ID: MGI:4432303). To generate this mouse line, the L1L2_gt1 cassette was inserted at position 69621025 of Chromosome 18 upstream of the transcription-regulating exons.62 The cassette is composed of an FRT flanked lacZ/neomycin sequence followed by a loxP site. An additional loxP site is inserted downstream of the targeted exons at position 69621797. Critical functional exons are thus flanked by loxP sites. A ‘conditional ready’ (floxed) allele can be created by flp recombinase expression in mice carrying this allele. Subsequent cre expression results in a knockout mouse. If cre expression occurs without flp expression, a lacZ reporter knockout mouse is created. This variation allows one to identify the cells in which TCF4 was deleted in vivo. Further information on targeting strategies used for this and other KnockOut Mouse Project (KOMP) alleles can be found at: http://www.knockoutmouse.org/aboutkompstrategies. An additional floxed allele of TCF4 has also been generated by the laboratory of Dan Holmberg, referred to as Tcf4tm1Hmb (transcription factor 4; targeted mutation 1, Dan Holmberg, MGI ID: MGI:3036170). For this mouse line, the exons encoding the bHLH and C-terminal domains are flanked by loxP sites, inserted by homologous recombination in ES cells.63
The availability of these additional ‘floxed’ TCF4 mice allow selective conditional deletion of the TCF4 gene in the post-developmental or adult CNS—this will allow investigators to generate mouse models to assess the role of TCF4 in adult learning and memory (through post-developmental knockout using the conditional allele). Furthermore, the availability of these mouse lines might allow the investigation of mice harboring a homozygous deletion of TCF4 in the adult CNS, as will be discussed in the next paragraph.
Initial attempts to create TCF4-null mice resulted in embryonic lethality. However, it is desirable to study the function of TCF4 in cells, including the CNS under conditions where the gene product is completely absent (homozygous deficiency). In part, this is because the effects of loss of TCF4 can be much more clearly identified when there is complete loss of the gene, as opposed to the hemizygous traditional knockout animals. To circumvent this problem, it is possible to use the Cre/LoxP recombination system to delete the TCF4 gene completely and in a tissue-specific manner but induce loss of the gene only after body and brain development is completed normally. The second type of mouse line(s) discussed above, the CRE/LoxP ‘floxed’ mouse line(s) can allow conditional homozygous deletion of the TCF4 gene in the mature CNS. Thus, in order to generate these conditional gene knockout animals the floxed TCF4 mouse line must be cross-bred with a different mouse line that expresses cre DNA recombinase preferentially in the CNS only after development. A variety of mouse lines of this sort are already available, for example, the CaMKII/cre mouse lines generated by the laboratories of Alcino Silva and Susumu Tonegawa. In this manner, selective conditional deletion of the TCF4 gene post-developmentally in only the mature CNS will allow assessment of the ongoing necessity of TCF4 for cognitive function in the adult brain.
Finally, another existing TCF4 mouse line has been generated, a transgenic overexpressor of TCF4. Although these animals are not a model for PTHS, published previous results from characterizing this mouse line are highly relevant as background for PTHS studies. Brzozka et al. 64 identified cognitive and sensorimotor gating impairments in transgenic mice overexpressing TCF4 in the brain. To investigate the possible function of TCF4 in the adult CNS, this group generated transgenic mice that moderately overexpress TCF4 postnatally in the brain to reduce the risk of developmental effects possibly interfering with adult brain functions. TCF4 transgenic mice were characterized with molecular, histological and behavioral methods. Brzozka et al. 64 found that TCF4 transgenic mice display profound deficits in contextual and cued fear conditioning and sensorimotor gating. Furthermore, they observed that TCF4 interacts with the neurogenic bHLH factors NEUROD and NDRF in vivo. Molecular analyses revealed dynamic circadian deregulation of neuronal bHLH factors in the adult hippocampus as well. The authors concluded that TCF4 likely acts in concert with other neuronal bHLH transcription factors, contributing to higher-order cognitive processing, and that moderate transcriptional deregulation of TCF4 in the brain through overexpression interferes with cognitive functions and might alter circadian processes in mice. These observations provided the first insights into the physiological function of TCF4 in the adult brain and its possible contributions to cognitive disorders. Overall, I view these studies as consistent with the hypothesis that TCF4 contributes in an ongoing way to memory function in the post-developmental CNS. Vis-à-vis the hypothesis that TCF4 is necessary for memory and that a loss of TCF4 will cause memory and plasticity deficits, I interpret the results of Brzozka as potentially being due to a dominant-negative effect of TCF4 overexpression in the CNS. This interpretation is consistent with what has become a standard model for MeCP2 function in the CNS, wherein both overexpression and gene deletion can lead to plasticity and memory deficits.65, 66, 67, 68, 69, 70
The existence of these knockout and transgenic mouse lines should allow a comprehensive behavioral and neurophysiological characterization of these mouse models relevant to PTHS, focusing on motor behaviors, learning, memory and synaptic function of neurons in the CNS. The behavioral and neurophysiological assessments can be performed using both the developmental (knockout/transgenic) and post-developmental (conditional allele) mouse models for PTHS. The use of these various models in concert will allow investigators to distinguish roles for the TCF4 gene in development versus ongoing function of the developed CNS. Subsequent in vivo studies of potential novel treatments for PTHS can also be undertaken in both the types of mouse lines. Functional assessment of the mouse lines and their potential use as drug screening platforms will be discussed in more detail in the following section.
Physiological and behavioral characterization of the available mouse models for PTHS
Contemporary learning and memory research has highlighted three themes. The first is the cellular basis of behavior, that is, that behaviors are rooted in the coordinated actions of neurons in the CNS. Second, that learning and memory are subserved by synaptic plasticity. Synapses are the connections between neurons, and learning and memory are mediated by alterations in synaptic connections in the CNS. Learning involves specific triggering events at synapses, which result in persistent biochemical changes that modify the strength of synaptic connections. A final theme that has emerged is the complexity of the molecular machinery underlying the changes in synaptic strength. Though this machinery is elaborate, and multicomponent, it operates in an orchestrated, carefully controlled fashion to achieve the proper end results. One critical and required component of long-term learning and memory is the regulation of gene transcription in the neuronal nucleus. As already described, I hypothesize that loss of TCF4 function leads to cognitive deficits in PTHS through disruption of the transcriptional regulation required for normal memory.
Fortunately, it is possible to test learning and memory behavior in the PTHS model mice described above. Much progress has been made recently in developing standardized procedures for behavioral learning and memory screening in mice.71 A fairly typical standard behavioral screen includes, in addition to basic assessments of such parameters as weight, temperature, general reflexes, and so on, evaluation of: open-field behavior, rotor-rod testing, pre-pulse inhibition, cued and contextual fear conditioning and visual- and hidden-platform tasks in the Morris water maze. This battery of tests allows evaluation of a variety of sensory responses, including hearing and vision, general activity, reflexes and motor coordination, motor learning, associative learning and spatial learning.
An additional component of a comprehensive characterization of a mouse model for any ID syndrome includes assessments of synaptic function and plasticity in the hippocampus. Part of the basis for these studies is the emerging consensus that synaptic plasticity in the hippocampus is involved in mammalian learning and memory. Thus, characterization of synaptic transmission, short-term synaptic plasticity and long-term potentiation (LTP) is important in terms of trying to understand the underlying cellular processes involved in learning and memory and any memory deficits observed in PTHS model mice. In this context, NMDA (N-methyl-D-aspartate) receptor-dependent LTP in area CA1 of the hippocampus is the standard model for LTP. This form of LTP is by far the most widely studied form of synaptic plasticity in the CNS and much more is understood concerning its biochemical mechanisms than is understood for the other forms of LTP.
Using model mice and in vitro systems as drug-discovery platforms for PTHS
The development of new valid targets for PTHS therapeutics is a pressing and compelling societal and biomedical issue and thus must proceed quickly. As already described, it is a virtual certainty that PTHS is caused by mutations resulting in a loss of TCF4-mediated transcriptional regulation. This is a very important consideration when proposing drug-development studies—in particular the drug screens should focus on finding regulators of the transcriptional activation triggered by TCF4. Moreover, more generally acting promoters of transcriptional activation such as histone de-acetylase inhibitors (HDACi) are also an appealing category of potential treatment for PTHS. HDACi, in general, act as ‘boosters’ of transcriptional activation and perhaps could compensate for the haplo-insufficiency of the TCF4 gene product, by augmenting the action of the remaining half-complement of TCF4 still present in PTHS patients. I will discuss these two different types of approaches in the following two sections.
In vitro drug screening
The first possible approach is developing in vitro systems for assessing TCF4 function that are suitable for application to high-throughput drug-screening procedures. Because of the complexity of these types of experiments and because very little relevant preliminary data are available, I will not try to describe these studies in detail for the present. The essence of this approach, however, is to develop in vitro procedures that can be used to assess the biochemical function of TCF4, in order to subsequently use these biochemical assays to screen chemical libraries for compounds that can augment TCF4 function. The screens can be based on the known function of TCF4 to regulate gene transcription. As already described, it is straightforward and parsimonious to interpret the human clinical and gene mutation data as indicating that loss of TCF4 gene regulatory function (transcriptional activation) is the basis for PTHS. Thus, screening candidate compounds would focus on reporter assays that will allow direct readout of TCF4 biochemical activity as manifest in altered gene transcription. The in vitro screens could be based, in general, on one of the two approaches. One approach would be to use purified TCF4 protein and screen its ability to bind normally to its regulatory element in DNA. The second approach would generate cell lines containing transfected DNA expressing a chemically tagged gene that would allow rapid quantitative assessment of the effect of a compound to block or augment TCF4-driven gene expression, for example, a green fluorescent protein gene driven by a TCF4-responsive promoter element.
An important and promising variation of this approach is to use inducible pluripotent stem cells derived directly from PTHS patients as the in vitro system for studying TCF4 control of gene transcription (Stephen Haggarty, personal communication). These inducible pluripotent stem cells, generated from fibroblasts from skin biopsies, provide an especially appealing model system for drug screening because they derive directly from a bona fide PTHS mutation, and are indeed human cells, the clinically relevant target. Using inducible pluripotent stem cell lines in vitro, pharmacological potentiators of TCF4 transcriptional activation or efficacy can be directly tested using human-derived material.
In general, four types of outcomes are possible for the chemical screens, using either biochemical activity or gene expression in cell lines as the readout. The vast majority of compounds will have no effect on TCF4 activity. However, those compounds that affect TCF4 activity will produce one of the three effects: (1) They could inhibit TCF4 activity, in which case they might be useful in modeling PTHS by mimicking the effect of the gene mutation. (2) They could directly activate TCF4, in which case they would be worthwhile testing in behavioral and physiological models of PTHS. (3). The most desirable outcome would be agents that augment TCF4 activity without directly activating the protein per se. This third type of compound is called a positive allosteric modulator, an agent which can enhance the normal physiological activation of TCF4 without triggering effects on TCF4 independent of another normal signal in the cell. Positive allosteric modulators are highly desirable potential therapeutic agents because of their selective actions and decreased likelihood of negative side effects.
Two broad types of chemical libraries are available to use as the sources of chemical compounds to test for effects on TCF4. First are large random libraries of miscellaneous compounds with a wide variety of structures. Second are libraries of chemicals that are already FDA-approved drugs that are currently in use in humans for other disorders. The most desirable approach is to first screen already-approved drugs—in that case if one gets a positive ‘hit’ it can proceed much more rapidly to use in human clinical trials. The second approach is to screen large random libraries—this has the benefit of increasing the likelihood of identifying an active compound and indeed is required in order to identify novel patentable compounds. However, as PTHS is an orphan disease with only a small number of affected individuals, the cost of this second approach is likely prohibitive. In both the cases, however, developing exactly the same screening methods is necessary in order to have a way to monitor effects of the compounds on TCF4 activity.
After the development of in vitro procedures that can be used to assess the biochemical function of TCF4, and the subsequent use of these biochemical assays to screen chemical libraries for compounds that can augment TCF4 function, candidate compounds can also be tested in the mouse lines described above to see whether they improve cognitive and neurophysiologialc function in the PTHS mouse models.
Using mouse models to screen for novel PTHS therapeutics
We are discussing the central hypothesis that TCF4 regulates the brain’s ability to store memories by actively regulating transcriptional activity during learning and memory formation and that disruption of this function leads to cognitive, behavioral and synaptic deficits in PTHS. Although testing this will initially focus on characterizing the behavioral and physiological effects of loss of TCF4 function in order to understand PTHS as described above, one additional scientific objective will be to use PTHS model mice to test novel potential routes of pharmacotherapy for PTHS. The compounds tested might arise from the in vitro screens as described above. However, one can also use a hypothesis-driven approach to choosing novel therapies that might be useful in PTHS patients. One such candidate type of compound is HDACi. Generally acting promoters of transcriptional activation such as HDACi are an appealing category of potential treatment for PTHS. HDACi, in general, act as ‘boosters’ of transcriptional activation and I hypothesize that this effect will compensate for the haplo-insufficiency of the TCF4 gene product, by augmenting the action of the remaining half-complement of TCF4 still present in PTHS patients.
TCF4 is a transcription factor and thus its function is to regulate gene expression. As described above, normally transcription factors lie dormant in the nucleus, subject to activation by signaling pathways upstream of them. Thus, TCF4 then binds to its appropriate DNA response elements found in the upstream regions of genes, a step necessary for transcription of downstream target genes. However, the efficacy of transcription factors for modulation of transcription also depends critically upon the recruitment and activation of a number of co-activators of transcription, including histone acetyl transferases (HATs) such as the CREB-binding protein. For transcription to occur, the native structure of chromatin must be modified, an effect mediated by post-translational modifications of histones, including acetylation, methylation and phosphorylation. Acetylation of the ɛ-amino group of lysine residues by HATs is necessary for recruiting transcriptionally competent complexes to the DNA to trigger gene expression. Thus, regulation of response element-dependent transcription by TCF4 likely depends upon the activity of HATs.
Furthermore, the Sweatt laboratory has also shown that dynamic regulation of histone modifications are involved in long-term memory formation.72 These considerations lead to our overall hypothesis that the mutation in TCF4 that underlies PTHS interferes with the mechanisms that regulate chromatin structure and gene transcription in areas of the brain important for memory and learning. This disregulation of these normal transcription-controlling mechanisms, we hypothesize, underlies aspects of memory and synaptic dysfunction in PTHS. With this in mind, it is desirable to test whether enhancing histone acetylation can rescue any plasticity and memory deficits observed in the PTHS model mice described above.
How can one enhance histone acetylation? The effects of HATs are reversed by HDACs and by this mechanism HDACs are also critical controllers of histone acetylation and transcriptional efficacy. There are 11 HDAC isoforms, and the specific isoforms involved in memory formation and HDACi-induced memory and LTP enhancement are not known. HDAC6 is the cytoplasmic ‘tubulin’ de-acetylase, but the other HDAC isoforms all affect histone acetylation. Although isoform-selective HDACi are being developed, these are not readily available at present and, in general, are not thoroughly characterized. Thus, at present, it is possible to use broad-spectrum HDACi, all of which have been shown to enhance memory formation in normal rodents, in studies of PTHS mice. These preclinical proof-of-principle studies can evaluate whether HDACi inhibitors can ameliorate the any memory and synaptic plasticity deficits exhibited by genetically engineered mouse models of PTHS. These are exciting studies that will provide an initial validation (or refutation) of the idea of regulation of chromatin structure as a ‘druggable’ target for novel PTHS therapeutics. The development of new valid targets for PTHS therapeutics is a pressing and a compelling societal and biomedical issue and thus must proceed quickly: one way to do this is to use currently available memory-enhancing agents (see Table 4).
Possible treatment through TCF4 replacement
Gene replacement therapy in the CNS is largely unachievable through contemporary methods, due to lack of effective and safe viral vectors for human use and concerns regarding mutational insertion of the replacement gene into the genome. In addition, overexpression of the therapeutic transgene is a real concern in the case of PTHS, because, as described above, transgenic mice overexpressing TCF4 in the CNS have learning deficits and potential schizophrenia-like phenotypes. A more subtle approach might be upregulation of the remaining endogenous normal copy of the TCF4 gene in PTHS patients, as this would presumably retain the normal cellular and subcellular TCF4 expression patterns in vivo. Also, the remaining normal copy would potentially be regulated by the endogenous promoters for generating splice variant isoforms, and so on. Although the endogenous regulatory mechanism are mysterious at this point, the work of the Timmusk group that was described above wherein they are identifying endogenous regulators of TCF4 expression and splicing in neurons is laying a valuable foundation in this regard. A final intriguing possibility in this conceptual area is the idea of regulating neuronal TCF4 through manipulating the endogenous activity of the miRNA miR-137. As has already been discussed, miR-137 has been shown to be a potent regulator of TCF4. Hypothetically one could decrease miR-137 levels through antisense oligonucleotide expression in the CNS and upregulate endogenous TCF4 levels, restoring function. However, important caveats apply to this approach, including the possibility of ‘off-target’ effects due to altered expression of other miR-137 targets, and the considerations listed above concerning potential effects of TCF4 overexpression.
Summary and future directions
PTHS is a rare human mental retardation syndrome associated with profound deficits in general learning and memory and an almost complete lack of language learning. The genetic basis of PTHS is clear—heterozygous deficiency of the TCF4 transcription factor. Thus, although the molecular neurobiology underlying PTHS has been essentially unexplored to date, transcriptional dysregulation in language- and memory-associated areas of the CNS is almost certainly a contributing basis for the cognitive defects in PTHS. However, because the disease is due to heterozygous mutation or deletion, half of the normal amount of TCF4 is still present in PTHS patients. This opens up an avenue for potential therapeutics. At the simplest level, restoring function in PTHS patients might be achieved either by epigenetically upregulating the remaining normal copy of the TCF4 gene or by using epigenetic therapy (for example, HDAC inhibition) to boost up the transcriptional efficiency of the remaining TCF4 in the CNS. Thus, understanding CNS epigenetic regulation in the TCF4-deficient CNS could have a profound effect on our understanding of PTHS and in identifying novel therapeutics. There is an additional real-life consideration that also motivates studying PTHS in terms of therapeutic development—as an extremely debilitating orphan disease, human clinical studies of novel therapeutics for PTHS would be candidates for expedited approval by IRBs and the FDA. Finally, in the basic science domain, studying PTHS will yield new insights into the roles and mechanisms by which the TCF4 transcription factor regulates normal learning, memory and language acquisition.
Even though PTHS is a rare disorder, understanding the mechanistic basis for the disease can have quite a broad-ranging impact both scientifically and in terms of improving medical treatment. PTHS patients manifest both learning disabilities and autistic behaviors, so discoveries concerning the mechanistic basis of PTHS will have direct relevance in a variety of intellectual disabilities, memory disorders and ASDs. Moreover, because the genetic basis of PTHS is already precisely known, research in PTHS may rapidly advance in understanding the underlying molecular neurobiology of this disorder. This advancement for a single-gene disorder might proceed much more quickly than would be achievable by studying the more complex learning, memory and autism disorders of unknown etiology. Yet, discoveries of how PTHS occurs would have direct relevance to these many other diseases. In essence, knowledge gained from studying the orphan disease PTHS would potentially allow broad understanding of a wide variety of learning and memory disabilities that afflict many families.
PTHS is an ASD and an orphan disease with likely only a few hundred patients in the United States. At present, there are very few molecular neurobiologists in the US working on this disorder. I feel there is an ethical and social imperative for the US basic neuroscience community to begin to investigate the molecular neurobiology of PTHS,and lay a cornerstone of research that will allow the hope of the development of a treatment for these patients in the future. In addition, an ancillary goal is needed to develop a cadre of basic and clinical researchers who are working on this syndrome, in order to have at least a minimal critical mass of clinicians and scientists who are knowledgeable in this area.
Accession codes
References
Ardinger HH, Welsh HI, Saunders CJ . Pitt-Hopkins Syndrome. In: Pagon RA, Bird TD, Dolan CR, Stephens K, Adam MP (eds). GeneReviews [Internet]. University of Washington: Seattle, WA, USA, 1993-2012. Aug 30 2012.
Whalen S, Héron D, Gaillon T, Moldovan O, Rossi M, Devillard F et al. Novel comprehensive diagnostic strategy in Pitt-Hopkins syndrome: clinical score and further delineation of the TCF4 mutational spectrum. Hum Mutat 2012; 33: 64–72.
Brockschmidt A, Todt U, Ryu S, Hoischen A, Landwehr C, Birnbaum S et al. Severe mental retardation with breathing abnormalities (Pitt-Hopkins syndrome) is caused by haploinsufficiency of the neuronal bHLH transcription factor TCF4. Hum Mol Genet 2007; 16: 1488–1494.
Zweier C, Peippo MM, Hoyer J, Sousa S, Bottani A, Clayton-Smith J et al. Haploinsufficiency of TCF4 causes syndromal mental retardation with intermittent hyperventilation (Pitt-Hopkins syndrome). Am J Hum Genet 2007; 80: 994–1001.
Amiel J, Rio M, de Pontual L, Redon R, Malan V, Boddaert N et al. Mutations in TCF4, encoding a class I basic helix-loop-helix transcription factor, are responsible for Pitt-Hopkins syndrome, a severe epileptic encephalopathy associated with autonomic dysfunction. Am J Hum Genet 2007; 80: 988–993.
Navarrete K, Pedroso I, De Jong S, Stefansson H, Steinberg S, Stefansson K et al. TCF4 (e2-2; ITF2): A schizophrenia-associated gene with pleiotropic effects on human disease. Am J Med Genet B Neuropsychiatr Genet 2013; 162: 1–16.
Pitt D, Hopkins I . A syndrome of mental retardation, wide mouth and intermittent overbreathing. Aust Paediatr J 1978; 14: 182–184.
Hasi M, Soileau B, Sebold C, Hill A, Hale DE, O'Donnell L et al. The role of the TCF4 gene in the phenotype of individuals with 18q segmental deletions. Hum Genet 2011; 130: 777–787.
Smith R, Taylor JP . Dissection and imaging of active zones in the Drosophila neuromuscular junction. J Vis Exp 2011 JoVE video 2676.
Brockschmidt A, Filippi A, Charbel Issa P, Nelles M, Urbach H, Eter N et al. Neurologic and ocular phenotype in Pitt-Hopkins syndrome and a zebrafish model. Hum Genet 2011; 130: 645–655.
Stavropoulos DJ, MacGregor DL, Yoon G . Mosaic microdeletion 18q21 as a cause of mental retardation. Eur J Med Genet 2010; 53: 396–399.
Blake DJ, Forrest M, Chapman RM, Tinsley CL, O'Donovan MC, Owen MJ . TCF4, schizophrenia, and Pitt-Hopkins syndrome. Schizophr Bull 2010; 36: 443–447.
Taddeucci G, Bonuccelli A, Mantellassi I, Orsini A, Tarantino E . Pitt-Hopkins syndrome: report of a case with a TCF4 gene mutation. Ital J Pediatr 2010; 36: 12.
Takano K, Lyons M, Moyes C, Jones J, Schwartz CE . Two percent of patients suspected of having Angelman syndrome have TCF4 mutations. Clin Genet 2010; 78: 282–288.
Rosenfeld JA, Leppig K, Ballif BC, Thiese H, Erdie-Lalena C, Bawle E et al. Genotype-phenotype analysis of TCF4 mutations causing Pitt-Hopkins syndrome shows increased seizure activity with missense mutations. Genet Med 2009; 11: 797–805.
Zweier C, de Jong EK, Zweier M, Orrico A, Ousager LB, Collins AL et al. CNTNAP2 and NRXN1 are mutated in autosomal-recessive Pitt-Hopkins-like mental retardation and determine the level of a common synaptic protein in Drosophila. Am J Hum Genet 2009; 85: 655–666.
Kato Z, Morimoto W, Kimura T, Matsushima A, Kondo N . Interstitial deletion of 18q: comparative genomic hybridization array analysis of 46, XX,del(18)(q21.2.q21.33). Birth Defects Res A Clin Mol Teratol 2010; 88: 132–135.
Fukumura S, Tachi N . Case of Pitt-Hopkins syndrome. No To Hattatsu 2009; 41: 365–367.
de Pontual L, Mathieu Y, Golzio C, Rio M, Malan V, Boddaert N et al. Mutational, functional, and expression studies of the TCF4 gene in Pitt-Hopkins syndrome. Hum Mutat 2009; 30: 669–676.
Cisse B, Caton ML, Lehner M, Maeda T, Scheu S, Locksley R et al. Transcription factor E2-2 is an essential and specific regulator of plasmacytoid dendritic cell development. Cell 2008; 135: 37–48.
Giurgea I, Missirian C, Cacciagli P, Whalen S, Fredriksen T, Gaillon T et al. TCF4 deletions in Pitt-Hopkins syndrome. Hum Mutat 2008; 29: E242–E251.
Zweier C, Sticht H, Bijlsma EK, Clayton-Smith J, Boonen SE, Fryer A et al. Further delineation of Pitt-Hopkins syndrome: phenotypic and genotypic description of 16 novel patients. J Med Genet 2008; 45: 738–744.
Kim SK, Jang HR, Kim JH, Kim M, Noh SM, Song KS et al. CpG methylation in exon 1 of transcription factor 4 increases with age in normal gastric mucosa and is associated with gene silencing in intestinal-type gastric cancers. Carcinogenesis 2008; 29: 1623–1631.
Kalscheuer VM, Feenstra I, Van Ravenswaaij-Arts CM, Smeets DF, Menzel C, Ullmann R et al. Disruption of the TCF4 gene in a girl with mental retardation but without the classical Pitt-Hopkins syndrome. Am J Med Genet A 2008; 146A: 2053–2059.
Ouvrier R . Hyperventilation and the Pitt-Hopkins syndrome. Dev Med Child Neurol 2008; 50: 481.
Andrieux J, Lepretre F, Cuisset JM, Goldenberg A, Delobel B, Manouvrier-Hanu S et al. Deletion 18q21.2q21.32 involving TCF4 in a boy diagnosed by CGH-array. Eur J Med Genet 2008; 51: 172–177.
Peippo MM, Simola KO, Valanne LK, Larsen AT, Kähkönen M, Auranen MP et al. Pitt-Hopkins syndrome in two patients and further definition of the phenotype. Clin Dysmorphol 2006; 15: 47–54.
Orrico A, Galli L, Zappella M, Lam CW, Bonifacio S, Torricelli F, Hayek G . Possible case of Pitt-Hopkins syndrome in sibs. Am J Med Genet 2001; 103: 157–159.
Van Balkom ID, Quartel S, Hennekam RC . Mental retardation, ‘coarse’ face, and hyperbreathing: confirmation of the Pitt-Hopkins syndrome. Am J Med Genet 1998; 75: 273–276.
Peippo M, Ignatius J . Pitt-Hopkins syndrome. Mol Syndromol 2012; 2: 171–180.
Pitt-Hopkins Syndrome Support Group http://groups.google.com/group/pitt-hopkins/about?pli=1.
Pitt-Hopkins Syndrome International Network http://pitthopkins.org/.
Takano K, Tan WH, Irons MB, Jones JR, Schwartz CE . Pitt-Hopkins syndrome should be in the differential diagnosis for males presenting with an ATR-X phenotype. Clin Genet 2011; 80: 600–601.
Marangi G, Ricciardi S, Orteschi D, Lattante S, Murdolo M, Dallapiccola B et al. The Pitt-Hopkins syndrome: report of 16 new patients and clinical diagnostic criteria. Am J Med Genet A 2011; 155A: 1536–1545.
Marangi G, Ricciardi S, Orteschi D, Tenconi R, Monica MD, Scarano G et al. Proposal of a clinical score for the molecular test for Pitt-Hopkins syndrome. Am J Med Genet A 2012; 158A: 1604–1611.
Lehalle D, Williams C, Siu VM, Clayton-Smith J . Fetal pads as a clue to the diagnosis of Pitt-Hopkins syndrome. Am J Med Genet A 2011; 155A: 1685–1689.
Ghosh PS, Friedman NR, Ghosh D . Pitt-Hopkins byndrome in a boy with Charcot Marie Tooth Disease type 1A: a rare co-occurrence of 2 genetic disorders. J Child Neurol 2012; 27: 1602–1606.
Verhulst SL, De Dooy J, Ramet J, Bockaert N, Van Coster R, Ceulemans B et al. Acetazolamide for severe apnea in Pitt-Hopkins syndrome. Am J Med Genet A. 2012; 158A: 932–934.
Van Balkom ID, Vuijk PJ, Franssens M, Hoek HW, Hennekam RC . Development, cognition, and behaviour in Pitt-Hopkins syndrome. Dev Med Child Neurol 2012; 54: 925–931.
Maini I, Cantalupo G, Turco EC, Paolis FD, Magnani C, Parrino L et al. Clinical and polygraphic improvement of breathing abnormalities after valproate in a case of Pitt-Hopkins syndrome. J Child Neurol 2012; 27: 1585–1588.
Takenouchi T, Yagihashi T, Tsuchiya H, Torii C, Hayashi K, Kosaki R et al. Tissue-limited ring chromosome 18 mosaicism as a cause of Pitt-Hopkins syndrome. Am J Med Genet A. 2012; 158A: 2621–2623.
Armani R, Archer H, Clarke A, Vasudevan P, Zweier C, Ho G et al. Transcription factor 4 and myocyte enhancer factor 2C mutations are not common causes of Rett syndrome. Am J Med Genet A 2012; 158A: 713–719.
Steinbusch C, van Roozendaal K, Tserpelis D, Smeets E, Kranenburg-de Koning T, de Waal K et al. Somatic mosaicism in a mother of two children with Pitt-Hopkins syndrome. Clin Genet 2013; 83: 73–77.
Forrest M, Chapman RM, Doyle AM, Tinsley CL, Waite A, Blake DJ . Functional analysis of TCF4 missense mutations that cause Pitt-Hopkins syndrome. Hum Mutat 2012; 33: 1676–1686.
Gregor A, Albrecht B, Bader I, Bijlsma EK, Ekici AB, Engels H et al. Expanding the clinical spectrum associated with defects in CNTNAP2 and NRXN1. BMC Med Genet 2011; 12: 106.
Harrison V, Connell L, Hayesmoore J, McParland J, Pike MG, Blaire E . Compound heterozygous deletion of NRXN1 causing severe developmental delay with early onset epilepsy in two sisters. Am J Med Genet A 2011; 155A: 2826–2831.
Alarcón M, Abrahams BS, Stone JL, Duvall JA, Perederiy JV, Bomar JM et al. Linkage, association, and gene-expression analysesidentify CNTNAP2 as an autism-susceptibility gene. Am J Hum Genet 2008; 82: 150–159.
Arking DE, Cutler DJ, Brune CW, Teslovich TM, West K, Ikeda M et al. A common genetic variant in the neurexin superfamily member CNTNAP2 increases familial risk of autism. Am J Hum Genet 2008; 82: 160–164.
Bakkaloglu B, O'Roak BJ, Louvi A, Gupta AR, Abelson JF, Morgan TM et al. Molecular cytogenetic analysis and resequencing of contactin associated protein-like 2 in autism spectrum disorders. Am J Hum Genet 2008; 82: 165–173.
Vernes SC . A functional genetic link between distinct developmental language disorders. N Engl J Med 2008; 359: 2337–2345.
Cohen J . First genetic clue to common speech disorder. ScienceNOW Daily 2008 November 5. http://news.sciencemag.org/sciencenow/2008/11/05-04.html?ref=hp.
Sepp M, Pruunsild P, Timmusk T . Pitt-Hopkins syndrome-associated mutations in TCF4 lead to variable impairment of the transcription factor function ranging from hypomorphic to dominant-negative effects. Hum Mol Genet 2012; 21: 2873–2888.
Murre C . Helix-loop-helix proteins and lymphocyte development. Nat Immunol 2005; 6: 1079–1086.
Murre C, McCaw PS, Vaessin H, Caudy M, Jan LY, Jan YN et al. Interactions between heterologous helix-loop-helix proteins generate complexes that bind specifically to a common DNA sequence. Cell 1989; 58: 537–544.
Sepp M, Kannike K, Eesmaa A, Urb M, Timmusk T . Functional diversity of human basic helix-loop-helix transcription factor TCF4 isoforms generated by alternative 5' exon usage and splicing. PLoS One 2011; 6: e22138.
Saarikettu J . Calcium/calmodulin inhibition of transcriptional activity of E-proteins by prevention of their binding to DNA. J Biol Chem 2004; 279: 41004–41011.
Fu H, Cai J, Clevers H, Fast E, Gray S, Greenberg R et al. A genome-wide screen for spatially restricted expression patterns identifies transcription factors that regulate glial development. J Neurosci 2009; 29: 11399–11408.
Ripke S, Sanders AR, Kendler KS, Levinson DF, Sklar P, Holmans PA et al. Genome-wide association study identifies five new schizophrenia loci. Nat Genet 2011; 43: 969–976.
Kwon E, Wang W, Tsai LH . Validation of schizophrenia-associated genes CSMD1, C10orf26, CACNA1C and TCF4 as miR-137 targets. Mol Psychiatry 2013; 18: 11–12.
Kim AH, Parker EK, Williamson V, McMichael GO, Fanous AH, Vladimirov VI . Experimental validation of candidate schizophrenia gene ZNF804A as target for hsa-miR-137. Schizophr Res 2012; 141: 60–64.
Weeber EJ, Sweatt JD . Molecular neurobiology of human cognition. Neuron 2002; 33: 845–848.
Skarnes WC, Rosen B, West AP, Koutsourakis M, Bushell W, Iyer V et al. A conditional knockout resource for the genome-wide study of mouse gene function. Nature 2011; 474: 337–342.
Bergqvist I, Eriksson M, Saarikettu J, Eriksson B, Corneliussen B, Grundström T et al. The basic helix-loop-helix transcription factor E2-2 is involved in T lymphocyte development. Eur J Immunol 2000; 30: 2857–2863.
Brzózka MM, Radyushkin K, Wichert SP, Ehrenreich H, Rossner MJ . Cognitive and sensorimotor gating impairments in transgenic mice overexpressing the schizophrenia susceptibility gene Tcf4 in the brain. Biol Psychiatry 2010; 68: 33–40.
Chen RZ, Akbarian S, Tudor M, Jaenisch R . Deficiency of methyl-CpG binding protein-2 in CNS neurons results in a Rett-like phenotype in mice. Nat Genet 2001; 27: 327–331.
Collins AL, Levenson JM, Vilaythong AP, Richman R, Armstrong DL, Noebels JL et al. Mild overexpression of MeCP2 causes a progressive neurological disorder in mice. Hum Mol Genet 2004; 13: 2679–2689.
Guy J, Hendrich B, Holmes M, Martin JE, Bird A . A mouse Mecp2-null mutation causes neurological symptoms that mimic Rett syndrome. Nat Genet 2001; 27: 322–326.
Moretti P, Levenson JM, Battaglia F, Atkinson R, Teague R, Antalffy B et al. Learning and memory and synaptic plasticity are impaired in a mouse model of Rett syndrome. J Neurosci 2006; 26: 319–327.
Pelka GJ, Watson CM, Radziewic T, Hayward M, Lahooti H, Christodoulou J et al. Mecp2 deficiency is associated with learning and cognitive deficits and altered gene activity in the hippocampal region of mice. Brain 2006; 129: 887–898.
Shahbazian M, Young J, Yuva-Paylor L, Spencer C, Antalffy B, Noebels J et al. Mice with truncated MeCP2 recapitulate many Rett syndrome features and display hyperacetylation of histone H3. Neuron 2002; 35: 243–254.
Shalin SC, Hernandez CM, Dougherty MK, Morrison DK, Sweatt JD . Kinase suppressor of Ras1 compartmentalizes hippocampal signal transduction and subserves synaptic plasticity and memory formation. Neuron 2006; 50: 765–779.
Sweatt JD . Experience-dependent epigenetic modifications in the central nervous system. Biol Psychiatry 2009; 65: 191–197.
Acknowledgements
I thank Andrew Kennedy, Audrey Davidow, Theresa Pauca, Sue Routledge and the entire PTHS support group for innumerable helpful discussions. The work in the author’s lab is supported by the PTHS Foundation, the Simons Foundation, NIMH Grant MH 57014, Civitan International and Evelyn F. McKnight Brain Research Foundation.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivs 3.0 Unported License. To view a copy of this license, visit http://creativecommons.org/licenses/by-nc-nd/3.0/
About this article
Cite this article
Sweatt, J. Pitt–Hopkins Syndrome: intellectual disability due to loss of TCF4-regulated gene transcription. Exp Mol Med 45, e21 (2013). https://doi.org/10.1038/emm.2013.32
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/emm.2013.32
Keywords
This article is cited by
-
Evaluation of Nav1.8 as a therapeutic target for Pitt Hopkins Syndrome
Molecular Psychiatry (2023)
-
Transcription factor 4 controls positioning of cortical projection neurons through regulation of cell adhesion
Molecular Psychiatry (2021)
-
Disorders of the enteric nervous system — a holistic view
Nature Reviews Gastroenterology & Hepatology (2021)
-
Survival control of oligodendrocyte progenitor cells requires the transcription factor 4 during olfactory bulb development
Cell Death & Disease (2021)
-
Transcription factor 4 and its association with psychiatric disorders
Translational Psychiatry (2021)
