
Abstract
Melanoma inhibiting activity/cartilage-derived retinoic acid-sensitive protein (MIA/CD-RAP) is a small soluble protein secreted from malignant melanoma cells and from chondrocytes. Recently, we revealed that MIA/CD-RAP can modulate bone morphogenetic protein (BMP)2-induced osteogenic differentiation into a chondrogenic direction. In the current study we aimed to find the molecular details of this MIA/CD-RAP function. Direct influence of MIA on BMP2 by protein-protein-interaction or modulating SMAD signaling was ruled out experimentally. Instead, we revealed inhibition of ERK signaling by MIA/CD-RAP. This inhibition is regulated via binding of MIA/CD-RAP to integrin α5 and abolishing its activity. Active ERK signaling is known to block chondrogenic differentiation and we revealed induction of aggrecan expression in chondrocytes by treatment with MIA/CD-RAP or PD098059, an ERK inhibitor. In in vivo models we could support the role of MIA/CD-RAP in influencing osteogenic differentiation negatively. Further, MIA/CD-RAP-deficient mice revealed an enhanced calcified cartilage layer of the articular cartilage of the knee joint and disordered arrangement of chondrocytes. Taken together, our data indicate that MIA/CD-RAP stabilizes cartilage differentiation and inhibits differentiation into bone potentially by regulating signaling processes during differentiation.
Similar content being viewed by others

Introduction
Melanoma inhibiting activity/cartilage-derived retinoic acid-sensitive protein (MIA/CD-RAP) was cloned as a secreted protein from human melanoma cell lines (Blesch et al., 1994) and independently by differential display comparing differentiated with dedifferentiated chondrocytes (Dietz and Sandell, 1996). Several studies indicate an important role of MIA/CD-RAP in tumor progression and metastasis of melanoma as MIA/CD-RAP mediates detachment of melanoma cells from extracellular matrix molecules such as fibronectin by inhibition of integrin activity where specifically integrin alpha 5 and alpha 4 beta 1 were shown to be affected by melanoma inhibiting activity (MIA) (Bosserhoff et al., 2003; Bauer et al., 2006; Stoll et al., 2006).
The three-dimensional structure of MIA/CD-RAP has been solved by multi-dimensional NMR (Stoll et al., 2001) and independently by x-ray crystallography (Lougheed et al., 2001) and indicated that MIA/CD-RAP defines a novel family of secreted proteins, which adopt an SH3 domain-like fold in solution (Stoll and Bosserhoff, 2008).
In non-neoplastic tissues MIA/CD-RAP expression is activated from the beginning of chondrogenesis throughout cartilage development (Dietz and Sandell 1996; Bosserhoff et al., 1997; Moser et al., 2002) and in vitro it is a specific marker for chondrocytic differentiation (Bosserhoff et al., 2003). Based on its highly restricted activity the MIA/CD-RAP promoter was used to study transcriptional mechanisms mediating chondrocyte differentiation. Expression of a LacZ-reporter under control of the 2251 bp MIA/CD-RAP promoter was exclusively observed in cartilage (Xie et al., 2000). These data suggested that MIA/CD-RAP may be functionally important for cartilage differentiation.
To study the consequences of MIA/CD-RAP deficiency in vivo we inactivated the gene by targeted germline mutation (Moser et al., 2002). These data indicate that MIA/CD-RAP function is required for the highly ordered fibrillar ultrastructure of cartilage.
In a recent study we revealed that MIA/CD-RAP can influence cartilage differentiation (Tscheudschilsuren et al., 2006). MIA/CD-RAP was not able to induce chondrocytic differentiation of human mesenchymal stem cells on its own but induced differentiation mediated by TGF-beta and reverted osteogenic differentiation by BMP2 into more chondrogenic differentiation.
We, therefore, aimed to analyze the molecular function of MIA/CD-RAP in cartilage differentiation using in vitro and in vivo models.
Results
As recent studies revealed an influence of MIA/CD-RAP on bone morphogenetic protein (BMP)-induced differentiation of mesenchymal stem cells showing inhibition of osteogenic differentiation and induction of chondrogenic differentiation, we aimed to analyze this mechanism in detail and to reveal whether MIA is important to stabilize chondrocytic differentiation.
Mechanisms of interaction with BMP signaling
Modulation of BMP signaling by MIA could be due to either direct binding of MIA to BMPs or by influencing the signaling capacity. Co-immunoprecipitation (Figure 1A) and ELISA (data not shown) clearly ruled out direct interaction of MIA/CD-RAP and BMP2. To analyze whether MIA/CD-RAP regulates BMP-induced SMAD signaling, phosphorylation of SMAD1/5 was determined by western blotting lysates derived from chondrocytes either treated with BMP2 alone or with BMP/MIA/CD-RAP (Figure 1B). No influence on SMAD1/5 regulation was observed by MIA/CD-RAP treatment.

No direct regulation of BMP signaling by MIA. (A) For co-immunoprecipitation biotinylated MIA was incubated with recombinant BMP2, purified via streptavidin-coated beads, separated on a 12% SDS-PAGE followed by blotting and detection with an anti-BMP2 antibody. No interaction of these two molecules was observed (lane: BMP2 + MIA). As a negative control BMP2 alone was used in the assays (BMP2); as a positive control BMP2 was separated on the gel, blotted and detected by the antibody (BMP2 control). (B) Induction of SMAD1/5 phosphorylation was determined by western blot. Treatment of chondrocytes with BMP2 (200 ng/ml) resulted in increase P-SMAD 1/5 detection (+BMP2). However, addition of MIA (100 ng/ml) did not change p-SMAD1/5 levels (+BMP2 + MIA).
In addition to regulation of SMAD signaling also ERK activation is induced by BMPs. To analyze the relevance of SMAD and ERK signaling for chondrogenic and osteogenic differentiation, human mesenchymal stem cells were treated with BMP2 to induce osteogenic differentiation. This process was either modulated by treatment with chordin, a molecule known as BMP inhibitor, or dorsomorphin, which was recently shown to inhibit specifically BMP-induced SMAD signaling (Yu et al., 2008). Dorsomorphin and chordin inhibited induction of both osteoblast specific markers osteocalcin and osteopontin significantly (Figures 2A and 2B). In contrast, only chordin, inhibiting BMP-induced SMAD and ERK signaling resulted in significant induction of chondrogenic markers like Sox9 and aggrecan. Dorsomorphin treatment, only inhibiting SMAD activation by BMP2, resulted in unchanged expression of these two genes (Figures 2C and 2D). Interestingly, studies of other groups revealed that ERK activity is a negative regulator of cartilage-specific gene expression (Bobick et al., 2004).

Influence of SMAD and ERK activity on HMSC differentiation. HMSCs were treated with BMP2 (200 ng/ml) to induce osteogenic differentiation. Cells were co-treated with either dorsomorphin (2 µM) or chordin (2 µg/ml), respectively. qRT-PCR revealed reduced expression of osteopontin (A) and osteocalcin (B) mRNA after treatment. Expression of aggrecan (C) and SOX9 (D) stayed unchanged after dorsomorphin treatment whereas chordin led to induced expression of these two genes. The experiments were repeated 3 times using cells from different donors. (*: P < 0.05 compared to BMP2 treatment)
Influence of MIA on chondrocytes
In a recent study we revealed that MIA/CD-RAP can inhibit ERK activity via negative regulation of integrin alpha 5 activity (Bauer et al., 2006). We confirmed strong expression of integrin alpha 5 in chondrocytes by RT-PCR and demonstrated MIA/CD-RAP binding to integrin alpha 5 of chondrocytes by co-immunoprecipitation (Figures 3A and 3B). To analyze MIA/CD-RAP effects on ERK signaling in chondrocytes, dedifferentiated chondrocytes in cell culture were treated with MIA/CD-RAP (100 ng/ml) for 30 min and regulation of ERK activity was analyzed by western blot (Figure 3C). Here, significant reduction of ERK activity was observed after MIA/CD-RAP treatment. Treatment with the ERK inhibitor PD098059 resulted in the expected reduction of ERK activity whereas the antibody JBS5 resulted in stimulation of ERK activity as already published (Forsyth et al., 2002). The ERK-stimulating effects of the antibody could be significantly reduced by PD treatment but not by MIA/CD-RAP.

MIA/CD-RAP binds to integrin alpha 5 on chondrocytes and modulates ERK activity and chondrocytic differentiation in vitro. (A) Integrin alpha 5 expression in chondrocytes (2 donors) was confirmed by RT-PCR. (B) Interaction of integrin alpha 5 and MIA/CD-RAP in chondrocytes was revealed by co-immunoprecipitation. Here, lysate of chondrocytes was incubated with or without biotinylated MIA/CD-RAP and precipitation was performed on streptavidin beads. Precipitation was followed by a western blot to detect integrin alpha 5. (C) Regulation of ERK signaling in dedifferentiated chondrocytes treated with MIA/CD-RAP (200 ng/ml), PD098059 (20 µM), JSB5 (5 µg/ml) or combinations was analyzed by western blot using an anti phospho-ERK1/2 antibody. The experiments were repeated 3 times using cells from different donors. (D, E) Dedifferentiated chondrocytes were treated with MIA/CD-RAP, PD098059 (ERK-inhibitor) and the activating integrin alpha 5 antibody JBS5 either alone or in combination, followed by RNA extraction and quantitative RT-PCR for aggrecan, a cartilage specific molecule (D) and osteocalcin (E), a osteoblast specific molecule.
MIA/CD-RAP influence on ERK activity in chondrocytes was determined in cell culture in more detail. Dedifferentiated chondrocytes were treated with MIA/CD-RAP, PD098059 (ERK-inhibitor) and the activating integrin alpha 5 antibody JBS5 either alone or in combination, followed by RNA extraction and quantitative RT-PCR for aggrecan, a cartilage specific molecule (Figure 3D). MIA was shown to induce aggrecan expression as well as the ERK inhibitor whereas the antibody JBS5 inhibited aggrecan expression, potentially by further activating ERK (Forsyth et al., 2002). Effects of PD098059 could not be reversed by JBS5 effects as PD098059 acts downstream directly inhibiting ERK activity. MIA effects were completely abolished by an excess of JBS5 as both molecules directly affect integrin alpha 5. In addition, we analysed osteocalcin as marker for osteogenic differentiation. Here, MIA was revealed to inhibit osteocalcin expression while the antibody JBS5 slightly induced expression of the marker. Again MIA effects were compensated by an excess of JSB5 (Figure 3E). We, therefore, speculate that MIA/CD-RAP modulates chondrogenic differentiation via MIA/CD-RAP-regulated ERK-signaling.
Effects of loss of MIA on bone formation in vivo
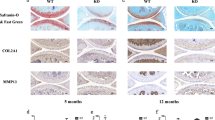
Following our results we hypothesized that MIA/CD-RAP modulates the balance between cartilage and bone formation. To get more insight we evaluated the height of the calcified zone of joint cartilage in tibia and femur of wild type and MIA/CD-RAP deficient at the age of 12 weeks (Figure 4A). The height of the calcified cartilage expressed as percentage of the overall cartilage height was significantly enhanced in MIA/CD-RAP deficient mice.

Effects of MIA/CD-RAP on the structure of articular cartilage in vivo. (A) In 12 weeks old mice, either wild type or MIA/CD-RAP-deficient, the height of the calcified cartilage of the femoral and tibia was measured in H&E sections. The height of the calcified cartilage expressed as percentage of the overall cartilage height was significantly enhanced in MIA/CD-RAP deficient mice. (B, C) The chondrocytes of layers 2 and 3 showed disordered cellular arrangement. In wild type mice chondrocytes were preferentially grouped in columns, whereas in MIA/CD-RAP deficient mice the chondrocytes were preferentially arranged in clusters. (C) H&E section of tibial cartilage of the knee joint of wild type (left micrograph) and MIA-deficient mice (right micrograph). The tidemark is indicated by an arrow, chondrocytes arranged in columns are indicated by arrowheads and chondrocytes arranged in clusters are indicated by closed circles (original magnification ×40).
The overall height of cartilage did not differ significantly between wild type and MIA/CD-RAP deficient mice (data not shown). Furthermore, the chondrocytes of layers 2 and 3 showed disordered cellular arrangement (Figure 4B). In wild type mice chondrocytes were preferentially grouped in columns, whereas in MIA/CD-RAP deficient mice the chondrocytes were preferentially arranged in clusters (Figure 4C).
Discussion
Until today the molecular function of MIA/CD-RAP in cartilage and bone formation has not been analyzed in detail. The data generated in MIA/CD-RAP-deficient mice indicate that MIA/CD-RAP function is required for the highly ordered fibrillar ultrastructure of cartilage (Moser et al., 2002). In previous studies, we presented evidence that MIA/CD-RAP can modulate BMP2 induced osteogenic differentiation into chondrogenic differentiation (Tscheudschilsuren et al., 2006). To further focus on this role of MIA/CD-RAP we aimed to understand the interplay between BMP2 and MIA/CD-RAP and signaling pathways controlled by MIA/CD-RAP.
We could demonstrate that there is no physical interaction of MIA and BMP2 and no interference of MIA with the SMAD-pathway in chondrocytes. Instead MIA seems to modulate the osteogenic effects of BMP2 in human mesenchymal stem cells via inhibition of ERK signaling. BMP2 induces osteogenic differentiation of human mesenchymal stem cells via SMAD and ERK signaling (Lou et al., 2000). Expression of the osteogenic markers osteopontin and osteocalcin is upregulated via Smads and can be inhibited by the BMP inhibitor chordin and the Smad inhibitor dorsomorphin (Figures 2A and 2B). The activation of the ERK-pathway by BMP2 suppresses the expression of the chondrogenic markers aggrecan and SOX9 as demonstrated by complete inhibition of BMP-signaling including ERK as opposed to selective inhibition of Smad signaling which had no effect on the expression of chondrogenic markers (Figures 2C and 2D). Inhibition of ERK signaling was already previously described to force chondrogenic differentiation (Forsyth et al., 2002) as confirmed by our in vitro experiments showing increased expression of aggrecan after inhibition of the ERK-pathway in chondrocytes (Figure 4).
Specific inhibition of integrin activity of integrins alpha 5 and alpha 4 beta 1 by MIA was reported in melanoma cells (Bosserhoff et al., 2003; Bauer et al., 2006; Stoll et al., 2006). Furthermore, ERK-signaling downstream of integrins has been described in several studies (summarized in Yee et al., 2008). Therefore, we hypothesized that MIA might inhibit BMP2 effects by interference with the ERK-pathway via integrin alpha5. Data gained from in vitro experiments in chondrocytes suggest that MIA/CD-RAP can inhibit ERK signaling via inhibition of integrin alpha 5 (Figure 3C) most likely by physical interaction (Figure 3B). An activating antibody of integrin alpha 5 was shown to overcome the effect of MIA/CD-RAP in in vitro experiments supporting our hypothesis (Figure 3C). The presented effects of MIA on the process of BMP2-induced osteogenic differentiation may be interpreted as retardation of osteogenesis or stabilization of a chondrogenic phenotype. To support this hypothesis we examined the knee joint cartilage of MIA/CD-RAP-deficient mice closely. MIA/CD-RAP-deficient mice displayed subtle differences compared to wild type mice concerning two features: (1) the layer of calcified cartilage was significantly enlarged and (Figure 4A) (2) the spatial arrangement of chondrocytes was slightly disturbed (Figure 4B). The layer of calcified cartilage separates the hyaline articular cartilage from the subchondral bone (Revell et al., 1990; Lane et al., 1977; Hughes et al., 2005). There is a line of calcification in tissue sections between calcified and hyaline cartilage known as the tidemark (Bullough and Jagannath, 1983). The tidemark is a metabolically active region involved in mineralization of the overlying of non-calcified hyaline cartilage (Revell et al., 1990). Ossification depletes the calcified cartilage at the cartilage/bone interface and the tidemark has been thought of as a calcification front advancing towards the non-calcified cartilage. Active though slow remodeling takes place at the articular cartilage/subchondral bone interface in adult humans, rabbits, rats and mice (Revell et al., 1990; Sriram et al., 2009; Tomiya et al., 2009). This process of transformation from hyaline cartilage to calcified cartilage and subsequently to bone is commonly viewed as a slow form of endochondral ossification (Revell et al., 1990; Lane et al., 1977; Tomiya et al., 2009). This view is supported by the fact that expression of molecules typically involved in endochondral ossification such as for instance type X collagen are expressed in the tidemark of mice, rabbits and humans (Gannon et al., 1991; Hoyland et al., 1991; Kirsch et al., 1991; von der Mark et al., 1992; Walker et al., 1995). The enlargement of the zone of calcified cartilage in MIA/CD-RAP-deficient mice indicates that MIA might be involved in the correct timing and sequence of transformation of cartilage to bone during the process of endochondral ossification of joint cartilage. It can be speculated that MIA deposited in the extracellular cartilage matrix may bind to integrin alpha 5 on chondrocytes and preserve the chondrocytic phenotype even under the influence of BMPs before ossification finally takes place at a slow rate. The disturbed cellular arrangement of chondrocytes in MIA/CD-RAP-deficient mice displaying higher numbers of clusters and fewer columns in layers 2 and 3 can also be interpreted as sign of an altered process of endochondral ossification possibly disturbing the sequence of cellular movements and arrangements during ossification. So MIA may be involved in the regulation and retardation of bone formation during remodeling of joints.
More general speculations may be raised from the results of this study whether MIA/CD-RAP is involved in the process of the preservation of cartilage in the body. It seems as MIA/CD-RAP has the ability to reduce the effect of molecules leading to osteogenic differentiation and by this stabilizes the chondrogenic status. Future studies have to aim at the identification of redundancy factors which should be found over expressed in the MIA/CD-RAP-deficient mice.
Methods
Cell isolation and cell culture
Human primary chondrocytes derived from articular cartilage (CellSystems, St. Katharinen, Germany) were cultivated in DMEM (PAA, Pasching, Austria) supplemented with penicillin (100 U/ml), streptomycine (10 µg/ml) (both Sigma, Deisenhofen, Germany), and 10% fetal calf serum (Pan Biotech GmbH, Aidenbach, Germany) under a humidified atmosphere of 5% CO2 at 37℃ and splitted 1:2 at 80% confluency. As chondrocytes spontaneously de-differentiate in cell culture, redifferentiation of the cells (passage 5) was induced by treatment with TGF-β1 (0.01 µg/ml, PeproTech, Rocky Hill). RNA of de-differentiated cells (day five of culturing) and redifferentiated cells (day 20 of culturing after induction of redifferentiation) was isolated and differentiation was controlled by determining collagen type II by RT-PCR. Dedifferentiated chondrocytes were treated with PD098059 (CALBIOCHEM, Nottingham, UK), MIA/CD-RAP and the antibody JBS5, binding and activating integrin alpha 5.
Human mesenchymal stem cells from CellSystems were cultivated in MSCGM medium (CellSystems) under a humidified atmosphere of 5% CO2 at 37℃. Cells were used in passages 4 to 10. Cells were treated with BMP2 (100 ng/ml) (PeproTech, Rocky Hill), MIA/CD-RAP (100 ng/ml), dorsomorphin (2 µM) (Sigma, Deisenhofen, Germany) and/or chordin (2 µg/ml) (R&D Systems, Minneapolis) for 48 h followed by RNA isolation.
RNA isolation and RT-PCR
For RT-PCR total cellular RNA was isolated from 3 cryo-sections of each embryo (100 µm) using the RNeasy kit (QIAGEN, Hilden, Germany). The integrity of the RNA preparations was controlled on a 1% agarose/formaldehyde gel. The reverse transcriptase reaction was performed in 20 µl reaction volume containing 2 µg of total cellular RNA, 4 µl of 5× first strand buffer (Invitrogen), 2 µl of 0.1 M DTT, 1 µl of dN6-primer (10 mM), 1 µl of dNTPs (10 mM, Amersham, Pittsburgh) and DEPCwater. The reaction mix was incubated for 10 min at 70℃. Then 1 µl of Superscript II reverse transcriptase (Invitrogen) was added and RNAs were transcribed for 1 h at 37℃. Subsequently, reverse transcriptase was inactivated at 70℃ for 10 min and RNA was degraded by digestion with 1 µl RNase A (10 mg/ml) at 37℃ for 30 min. cDNAs were controlled by PCR amplification of β-actin. PCR was performed using specific primers for mMIA, osteopontin, osteocalcin, aggrecan, Sox9, and ITGA5 (Tscheudschilsuren et al., 2006). mRNA expression was analyzed by quantitative real time-PCR on a Lightcycler (Roche, Mannheim, Germany) as described previously (Wenke et al., 2009).
Co-immunoprecipitation and western blot analysis
For analysis of BMP2 and MIA/CD-RAP interaction, BMP2 (2 µg, Peprotech) or BMP2 and biotinylated MIA/CD-RAP (2 µg (Bauer et al., 2006)) were added to 50 µl streptavidin-coupled G-sepharose (Pharmacia, Freiburg, Germany). The mixture was incubated for 1 h, pelleted, washed three times with binding buffer, resuspended in 20 µl Laemmli's buffer (Roth, Karlsruhe, Germany), heated at 95℃ for 5 min and separated on 12% SDS-polyacrylamide gels. BMP2 binding to MIA/CD-RAP was analyzed by western blotting using an anti BMP2 antibody (R&D Systems, Minneapolis).
For integrin alpha 5 co-immunoprecipitation 100 µg cell lysates dissolved in binding buffer (20 mM NaPO4, 150 mM NaCl, pH 7,5) were precleared with 25 µl protein streptavidin-coupled G-sepharose (Pharmacia) at 4℃ overnight. After centrifugation the supernatant was transferred into a fresh vial and incubated with biotinylated MIA with shaking at 4℃ overnight (Bauer et al., 2006). Then 50 µl streptavidin-coupled G-sepharose was added for 1 h, pelleted, washed three times with binding buffer, resuspended in 20 µl Laemmli's buffer, heated at 95℃ for 5 min and separated on 12% SDS-polyacrylamide gels. Integrin alpha 5 binding to MIA was confirmed by western blotting using an antibody against integrin alpha 5 (Chemicon International, Temecula).
Protein analysis in vitro (western blot analysis)
For protein isolation cells were washed with 1× PBS and lyzed in 200 µl RIPA-buffer (Roche, Mannheim, Germany). The protein concentration was determined using the BCA protein assay reagent (Pierce, Rockford). Balanced amounts of cell proteins (50 µg) were denatured at 70℃ for 10 min after addition of Roti-load-buffer (Roth, Karlsruhe, Germany) and subsequently separated on NuPage-SDS-gels (Invitrogen, Groningen, The Netherlands). After transferring the proteins onto PVDF-membranes (BioRad, Richmond), the membranes were blocked in 3% BSA/PBS for 1 h and incubated with a 1:1000 dilution of primary polyclonal rabbit anti-integrin alpha 5 or anti-P-ERK1/2 (R&D, antibody (Richmond; Chemicon, Hampshire, UK) over night at 4℃. A 1:2000 dilution of anti-rabbit-AP (Chemicon) was used as secondary antibody. Staining was performed using NBT/BCIP solution (Zytomed, Heidelberg, Germany). All experiments were repeated at least three times with similar results. For quantification blots were scanned and band intensity was evaluated in relation to the loading control.
Construction of the MIA targeting vector and generation of gene-disrupted mice
MIA/CD-RAP-deficient mice were generated as previously described (Moser et al., 2002).
For routine genotyping of the MIA/CD-RAP breeding pairs genomic tail-DNA was prepared and analyzed by PCR. The primer pair to amplify the wild type allele was: MIA-wt sense 5'-ATCCTATCTCCATGGCTGTGG-3'; MIA-wt anti 5'-GCCACTTTTGATGGTTTGCTGG-3'. The primer pair to amplify the targeted allele was: MIA-ko anti 5'-CCCATCAGCCTCACCGTAGGT-3', PGK-polyA down 5'-CGTCTCTTTACTGAAGGCTCTTT-3' (Sigma).
Histological analysis
The overall height of articular cartilage and calcified cartilage of the knee joint was measured on H&E sections by means of an ocular length scale (Olympus) (magnification ×40) in 12 week old wild type mice and MIA/CD-RAP-deficient mice. The cartilage height was measured in both the tibial and femoral articular surfaces of the joint in three high powered fields (HPF) next to the medial end of the lateral meniscus. Overall cartilage height was measured from the bone/cartilage interface to the surface of the cartilage. The height of the calcified cartilage was measured from bone/cartilage interface to tidemark. The number of groups of chondrocytes arranged in clusters or columns and of single chondrocytes was counted in 5 HPF in layers 2 and 3 of the tibial and femoral articular cartilage.
Statistical analysis
Results are expressed as mean ± S.E.M (range) or percent. Comparison between groups was made using the Student's paired t-test. A P value < 0.05 was considered significant. All calculations were performed using the GraphPad Prism software (GraphPad software Inc, San Diego).
Abbreviations
- BMP:
-
bone morphogenetic protein
- MIA:
-
melanoma inhibiting activity
- MIA/CD-RAP:
-
melanoma inhibiting activity/cartilage-derived retinoic acid-sensitive protein
References
Bauer R, Humphries M, Fassler R, Winklmeier A, Craig SE, Bosserhoff AK . Regulation of integrin activity by MIA . J Biol Chem 2006 ; 281 : 11669 - 11677
Blesch A, Bosserhoff AK, Apfel R, Behl C, Hessdoerfer B, Schmitt A, Jachimczak P, Lottspeich F, Buettner R, Bogdahn U . Cloning of a novel malignant melanoma-derivedgrowth-regulatory protein, MIA . Cancer Res 1994 ; 54 : 5695 - 5701
Bobick BE, Kulyk WM . The MEK-ERK signaling pathway is a negative regulator of cartilage-specific gene expression in embryonic limb mesenchyme . J Biol Chem 2004 ; 279 : 4588 - 4595
Bosserhoff AK, Buettner R . Establishing the protein MIA (melanoma inhibitory activity) as a marker for chondrocyte differentiation . Biomaterials 2003 ; 24 : 3229 - 3234
Bosserhoff AK, Kondo S, Moser M, Dietz UH, Copeland NG, Gilbert DJ, Jenkins NA, Buettner R, Sandell LJ . Mouse CD-RAP/MIA gene: structure, chromosomal localization, and expression in cartilage and chondrosarcoma . Dev Dyn 1997 ; 208 : 516 - 525
Bosserhoff AK, Stoll R, Sleeman JP, Bataille F, Buettner R, Holak TA . Active detachment involves inhibition of cell-matrix contacts of malignant melanoma cells by secretion of melanoma inhibitory activity . Lab Invest 2003 ; 83 : 1583 - 1594
Bullough PG, Jagannath A . The morphology of the calcification front in articular cartilage. Its significance in joint function . J Bone Joint Surg Br 1983 ; 65 : 72 - 78
Dietz UH, Sandell LJ . Cloning of a retinoic acid-sensitive mRNA expressed in cartilage and during chondrogenesis . J Biol Chem 1996 ; 271 : 3311 - 3316
Forsyth CB, Pulai J, Loeser RF . Fibronectin fragments and blocking antibodies to alpha2beta1 and alpha5beta1 integrins stimulate mitogen-activated protein kinase signaling and increase collagenase 3 (matrix metalloproteinase 13) production by human articular chondrocytes . Arthritis Rheum 2002 ; 46 : 2368 - 2376
Gannon JM, Walker G, Fischer M, Carpenter R, Thompson RC, Oegema TR . Localization of type X collagen in canine growth plate and adult canine articular cartilage . J Orthop Res 1991 ; 9 : 485 - 494
Hoyland JA, Thomas JT, Donn R, Marriott A, Ayad S, Boot-Handford RP, Grant ME, Freemont AJ . Distribution of type X collagen mRNA in normal and osteoarthritic human cartilage . Bone Miner 1991 ; 15 : 151 - 163
Hughes LC, Archer CW, ap Gwynn I . The ultrastructure of mouse articular cartilage: collagen orientation and implications for tissue functionality. A polarised light and scanning electron microscope study and review . Eur Cell Mater 2005 ; 9 : 68 - 84
Kirsch T, von der Mark K . Isolation of human type X collagen and immunolocalization in fetal human cartilage . Eur J Biochem 1991 ; 196 : 575 - 580
Lane LB, Villacin A, Bullough PG . The vascularity and remodelling of subchondrial bone and calcified cartilage in adult human femoral and humeral heads. An age- and stress-related phenomenon . J Bone Joint Surg Br 1977 ; 59 : 272 - 278
Lou J, Tu Y, Li S, Manske PR . Involvement of ERK in BMP-2 induced osteoblastic differentiation of mesenchymal progenitor cell line C3H10T1/2 . Biochem Biophys Res Commun 2000 ; 268 : 757 - 762
Lougheed JC, Holton JM, Alber T, Bazan JF, Handel TM . Structure of melanoma inhibitory activity protein, a member of a recently identified family of secreted proteins . Proc Natl Acad Sci USA 2001 ; 98 : 5515 - 5520
Moser M, Bosserhoff AK, Hunziker EB, Sandell L, Fassler R, Buettner R . Ultrastructural cartilage abnormalities in MIA/CD-RAP-deficient mice . Mol Cell Biol 2002 ; 22 : 1438 - 1445
Revell PA, Pirie C, Amir G, Rashad S, Walker F . Metabolic activity in the calcified zone of cartilage: observations on tetracycline labelled articular cartilage in human osteoarthritic hips . Rheumatol Int 1990 ; 10 : 143 - 147
Sriram D, Jones A, Alatli-Burt I, Darendeliler MA . Effects of mechanical stimuli on adaptive remodeling of condylar cartilage . J Dent Res 2009 ; 88 : 466 - 470
Stoll R, Bosserhoff A . Extracellular SH3 domain containing proteins--features of a new protein family . Curr Protein Pept Sci 2008 ; 9 : 221 - 226
Stoll R, Lodermeyer S, Bosserhoff AK . Detailed analysis of MIA protein by mutagenesis . Biol Chem 2006 ; 387 : 1601 - 1606
Stoll R, Renner C, Zweckstetter M, Bruggert M, Ambrosius D, Palme S, Engh RA, Golob M, Breibach I, Buettner R, Voelter W, Holak TA, Bosserhoff AK . The extracellular human melanoma inhibitory activity (MIA) protein adopts an SH3 domain-like fold . EMBO J 2001 ; 20 : 340 - 349
Tomiya M, Fujikawa K, Ichimura S, Kikuchi T, Yoshihara Y, Nemoto K . Skeletal unloading induces a full-thickness patellar cartilage defect with increase of urinary collagen II CTx degradation marker in growing rats . Bone 2009 ; 44 : 295 - 305
Tscheudschilsuren G, Bosserhoff AK, Schlegel J, Vollmer D, Anton A, Alt V, Schnettler R, Brandt J, Proetzel G . Regulation of mesenchymal stem cell and chondrocyte differentiation by MIA . Exp Cell Res 2006 ; 312 : 63 - 72
von der Mark K, Kirsch T, Nerlich A, Kuss A, Weseloh G, Gluckert K, Stoss H . Type X collagen synthesis in human osteoarthritic cartilage. Indication of chondrocyte hypertrophy . Arthritis Rheum 1992 ; 35 : 806 - 811
Walker GD, Fischer M, Gannon J, Thompson RC, Oegema TR . Expression of type-X collagen in osteoarthritis . J Orthop Res 1995 ; 13 : 4 - 12
Wenke AK, Grassel S, Moser M, Bosserhoff AK . The cartilage-specific transcription factor Sox9 regulates AP-2epsilon expression in chondrocytes . FEBS J 2009 ; 276 : 2494 - 2504
Xie WF, Zhang X, Sandell LJ . The 2.2-kb promoter of cartilage-derived retinoic acid-sensitive protein controls gene expression in cartilage and embryonic mammary buds of transgenic mice . Matrix Biol 2000 ; 19 : 501 - 509
Yee KL, Weaver VM, Hammer DA . Integrin-mediated signalling through the MAP-kinase pathway . IET Syst Biol 2008 ; 2 : 8 - 15
Yu PB, Hong CC, Sachidanandan C, Babitt JL, Deng DY, Hoyng SA, Lin HY, Bloch KD, Peterson RT . Dorsomorphin inhibits BMP signals required for embryogenesis and iron metabolism . Nat Chem Biol 2008 ; 4 : 33 - 41
Acknowledgements
We thank Sibylla Lodermeyer for excellent technical assistance. This work was supported by grants from the Deutsche Forschungsgemeinschaft to AKB.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Schubert, T., Schlegel, J., Schmid, R. et al. Modulation of cartilage differentiation by melanoma inhibiting activity/cartilage-derived retinoic acid-sensitive protein (MIA/CD-RAP). Exp Mol Med 42, 166–174 (2010). https://doi.org/10.3858/emm.2010.42.3.017
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3858/emm.2010.42.3.017
Keywords
This article is cited by
-
Bone secreted factors induce cellular quiescence in prostate cancer cells
Scientific Reports (2019)
-
Influence of cryopreservation, cultivation time and patient’s age on gene expression in Hyalograft® C cartilage transplants
International Orthopaedics (2013)
-
MIA is a potential biomarker for tumour load in neurofibromatosis type 1
BMC Medicine (2011)
-
Enhanced cartilage regeneration in MIA/CD-RAP deficient mice
Cell Death & Disease (2010)
